UNIVERSIDADE DE LISBOA Faculdade de Medicina Veterinária
QUALIDADE NUTRICIONAL DA CARNE DE CODORNIZ
ANDRÉ LUÍS DUARTE PARADA
CONSTITUIÇÃO DO JÚRI
Doutor José António Mestre Prates Doutor Rui José Branquinho de Bessa Doutor Mário Alexandre Gonçalves Quaresma
ORIENTADOR
Doutor Mário Alexandre Gonçalves Quaresma
CO-ORIENTADOR
Dr. Dário Alexandre Nunes de Sá Guerreiro
2019
LISBOA
UNIVERSIDADE DE LISBOA Faculdade de Medicina Veterinária
QUALIDADE NUTRICIONAL DA CARNE DE CODORNIZ
ANDRÉ LUÍS DUARTE PARADA
DISSERTAÇÃO DE MESTRADO INTEGRADO EM MEDICINA VETERINÁRIA
CONSTITUIÇÃO DO JÚRI
Doutor José António Mestre Prates Doutor Rui José Branquinho de Bessa Doutor Mário Alexandre Gonçalves Quaresma
ORIENTADOR
Doutor Mário Alexandre Gonçalves Quaresma
CO-ORIENTADOR
Dr. Dário Alexandre Nunes de Sá Guerreiro
2019
LISBOA
i Agradecimentos
Ao Professor Doutor Mário Quaresma, orientador deste trabalho, dirijo o meu sincero agradecimento pela orientação, prestabilidade, valiosas sugestões e críticas e ainda a colaboração oferecida na resolução de todos os problemas relacionados com a realização desta dissertação.
À Doutora Luísa Cristina Roseiro, o apoio incansável e incondicional na realização do trabalho prático deste estudo, empenho, incentivo e pelos preciosos ensinamentos transmitidos.
Ao Doutor Carlos Santos, as valiosas sugestões, colaboração dispensada e incentivo sempre demonstrados em qualquer circunstância.
À Dra Helena Viegas pela valiosa ajuda no trabalho prático e à sua constante boa disposição diária.
À Engenheira Irene Cristina Antunes pelo incentivo amigo, preciosa paciência e ajuda na resolução de problemas informáticos.
Ao Dr. Dário Alexandre Nunes de Sá Guerreiro, co-orientador deste trabalho, apresento um agradecimento muito especial, pelo apoio científico que sempre me ofereceu, pelo estímulo diário, pela sua amizade demonstrada bem como pela confiança que me foi depositada. Ao Dr. Marco Barros, director geral da empresa Interaves, um agradecimento muito especial por ter cedido gentilmente as codornizes que tornaram possível a realização desta dissertação.
Ao Sr. Jacinto Amaro, presidente da Fencaça pela cedência de metade das codornizes selvagens usadas no estudo (provenientes do Alentejo e Ribatejo, ao Eng. Agostinho Beça e ao Eng. Rui Tadeu pela cedência das restantes codornizes (provenientes dos Concelhos de Mirandela e Bragança).
Ao Dr. José Cordeiro e ao Dr. João Barbosa por me proporcionarem o estágio na Auchan e pela amizade demonstrada.
Aos meus amigos, em especial à Sandra Liz, Inês Sanches, Vitória Daniel, Joana Domingues, Diana Lopes, Rita Reis, Fábio Santos e João Vinagre pela sua amizade, paciência e boa disposição sempre demonstrada principalmente nos momentos de desespero.
Ao meu pai pelo apoio incondicional, amizade e disponibilidade que sempre me concedeu ao longo da minha vida académica.
À Ana, a minha companheira, que sempre me apoiou e incentivou em todos os momentos de crise e desespero desta minha jornada com sorriso, coragem, paciência e muito mais. À minha mãe.
ii Resumo
Dentro das aves de capoeira, a codorniz é simultaneamente usada para a produção de carne e ovo. A codorniz europeia (Coturnix coturnix) é simultaneamente uma espécie cinegética, sendo a caça à codorniz uma actividade venatória que agrega o interesse de muitos caçadores na Península Ibérica.
A Codorniz japonesa (Coturnix japonica) é a espécie usada para produção de carne e ovo, sendo a Codorniz europeia criada essencialmente para a produção de carne.
A ausência de estudo que compare a codorniz europeia silvática com a proveniente da produção, nem um estudo que faça a comparação da codorniz europeia com a japonesa, levou-nos a pensar realizar este estudo. Assim sendo, foi objectivo deste estudo caracterizar o perfil de ácidos gordos e de aminoácidos e quantificar o teor de colesterol e α-tocoferol da carne de codorniz, Coturnix coturnix (produção e silvática) e Coturnix japonica.
O estudo utilizou 40 codornizes de aviário (20 exemplares de codorniz europeia e 20 exemplares de codorniz japonesa) e 20 codornizes europeias silváticas, abatidas durante a época reservada à caça a esta espécie. A caracterização dos ácidos gordos foi realizada por Cromatografia gasosa. A caracterização do perfil de aminoácidos e a determinação do teor de colesterol e de vitamina E foi efectuado por Cromatografia Líquida de Alta Pressão. Os resultados obtidos permitiram verificar que a carne de codorniz é, independentemente da espécie e origem, uma excelente fonte de aminoácidos (21,19-23,50 g/100 g de carne), sendo esta rica em aminoácidos essenciais (7,94-9,10 g/100 g de carne). O perfil de ácidos gordos é dominado pelos PUFA (39,9-47,1 g/100 g de ácidos gordos). O teor de colesterol e de α-tocoferol oscilaram entre 64,4-72,7 mg/100 g de carne e 12,4-14,94 µg/g de carne, respectivamente.
Palavras Chave: Coturnix coturnix, Coturnix japonica, aminoácidos, ácidos gordos, colesterol, α-tocoferol.
iii Abstract
Hunting birds, especially quail, have always attracted a great deal of attention from both gastronomers and hunters. Global meat consumption shows a growing demand for poultry meat and there is also a demand for different products with high nutritional value and gourmet qualities. We think that quail meets these four characteristics, but there is no study in the literature comparing European quail production with sylvatic, nor a study comparing the european quail with japonica.
Thus, the objective of this study was to characterize the fatty acid and amino acid profiles and to quantify the cholesterol and α-tocopherol content of the quail meat, Coturnix coturnix (production and sylvatic) and Coturnix japonica.
The study used 40 poultry quail (20 specimens of European quail and 20 specimens of Japanese quail) and 20 European silvatic quail, slaughtered during the time reserved for hunting this species. The characterization of fatty acids was performed by gas chromatography. The characterization of the amino acid profile and the determination of the cholesterol and vitamin E content was performed by high pressure liquid chromatography. The results obtained showed that quail meat is an excellent source of protein (21,19-23,50 g/100 g of meat), which is rich in essential amino acids (7,94-9.10 g/100 g of meat). The fatty acid profile is dominated by PUFAs (39.9-47.1 g/100 g fatty acids). The cholesterol and α-tocopherol content ranged from 64.4-72.7 mg/100 g of meat and 12.4-14.94 μg/g of meat, respectively.
Keywords: Coturnix coturnix, Coturnix japonica, amino acid, fatty acids, cholesterol, α-tocopherol.
iv Índice Agradecimentos ... i Resumo ... ii Abstract ... iii Índice ... iv
Índice de Quadros ... viii
Índice de Figuras ... ix Abreviaturas ... x
1.
Relatório de Estágio ... 1
2.
Introdução ... 2
3.
Revisão bibliográfica ... 3
3.1. Género ... 3 3.2. Codorniz europeia/comum ... 5 3.2.1. Taxonomia ... 5 3.2.2. Características morfológicas ... 5 3.2.3. Distribuição/Migração ... 63.2.4. Estimativa populacionais de Coturnix coturnix ... 8
3.2.5. Reprodução ... 10
3.3. Codorniz japonesa ... 11
3.3.1. Taxonomia ... 11
3.3.2. Características ... 11
3.3.3. Domesticação ... 12
3.4. Características comuns de C. coturnix e C. japonica ... 12
3.4.1. Dimorfismo sexual ... 12
3.4.2. Distinção jovem/adulto ... 13
3.4.3. Hibridismo ... 14
3.4.4. Vocalização ... 15
3.5. Potencial económico e de investigação ... 15
v
3.5.1.1. Produção de carne ... 16
3.5.1.2. Produção de ovos ... 17
3.5.1.3. Comparação da codorniz com outras aves de capoeira usadas tradicionalmente para a produção de carne e ovo ... 18
3.5.1.4. Produção da codorniz como animal de caça e desporto ... 19
3.5.1.5. Produção da codorniz como animal ornamental ... 20
3.5.1.6. Comercialização ... 20
3.5.2. Potencial de investigação ... 20
3.6. Enquadramento Geral da Produção ... 21
3.7. Composição da carne ... 22
3.7.1. Ácidos gordos ... 23
3.7.2. Aminoácidos ... 24
3.7.3. Colesterol ... 26
3.7.4. Vitamina E ... 27
4.
Caracterização da qualidade nutricional da carne de codorniz ... 29
4.1. Objectivo ... 29
4.2. Material e métodos ... 29
4.2.1. Material biológico ... 29
4.2.2. Preparação da amostra ... 30
4.2.3. Determinação do perfil de aminoácidos por HPLC ... 30
4.2.3.1. Material utilizado ... 30
4.2.3.2. Reagentes utilizados ... 30
4.2.3.3. Soluções preparadas ... 30
4.2.3.4. Hidrólise ... 31
4.2.3.5. Derivatização ... 31
4.2.3.5.1. Derivatização de aminoácidos primários ... 31
4.2.3.5.2. Derivatização de aminoácidos secundários ... 31
4.2.3.6. HPLC ... 31
4.2.3.6.1. Equipamento ... 31
vi
4.2.3.6.3. Separação cromatográfica dos aminoácidos secundários ... 32
4.2.3.7. Preparação dos padrões ... 32
4.2.4. Determinação do perfil de ácidos gordos ... 32
4.2.4.1. Material utilizado ... 32
4.2.4.2. Reagentes utilizados ... 32
4.2.4.3. Soluções preparadas ... 32
4.2.4.4. Transesterificação e extracção dos ácidos gordos ... 33
4.2.4.5. Quantificação e identificação dos ácidos gordos ... 33
4.2.4.6. Cálculo dos índices e rácios de qualidade do perfil de ácidos gordos ... 33
4.2.4.6.1. Índice de Aterogenicidade ... 33 4.2.4.6.2. Índice de Trombogenicidade ... 34 4.2.4.6.3. Índice de Peroxibilidade... 34 4.2.4.6.4. Rácio P/S ... 34 4.2.4.6.5. Rácio n-6/n-3 ... 34 4.2.4.6.6. Rácio h/H ... 34
4.2.5. Determinação do colesterol e vitamina E ... 34
4.2.5.1. Material utilizado ... 34 4.2.5.2. Reagentes ... 35 4.2.5.3. Soluções preparadas ... 35 4.2.5.4. Extracção de saponificação ... 35 4.2.5.5. HPLC ... 36 4.2.5.5.1. Equipamento ... 36 4.2.5.5.2. Reagentes ... 36 4.2.5.6. Análise ... 36 4.2.6. Análise Estatística ... 36 4.3. Resultados e Discussão ... 37
4.3.1. Caracterização do perfil de ácidos gordos ... 37
4.3.2. Caracterização do perfil de aminoácidos ... 40
4.3.3. Teor de colesterol e vitamina E... 42
vii
viii Índice de Quadros
Quadro 1 - Espécies de codornizes, sub-espécies e sua localização ... 4
Quadro 2 - População europeia reprodutiva de Coturnix coturnix. ... 9
Quadro 3 - Distinção entre macho e fêmea ... 12
Quadro 4 - Distinção entre animais jovens e adultos... 13
Quadro 5 - Aves abatidas e aprovadas para consumo público ... 22
Quadro 6 - Efectivo de codornizes em cabeças e toneladas em Portugal ... 22
Quadro 7 - Aminoácidos essenciais e não essenciais ... 25
Quadro 8 - Amostras de carne utilizados no presente estudo, sua designação, espécie, forma e origem geográfica. ... 29
Quadro 9 – Perfil de ácidos gordos em carne de peito de codorniz de produção e silvática. ... 38
Quadro 10 - Somatórios parciais dos ácidos gordos, rácios e índices de qualidade nutricional em peito de codorniz silváticas e de produção. ... 39
Quadro 11 – Perfil de aminoácidos no peito de codorniz de produção e de caça (expresso em g/100 g de carne), somatório parcial de aminoácidos essenciais (AAE), não essenciais (AANE) e teor total de aminoácidos (TAA). ... 41
Quadro 12 – Teores de colesterol e de vitamina E em carne de peito de codorniz de produção e silváticas. ... 43
ix Índice de Figuras
Fig. 1 - Diversas espécies do género Coturnix. Fig. 1a. Coturnix coromandelica. Fig. 1b.
Coturnix pectoralis. Fig. 1c. Coturnix adansonii. Fig. 1d. Coturnix novaezealandiae. Fig. 1e. Coturnix japonica. Fig. 1f. Coturnix delegorguei. Fig. 1g. Coturnix ypsilophora. Fig. 1h.
Coturnix chinensis. Fig 1i. Coturnix coturnix ... 3
Fig. 2- Codorniz comum macho (a) e codorniz comum fêmea (b). ... 5
Fig. 3 - Distribuição da população Coturnix coturnix. ... 6
Fig. 4 – Principais rotas migratórias de Coturnix coturnix ... 7
Fig. 5- Áreas de reprodução da codorniz europeia (linhas verticais) e da codorniz japonesa (linhas horizontais), mostrando a área de sobreposição ... 14
Fig. 6- Produtores mundiais de carne de codorniz ... 21
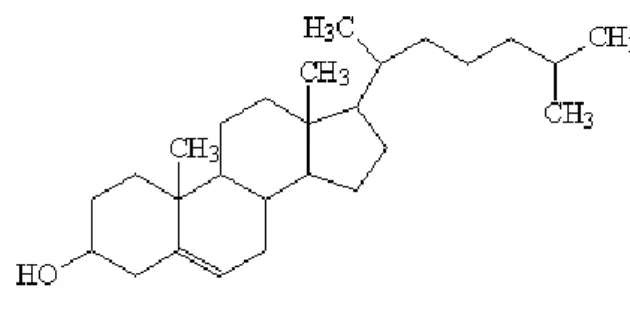
Fig. 7 - Estrutura molecular do colesterol ... 26
x Abreviaturas % - Percentagem µg - Micrograma μl - Microlitro μm - Micrómetro AA – Aminoácido
AAE – Aminoácido essencial AANE – Aminoácido não essencial ARA - Ácido araquidónico
AG - Ácido gordo AL - Ácido linoleico ALA – Ácido α-linolénico
CCP – Coturnix coturnix Produção CCS – Coturnix coturnix Selvagem CJP – Coturnix japonica Produção cm – Centímetro
DFD – Dark, firm, dry
DHA - Ácido docosahexaenóico DPA – Ácido docosapentaenóico EPA - Ácido eicosapentaenóico
FMOC-Cl – Cloreto 9-Fluorenilmetoxicarbonilo g - Grama
h/H - Rácio de hipo/hipercolesterolémico
HPLC - Cromatografia líquida de alta performance IA - Índice de Aterogenicidade
INIAV –Instituto Nacional de Investigação Agrária e Veterinária Kcal – Kilocaloria
Kj - Kilojoule KPa - Kilopascal L - Litro
LDL - Lipoproteínas de baixa densidade MUFA - Ácidos gordos monoinsaturados m - Metro
mg - Miligrama min - Minuto mL - Mililitro mm - Milímetro
xi n6/n3 - Rácio ómega-6/ómega-3 nm - Nanómetro ºC - Graus Celsius OPA – O-ftaldeído P - p-value
P/S - Rácio ácidos gordos polinsaturados/ ácidos gordos saturados pH - Potencial de hidrogénio
PI - Índice de Peroxibilidade PSE – Pale, soft, exsudative
PUFA – Ácidos gordos polinsaturados Rpm- Rotações por minuto
RSD – Residual Standard Deviation SEM – Standard Error of Mean SFA - Ácidos gordos saturados TAA – Total de aminoácidos TI - Índice de Trombogenicidade WHO – World Health Organization
WHO/FAO - World Health Organization/Food and Agriculture Organization UTI – Unidade de Tecnologia e Inovação
α-TTP - Proteína de transferência do α-Tocoferol β - Beta
γ - Gama δ - Delta Σ - Somatório
1 1. Relatório de Estágio
O sexto ano do Mestrado Integrado em Medicina Veterinária (MIMV) foi dividido em três períodos de estágio. O primeiro, de âmbito extracurricular, foi realizado na direcção de qualidade do grupo Auchan, sob a orientação do Dr. José Cordeiro e do Dr. João Barbosa, entre Outubro e Dezembro de 2017 tendo realizado trabalhos emacções de auditoria a fornecedores, acompanhamento de operações de recepção, execução e expedição do sector de controlo e da qualidade e segurança dos alimentos. O segundo estágio, curricular, foi realizado na Sá Guerreiro Vet, Lda, sob a égide do Dr. Dário Sá Guerreiro, com a duração de seis meses, entre Março e Agosto de 2018, completando um total de 2200 horas. Os principais objectivos deste estágio curricular incidiram sobretudo na área clínica, cirúrgica, reprodutiva e saneamento de bovinos e pequenos ruminantes. Assim este estágio permitiu o desenvolvimento e participação em diversas áreas e actividades, tais como:
a) Acompanhamento na realização de saneamentos a bovinos, onde foram efectuados testes intradérmicos de tuberculina, recolha de sangue para despiste da Brucelose; b) Acompanhamento na leitura de reacções dos testes intradérmicos da tuberculina; c) Acompanhamento na realização de tarefas cirúrgicas, nomeadamente cesarianas,
deslocações do abomaso à esquerda e à direita, prolapso uterinos, vaginais e rectais e amputações de cauda em bovinos;
d) Acompanhamento de casos clínicos em bovinos, pequenos ruminantes e animais de companhia;
e) Realização de diagnósticos de gestação por palpação rectal em bovinos leiteiros e de carne.
O último período de estágio decorreu no âmbito da dissertação de Mestrado Integrado de Medicina Veterinária, tendo sido realizado em duas instituições diferentes entre Outubro e Dezembro de 2018, totalizando 550 horas. Primeiramente foi realizado a transesterificação e extracção dos ácidos gordos e extracção e saponificação do colesterol e vitamina E, no laboratório do departamento de Bioquímica da Faculdade de Medicina Veterinária da Universidade de Lisboa, sob a orientação do Professor Doutor Mário Quaresma (docente da Faculdade de Medicina Veterinária da Universidade de Lisboa). Na UTI do INIAV, sob a supervisão da Doutora Cristina Roseira e da Doutora Ana Partidário foi realizado, a separação cromatográfica dos aminoácidos, a análise do colesterol e vitamina E, identificação e quantificação dos ácidos gordos.
2 2. Introdução
A codorniz é um animal muito conhecido e apetecido entre caçadores e gastrónomos. Pensa-se que a primeira domesticação tenha ocorrido no século XII no Japão. Embora existam representações de codornizes nos Hieróglifos Egípcios do ano 2000 A.C (Wakasugi, 1984).
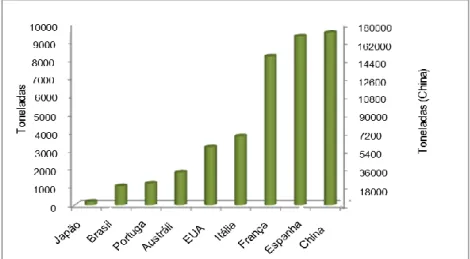
Nos dias de hoje a coturnicultura, designação dada à criação de codornizes, espalhou-se rapidamente pela Europa devido a algumas das suas características, como ser dotada de uma alta fertilidade, robustez, resistência a doenças, postura abundante e necessidade de pouco espaço para o seu confinamento. Nas últimas décadas, o interesse comercial tem aumentado sendo a China o principal produtor, 168,000 toneladas, seguido de Espanha e de França, respectivamente com 9,300 toneladas e 8,200 toneladas (Poultry World, 2018). A codorniz é utilizada na produção de carne e ovo devido, ao seu alto potencial reprodutivo, baixo consumo de ração e baixo custo de criação (Minvielle, 2004). Nos países em desenvolvimento, a criação de codornizes é uma boa opção porque possibilita a produção de proteína de boa qualidade, produzida de forma econónima e num espaço reduzido (Sundaram, 1988; Narinc, Aksoy & Karaman, 2010) e dos seus ovos cada vez mais requisitados pelo mercado de produtos “gourmet” devido ao seu sabor exótico (Fabichak, 1989).
Os comportamentos alimentares das pessoas, principalmente nas grandes cidades, sofreram grandes alterações associado ao modo de vida, alterações dos hábitos e tendências sociais. A procura dos alimentos nutricionalmente equilibrados e benéficos para a saúde, aumentou graças à consciencialização dos consumidores. Os produtos cárneos e a carne tiveram o mesmo tipo de selecção. Devido à associação do consumo de carne vermelha com doenças cardiovasculares e oncológicas, verificou-se um aumento do consumo de carne de aves de capoeira, nomeadamente de frango, peru e codorniz (Sadler, 2004; Karakok, Ozogul, Sager, Mucella & Ozogul, 2010; Amaral, Mafra & Oliveira, 2015). Este trabalho surgiu, face à crescente procura da carne de aves cinegéticas e à grande lacuna de informação existente na composição da sua carne, nomeadamente a codorniz. O objectivo deste trabalho incidiu na avaliação da composição da carne de codorniz, de produção e de caça, com foco para a determinação do teor de colesterol, vitamina E, perfil de ácidos gordos e perfil de aminoácidos.
3 3. Revisão bibliográfica
3.1. Género
O género Coturnix é composto por nove espécies (Figura 1) e encontra-se dividido, conforme a sua origem geográfica, África, Ásia, Austrália e ilhas do Atlântico (Lucotte, 1982). A codorniz é frequentemente classificada como galináceo. Este termo não possui qualquer valor taxonómico, mas é frequentemente utilizado pela maioria dos ornitologistas para englobar todas as aves das Ordens Galliformes e Craciformes. Aves como a codorniz, a perdiz, o faisão, o pavão, o perú e muitas outras aves estão incluídas nestas ordens. Recentemente, alguns autores têm sugerido apenas a ordem dos Galliformes, estando esta dividida nas subordens Craci, com as famílias Megapodiidae e Cracidae, e a subordem Phasiani que inclui as famílias Numididade, Meleagridae, Tetraonidade, Odontophoridae e Phasianidae (Dias, 1995).
A ordem dos Galliformes conta com mais de 70 géneros, sendo este um dos mais ricos em espécies. No género Coturnix dentre as diversas espécies, as mais conhecidas e as estudadas neste trabalho são a codorniz comum, Coturnix coturnix, e a codorniz japonesa Coturnix japonica. Ambas pertencem à família Phasianidae da ordem Galliformes, grupo taxonómico onde se encontram aves domésticas como galinhas, pérus e ainda diversas espécies cinegéticas como faisões e perdizes (del Hoyo, Elliott & Sargatal, 1994). No Quadro 1, são indicadas as espécies, os nomes científicos, a localização e as suas sub-espécies.
Fig. 1 - Diversas espécies do género Coturnix. Fig. 1a. Coturnix coromandelica. Fig. 1b.
Coturnix pectoralis. Fig. 1c. Coturnix adansonii. Fig. 1d. Coturnix novaezealandiae. Fig.
1e. Coturnix japonica. Fig. 1f. Coturnix delegorguei. Fig. 1g. Coturnix ypsilophora. Fig. 1h. Coturnix chinensis. Fig 1i. Coturnix coturnix (adaptado de orientalbirdimages.org,
2018; biolib.cz, 2018).
a b c d e
4
Quadro 1 - Espécies de codornizes, sub-espécies e sua localização (adaptado del Hoyo et al., 1994;
Integrated Taxonomic Information System, 2018).
Espécie Nome Científico Sub-espécies Localização
Codorniz coromandelica
Coturnix coromandelica
Gmelin, 1979 Monotípica Sul da Ásia
Codorniz pectoral Coturnix pectoralis
Gould, 1837 Monotípca Austrália
Codorniz azul
Coturnix adansonii
J. Verreaux & E. Verreaux, 1851
Monotípica África
Codorniz da Nova Zelândia
Coturnix novaezealandiae
Quoy & Gaimand, 1830 Monotípica Nova Zelândia
Codorniz japonesa Coturnix japonica
Temminck & Schlegel, 1849 Monotípica Oceania, Europa, Ásia
Codorniz arlequim Coturnix delegorguei
Delegorgue, 1847 3 Sub-espécies
África do Sul, Costa do Marfim, Etiópia e
Madagáscar
Codorniz marrow Coturnix ypsilophora
Bosc, 1792 9 Sub-espécies
Austrália Sudoeste Ásia
Codorniz chinesa
Coturnix chinensis
Linnaeus, 1766 9 Sub-espécies Oceânia, Austrália, Índia,
Codorniz comum Coturnix coturnix
Linnaeus, 1758 Europa, Ásia e África
Coturnix coturnix Africana
Temminck & Schlegel, 1849
Coturnix coturnix conturbans
Hartert, 1917
Coturnix coturnix coturnix
Linnaeus, 1758
Coturnix coturnix erlangeri
Zedlitz, 1912
Coturnix coturnix inopinata
__________________________
5
Revisão bibliográfica
3.2. Codorniz europeia/comum 3.2.1. Taxonomia
A codorniz europeia ou a codorniz comum, Coturnix coturnix Linnaeus, 1758, encontra-se espalhada por toda a Europa, Ásia, África e nas ilhas do Atlântico (Lucotte, 1982), estando classificada da seguinte forma (www.itis.gov, 2018):
Reino – Animalia Filo – Chordata Classe – Aves Ordem – Galliformes Família – Phasianidae Género – Coturnix
Espécie – Coturnix coturnix
3.2.2. Características morfológicas
A estatura média da codorniz comum apresenta um comprimento entre 19 a 25 cm, uma envergadura de 32 a 35 cm e um peso médio entre 85 a 120 gramas (Gallego, Rodríguez-Teijeiro & Puigcerver, 1990). Estas aves não apresentam dimorfismo sexual a nível do peso, com excepção das fêmeas em cativeiro que são mais pesadas que os machos. A cor da plumagem em ambos os sexos é bastante semelhante, apresentando padrões de castanho, malhado, listado no dorso e a parte inferior castanho alaranjado (Fig. 2) (Wetherbee, 1961).
A ave quando se encontra em repouso apresenta um corpo arredondado e cauda muito curta, o que lhe permite uma camuflagem no meio da vegetação. As asas são compridas e estreitas em relação ao corpo, adaptadas a voos de longa distância, nomeadamente os intercontinentais. O seu voo caracteriza-se por ser rápido, recto e baixo com batidas rápidas das asas (Sousa, 2014).
Fig. 2- Codorniz comum macho (a) e codorniz
comum fêmea (b) (adaptado de
orientalbirdimages.org, 2018).
6 3.2.3. Distribuição/Migração
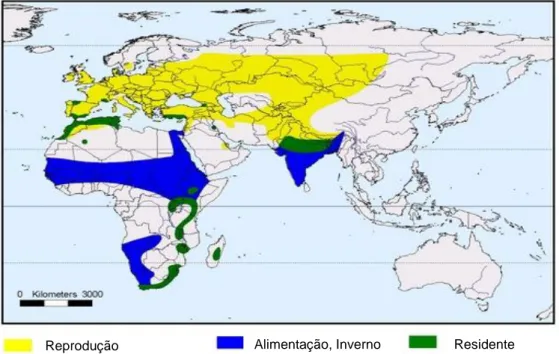
A codorniz comum é a única espécie migrante de longa distância de toda a família Phasianidae (Snow & Perrins, 1998), distribuída pela área geográfica Paleártico ocidental e Ásia ocidental e central (Rodríguez-Teijeiro et al., 2010) (Fig. 3).
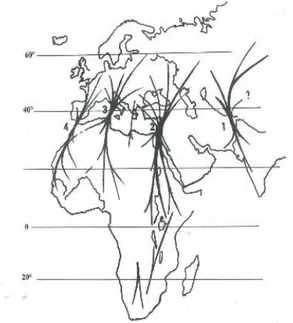
Em 2003, Guyomarc`h descreveu quatro rotas migratórias para a codorniz, o fluxo do grande vale do Indus, o fluxo da Arábia, o fluxo Apeniniano e o fluxo através da costa Atlântica (Marrocos para a Península Ibérica e desta para França, Alemanha, Escandinávia e Inglaterra) (Fig. 4).
O processo de migração da C. coturnix é bastante complexo (Fig. 4), sem similaridade com as restantes aves. Além de longos, médios e curtos migradores, há também indivíduos sedentários (Fontoura & Gonçalves, 1993). Em 2010, Rodríguez-Teijeiro e seus colaboradores descreveram as movimentações desta ave, como migratórios de chegada (para a reprodução na primavera) e de partida (no fim do verão), movimentos nómadas ou erráticos (simples deslocações de curta distância, à procura de habitats mais favoráveis mesmo que temporariamente) e movimentos dos machos à procura de fêmeas para acasalar.
A distribuição das populações é determinada principalmente pelo habitat, que depende de factores agrícolas e climáticos (Puigcerver, Sardá-Palomera & Rodríguez-Teijeiro, 2012). A acção da chuva provoca o atraso da colheita dos campos de cereais, o que favorece um habitat adequado à espécie (Sardà-Palomera & Vieites, 2011), porque facilita o crescimento de plantas herbáceas nas margens dos campos e de restolho, incrementando uma superfície com uma cobertura vegetal adequada para a codorniz (Rodríguez-Teijeiro,
Reprodução Alimentação, Inverno Residente
__________________________
7
Revisão bibliográfica
Puigcerver, Rodrigo-Rueda & Gallego, 1996). A codorniz comum reproduz-se, em todas as latitudes da Península Ibérica e com altitudes inferiores a 2100 metros (Guyomarc´h, 2003). No entanto, os habitats mais adequados para as espécies são as culturas de cereais secos, que representam 19% da área mediterrânica da península (Sardà-Palomera & Vieites, 2011).
Segundo Fontoura & Gonçalves em 1993, o estatuto migratório das aves pode ser avaliado, pela largura da banda de gordura subcutânea na região peitoral (quando superior a 10 mm, a ave está em migração) e pela cor da gordura (branca o indivíduo encontra-se no início da migração, quando amarela e oleosa indica o fim da migração).
A migração das fêmeas é feita conjuntamente com a sua descendência, separadas dos machos, ocorrendo normalmente durante a noite (Uljanin, 1941).
Fig. 4 – Principais rotas migratórias de
Coturnix coturnix (adaptado de Perennou, 2009).
8
3.2.4. Estimativa populacionais de Coturnix coturnix
São vários os registos, que demonstram a dificuldade em fornecer estimativas confiáveis de populações de C. coturnix (Puigcerver et al., 2012). Tal facto resulta de várias dificuldades, nomeadamente: 1) a prática de alguns países da Europa, repovoarem as populações de C. coturnix com codornizes da espécie C. japonica ou híbridas, embora estas tenham sido proibidas em Portugal, Espanha e França (Barilani et al., 2005; Amaral et al., 2007; Sanchez-Donoso et al., 2012); 2) o momento da colheita dos cereais (trigo e cevada) combinado com o relevo geográfico poderá actuar como funil, forçando as populações de codorniz a concentrarem-se com movimentos pré-migratórios em determinadas áreas (Rodríguez-Teijeiro et al., 2010); 3) o ciclo reprodutivo desta ave, que ocorre dentro das culturas de cereais, passando despercebido. Os machos podem ser detectados acusticamente, enquanto que as fêmeas permanecem acústica e visualmente imperceptíveis (Guyomarc`h, 2003). O tamanho da população destas aves tem sofrido grande flutuação no passado, tendo registado uma diminuição acentuada nos anos 70 (Derégnacourt, Guyomac´h & Belhormra, 2005). Foram reportados muitos factores que causaram este declínio, nomeadamente a destruição de habitats pela intensificação agrícola e pela caça excessiva, que esta espécie é submetida na bacia do mediterrâneo (Hagemeijer & Blair, 1997).
Registos mais recentes sugerem que nas últimas duas décadas, as populações portuguesa, espanhola e francesa de C. coturnix mantiveram-se estáveis, contudo as duas últimas populações mostraram um certo declínio (Rodríguez-Teijeiro et al., 2010).
Tendo em consideração, a dificuldade metodológica em fornecer dados confiáveis da população de Coturnix coturnix, a população europeia foi estimada em 1,4 a 2,75 milhões de casais de codorniz (Quadro 2) (Guyomarc`h, 2003). Embora a Bird lifeInternational tenha, em 2004, estimado os casais de codorniz em 2,8 a 4,7 milhões (Quadro 2).
Da população mundial de codornizes apenas 5 a 24% se reproduz na Europa. Desta população europeia 35 a 37% reproduz-se na Rússia, com a presença de grandes populações na Bielorrússia, Polónia, Turquia, Roménia e Ucrânia (Quadro 2) (Guyomarc`h, 2003). Grande proporção da população europeia de codornizes reproduz-se em Espanha, França e Polónia. A população de codornizes da União Europeia totaliza valores entre 884,000 e 1.912,000 de machos. Estes registos quando comparados com a população total, possivelmente aferem a sua favorável conservação (Perennou, 2009).
__________________________
9
Revisão bibliográfica
Quadro 2 - População europeia reprodutiva de Coturnix coturnix.
Bird Life International, 2004 Guyomarc`h, 2003
País Casais reprodutores Ano Tendência 1990-2000 Machos territoriais Alemanha 12000-32000 1995-1999 (+) 20000-100000 Áustria 5000-15000 1998-2002 (+) (5000-15000) Bielorrússia 15000-30000 100000-150000 Bélgica 2400-5700 2001-2002 (0) 3000-7000 Bulgária 8000-15000 50000-200000 Chipre (1000-4000) 1994-2002 (0) / Dinamarca 200-600 2000-2001 + / Eslováquia 2000-6000 1990-1999 0 / Eslovénia 1000-2000 1999 0 / Espanha (320000-435000) 1992 ? 500000-1000000 Estónia 10-50 1998 0 / Finlândia 10-100 1998-2002 + / França (100000-500000) 2000 (F) 150000-300000 Grécia (2000-5000) 1995-2000 (-) / Holanda 2000-6500 1998-2000 + 1000-10000 Hungria 70000-94000 1999-2002 (0) 30000-45000 Irlanda 0-20 1988-1991 F / Itália (5000-20000) 2003 ? 100000-150000 Letónia 20-500 1990-2000 + / Lituânia (1000-2000) 1999-2001 + / Luxemburgo 10-25 2002 0 / Malta 1-3 1990-2002 - / Polónia 100000-150000 2000-2002 (+) 20000-130000 Portugal (5000-50000) 2002 (0) 50000-150000 Reino Unido 5-450 1996-2000 0 / República Checa 5000-10000 2000 + 10000-20000 Roménia 160000-220000 1999-2002 (-) 30000-50000 Rússia 1500000-2000000 2000 F 1000000-2000000 Suécia (10-40) 1999-2000 (F) / Turquia (300000-800000) 2001 (-) 250000-500000 Ucrânia 100000-160000 1990-2000 - 500000-600000
+ Aumentar; 0 Estável; - Diminuir; F Flutuante; ? Desconhecido; () Pouco conhecido, sem dados quantitativos disponíveis; / Sem dados
10 3.2.5. Reprodução
O macho da codorniz comum adquire a sua maturidade sexual aos 28 dias de idade, enquanto a ovopostura de ovos férteis ocorre pelas fêmeas aos 31 dias de idade (Wetherbee, 1961).
As fêmeas podem ser beneficiadas por um ou vários machos (Puigcerver, Gallego, Pérez, Barroso & Rodríguez-Teijeiro, 2000). Muitos autores consideram a espécie Coturnix coturnix poligâmica (Dementev & Gladkov, 1952; Rodrigo-Rueda et al., 1997; Rodríguez-Teijeiro et al., 2003), enquanto que outros consideram-na monogâmica sazonal, excepto quando existe um excedente de fêmeas num determinado local, tornando deste modo possível a poligamia (Moreau, 1951). Mais recentemente em 2004, Puigcerver considerou as fêmeas poliândricas e os machos poligâmicos.
Os machos permanecem junto das fêmeas apenas durante o tempo estritamente necessário (aproximadamente 18 dias) para garantir que esta ficou fecundada. Este acontecimento designa-se por competência do esperma (Gallego, Rodríguez-Teijeiro & Puigcerver, 1990) ou por custódia da fêmea (Rodrigo-Rueda, Puigcerver, Rodríguez-Teijeiro, Sanz & Gallego, 1993).
Durante o período de reprodução, num espaço de tempo de 15 dias, existe um intercâmbio de machos (95%) entre diferentes nichos reprodutivos (Rodríguez-Teijeiro et al., 2010). As fêmeas durante a época reprodutiva encontram-se, em diferentes fases do seu ciclo reprodutivo, caracterizando-se por assincronia reprodutiva (Gallego et al., 1990).
A codorniz nidifica no chão, devido aos seus hábitos terrestres, num local onde ficará praticamente invisível. Na Europa, em geral a fase de nidificação vai desde meados de maio até finais de junho (Lack, 1950; Fontoura & Gonçalves, 1993). O nascimento das jovens codornizes, normalmente, encontra-se relacionado com o crescimento de culturas num determinado país ou região (Lack, 1950). Como já foi referido anteriormente, e citado por Guyomarc`h em 2003, a Península Ibérica reúne características propícias à reprodução desta espécie. A C. coturnix chega a Portugal e Espanha, precedente de África, para se reproduzir (Rodríguez-Teijeiro et al., 1996).
Para avaliação da maturidade e actividade sexual, Fontoura & Gonçalves em 1993, propõem um método baseado nas dimensões e no aspecto da fenda cloacal. As fêmeas com 3 mm de fenda cloacal encontram-se em repouso, com 5 mm no início do processo reprodutivo e com 10 mm quando em postura. Nos machos a fenda cloacal não excede os 7 mm, intumescida e com bolsa vermelha contendo secreção branca, visível quando pressionada.
__________________________
11
Revisão bibliográfica
3.3. Codorniz japonesa 3.3.1. Taxonomia
A codorniz japonesa, Coturnix japonica Temminck & Schlegel, 1849, foi domesticada há muito tempo no Japão e importada para os Estados Unidos e Europa, anos mais tarde (Lucotte, 1982), encontrando-se classificada da seguinte forma (www.itis.gov, 2018):
Reino – Animal Filo – Chordata Classe – Aves Ordem – Galliformes Família – Phasianidae Género – Coturnix
Espécie – Coturnix japonica
Na literatura, tem havido considerável confusão e controvérsia relativamente à categorização taxonómica da codorniz japonesa, persistindo uma variedade de nomes vernaculares. A Coturnix japonica tem sido considerada, como uma subespécie da espécie Coturnix coturnix detendo, a designação científica de Coturnix coturnix japonica (Wetmore, 1952). Contudo, evidências taxonómicas sobre diferenças na vocalização e inviabilidade híbrida sugerem que a codorniz europeia e a codorniz japonesa são espécies diferentes (Baker & Manwell, 1967). Desta forma, a designação científica para a codorniz japonesa adoptada neste trabalho será Coturnix japonica.
3.3.2. Características
A C. japonica é uma ave terrestre omnívora que habita preferencialmente em áreas com pastagens, terras cultivadas e encostas secas localizadas perto de água. A sua dieta baseia-se numa ampla variedade de matéria vegetal e invertebrados terrestres, nomeadamente gastrópodes, moluscos e aracnídeos (Combreau & Guyomarc`h, 1989). Esta ave reproduz-se no leste da Ásia, incluindo o norte da Mongólia, a ilha de Sakhalin no nordeste da China, o Japão, a Coreia do Norte, a Coreia do Sul e a Indochina (del Hoyo, Elliot & Sargatal, 1994). A maioria das aves migra para o sul da China, Laos, Vietnam, Camboja, Mianmar e nordeste da Índia, ficando algumas residentes no Japão. No entanto, em 1921 algumas populações desta ave foram introduzidas no Hawai e em Itália (del Hoyo et al., 1994; Mills, Crawford, Domjan & Faure, 1997). Em 1994 del Hoyo e a sua equipa consideraram, que a espécie Coturnix japonica era bastante comum na China, contudo foram registados declínios de populações da mesma espécie em Laos (Duckworth, 2009) e no Japão (Okuyama, 2004).
12 3.3.3. Domesticação
Os primeiros registos de C. japonica silvática foram datados desde do século XII no Japão, tendo a espécie sido domesticada no século XI, ou importada da China na sua forma domesticada no século XI (Howes, 1964). Esta codorniz foi originalmente mantida devido ao seu canto, as linhas seleccionadas com determinados tipos de canto particulares foram criados para competir em concursos de músicas (Wakasugi, 1984). Contudo, durante a Segunda Guerra Mundial todas as linhas genéticas seleccionadas da C. japonica foram perdidas. Após a guerra, a indústria de codornizes japonesas foi reconstruída com base nas restantes codornizes existentes em ambiente doméstico, a partir de codornizes capturadas no estado selvagem e de aves provenientes da Coreia, China e Taiwan (Howes, 1964; Wasasugi, 1984).
3.4. Características comuns de C. coturnix e C. japonica 3.4.1. Dimorfismo sexual
A distinção entre machos e fêmeas, no seu habitat natural e por visualização à distância não mostrou evidência clara de dimorfismo sexual, com excepção de episódios fortuitos e raros de se observar, nomeadamente a presença da fêmea no choco ou a mesma seguida das suas crias (Sousa, 2014). As aves quando manuseadas apresentam dimorfismo sexual mais acentuado e fácil de reconhecer, nas codornizes, as fêmeas são mais pesadas que os machos, diferença que ocorre por volta da terceira e quarta semana de idade, sendo atribuída ao peso dos ovários e do fígado (Oguz, Altan & Kirkpinar, 1996; Preez & Sales, 1997). A forma selvagem e a forma doméstica da codorniz japonesa não apresentam diferenças no seu cariótipo, a sua maior diferença, baseia-se principalmente pelo facto destas últimas serem mais pesadas e apresentarem uma maior produção de ovos (Mills et al., 1997).
Em ambos os sexos, a plumagem do corpo é predominantemente de cor castanha escura (Wetherbee, 1961). Para a identificação do sexo devem ser julgados dois aspectos essenciais, observação do peito e observação da garganta (Quadro 3).
As fêmeas apresentam penas de cor pálida no peito, salpicadas de manchas escuras. Os machos expõem penas de cor vermelho ferrugem escuras uniformes e pigmentação de cor similar nas bochechas, que é ausente nas fêmeas (Rodriguez-Teijeiro & Puigcerver, 2010).
Macho Fêmea
Peito Sem pintas Mosqueado
Cor penas do peito Vermelho ferrugem Pálida
Âncora na garganta Presente Ausente
__________________________
13
Revisão bibliográfica
Os indivíduos do sexo masculino são identificados pela presença na região da garganta, de uma mancha escura, longitudinal que se prolonga para ambos os lados com idade, figurando uma âncora. Durante o inverno, os machos desenvolvem um colarinho branco e geralmente têm uma plumagem mais clara do que durante o verão. Esta alteração é independente da testosterona, estudos elaborados por Urbanski em 1984 demonstraram que machos castrados desenvolveram a cor das penas típica do verão. As fêmeas ovariectomizadas em idade jovem desenvolveram a plumagem típica dos machos (Gibson, Follett & Gledhill, 1975).
Durante a migração outonal, o rácio de sexo entre os animais é equilibrado, contrariamente ao que acontece na época de acasalamento, em que a proporção de machos é favorecida (Schleidt, 1983; Rodríguez-Teijeiro et al., 1992).
3.4.2. Distinção jovem/adulto
As proporções de idade dos vários animais, entre as diversas localidades têm-se mantido constantes ao longo dos anos. O rácio da idade mostra, de uma forma global, diferenças interanuais sendo os indivíduos jovens representantes de 44% das peças caçadas (Rodríguez-Teijeiro et al., 1996).
A distinção de animais jovens e animais adultos, por simples observação das aves no campo, só será possível com idades inferiores a 60 dias de vida, a partir deste tempo o aspecto exterior é muito semelhante em todas as idades (Rodríguez-Teijeiro et al., 1996). Ao manusear os indivíduos é possível fazer uma boa avaliação da sua idade, de acordo com algumas características anatómicas, estando estas dependentes da cor da barra supraciliar, da íris e da base da comissura do bico (Quadro 4) (Fontoura & Gonçalves, 1993).
Jovem Adulto
Barra supraciliar Amarela Creme ou branca
Íris Esverdeada Castanha
Base da comissura do bico Rosada /Violácea Negra
Quadro 4 - Distinção entre animais jovens e adultos (adaptado de Fontoura & Gonçalves,
14 3.4.3. Hibridismo
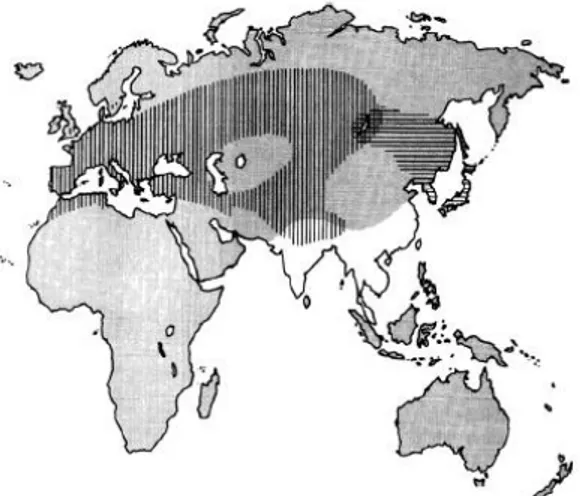
As duas espécies de codorniz em estudo, a C. japonica e C. coturnix, apresentam similaridade morfológica razão pela qual são facilmente confundidas, principalmente onde as áreas de distribuição se contactam. A distribuição destas duas espécies é alopátrica, existindo uma área simpátrica na região Baikal na Rússia e na região Kentei na Mongólia (Fig. 5) (Puigcerver et al., 2007).
Quando se trata de aves no estado selvagem, estas apenas se distinguem pela vocalização dos machos e pelo maior volume corporal da codorniz japonesa. Em animais criados em cativeiro, a sua diferenciação é feita com base na dificuldade que a codorniz japonesa tem em levantar voo (Puigcerver, Vinyoles & Rodríguez-Teijeiro, 2007). Para contornar esta dificuldade na distinção das duas espécies ou de híbridos, Barilani e os seus colaboradores em 2005 recorreram à utilização de marcadores genéticos, tendo sido evidenciado apenas uma pequena diferença entre as duas espécies. As populações naturais da Coturnix coturnix podem cruzar-se na natureza, com indivíduos que não sejam nativos (Coturnix japonica ou híbridos), como resultado do repovoamento de caça. Esta situação poderá constituir, a longo prazo, uma diminuição migratória da codorniz, variação genética e o aparecimento de novas espécies (Puigcerver et al., 2007). O cruzamento inter-espécies pode ser controverso, alguns autores consideram que pode representar um importante mecanismo de evolução, enquanto que outros se opõem a esta ideia, refutando que a hibridização reduz a sua aptidão e que as espécies indígenas afastavam as nativas (Mayr, 1992; Mallet, 2005). O hibridismo pode conduzir a uma perturbação dos animais à adaptação às condições locais, sendo considerado uma das principais causas pelo aumento de aves que não migram para o continente africano no inverno (Huisman, 2006).
Fig. 5- Áreas de reprodução da codorniz europeia
(linhas verticais) e da codorniz japonesa (linhas horizontais), mostrando a área de sobreposição (adaptado de Puigcerver et al., 2007).
__________________________
15
Revisão bibliográfica
As codornizes criadas em cativeiro (Coturnix japonica e híbridos) foram em tempos, libertadas para actividade de carácter venatório (Amaral et al., 2007; Puigcerver, Sardà-Palomera & Rodríguez-Teijeiro, 2012). Segundo Puigcerver e a sua equipa em 2007, registaram a largada em Espanha, de mais de um milhão de aves criadas em cativeiro. De acordo com este registo, o cruzamento entre as duas espécies e híbridos ocorreram com frequência em determinadas áreas de Portugal e Espanha. Registos observados em 2012 por Sanchez-Donoso e seus colaboradores verificaram, que 85% das codornizes soltas não pertenciam à espécie Coturnix coturnix, mas sim a aves com origem em codornizes japonesas ou híbridas. A longo prazo esta situação pode pôr em perigo a sobrevivência da codorniz comum, ameaçando a preservação das espécies migratórias nativas. Segundo as conclusões obtidas por Puigcerver et al. em 2002 e em 2007, a conservação da codorniz comum não constituirá um problema, devido à codorniz japonesa ou híbridos apresentarem uma alta taxa de mortalidade, a sua má adaptação ao estado selvagem, a sua sensibilidade ao frio, a sua ineficiência na defesa contra os predadores, a sua fraca capacidade para procurar e seleccionar alimentos e devido às políticas de repovoamento implementadas. 3.4.4. Vocalização
As codornizes ao contrário das outras espécies galiformes, não revelam um conjunto de indícios que demonstrem a sua presença, nomeadamente a deposição de fezes, pegadas no pó dos caminhos ou na terra húmida de alguns charcos. Os primeiros cantos ocorrem entre a quinta e a sexta semana de idade (Wetherbee, 1961). O canto trissilábico é inequívoco dos machos durante a época nupcial e normalmente ocorre entre Março e Agosto (Moreau, 1951). O canto dos machos é apenas anunciador da sua presença para as fêmeas que estejam prontas acasalar e não é territorial, ao contrário do que acontece nas outras espécies avícolas (Puicerver et al., 2000).
3.5. Potencial económico e de investigação 3.5.1. Potencial económico/produtivo
A codorniz uma pequena espécie aviária doméstica, diferencia-se das outras espécies avícolas pelo seu rápido crescimento, precoce maturidade sexual, geração de curto intervalo de tempo e uma maior eficiência de conversão de alimentos em produção de carne e ovos. Estas características assumem, a codorniz como um recurso importante a nível mundial na produção de carne e ovos (Thiyagasundaram, 1989; Panda & Singh, 1990).
A produção comercial de codorniz japonesa, para a obtenção de carne e ovos na Europa, Ásia, Índia e América do Norte têm uma importância económica crescente, particularmente para um determinado nicho de mercado, embora este crescimento tenha sido desigual nos diversos países (Baumgartner, 1994; Jones, Hughes & Hale, 1979; Minvielle, 2004; Costa et
16
al., 2008; Narinc et al., 2013). O Japão apresenta uma alta produção de ovos, enquanto Espanha e França apresentam uma significante produção de carne, no entanto, verifica-se uma baixa produção ou mesmo nula na Holanda, Alemanha e Reino Unido (Minvielle, Hirigoyen & Boulay, 1999). Na Ásia, para além dos produtos frescos de codornizes, a sua preparação em conservas têm sido comercializadas e ainda foram desenvolvidas tecnologias para fazer ovos em conserva (Minvielle, 2004).
Estas aves quando comparados com outras aves domésticas são eficientes produtores de ovos. Com a introdução de linhas genéticas especializadas principalmente na produção de carne ou ovos, o peso corporal das codornizes aumentou substancialmente, podendo o seu peso passar de 100 gramas para 300 gramas (Minvielle, 2004). A inclusão destas linhagens genéticas nas aves afecta, significativamente a conversão alimentar, a eficiência alimentar e o ganho de peso (Panda & Singh, 1990; Ardiningsasi, Maeda & Okamoto, 1992; Marks, 1993).
3.5.1.1. Produção de carne
Nos últimos anos, a criação de aves de capoeira foi um dos sectores que sofreu maior desenvolvimento na criação de animais. Adjudicado ao aumento de produção de ovos e carne, foram feitas tentativas de diversificação da mesma. É cada vez mais comum, a presença de carne de avestruz, faisão e codorniz em grandes lojas de distribuição. A produção de codorniz ainda não é um ramo estabelecido, mas não obstante ocupa um lugar relevante na criação de aves e contribui para a variedade na produção de carne de aves (Genchev, Mihaylova, Ribarski, Pavlov & Kabakchiev, 2008).
O regime intensivo da produção de carne de codorniz baseia-se nas suas propriedades reprodutivas, rápido ciclo de crescimento e desenvolvimento, alcance da sua plenitude sexual e reprodutora aos 45 dias e a grande resistência desta espécie animal a doenças contagiosas (Lofti, Zerehdaran & AhaniAzari, 2011). A particular capacidade reprodutiva destas aves torna-a, num recurso importante para a produção de alimentos, uma codorniz é capaz de originar mais de quatrocentos descendentes ao ano. Países como a China, Índia e o Japão têm impulsionado a indústria da produção da carne de codorniz, com recurso aos seus avanços científicos alcançando produções surpreendentes (Pérez, 1978).
A aparência/cor e a textura/tenrura da carne de aves são as características mais importantes, sendo estas que mais influenciam a selecção inicial e a satisfação final dos consumidores (Fletcher, Qia & Smith, 2000; Genchev et al., 2008). Os estudos realizados por Flectcher e a sua equipa em 2000 concluíram, que a cor de carne de aves é afectada por diversos factores, sendo estes o teor do pigmento heme presente na carne, o pré e o pós abate e as variáveis de abate. Muitos estudos foram realizados com o intuito de melhorar a composição da carcaça (Lofti et al., 2011) e a conversão alimentar (Khaldari et al., 2010; Varkoohi et al., 2010).
__________________________
17
Revisão bibliográfica
A qualidade da carne está intimamente relacionada, com a descida do pH da carne post-mortem. O rápido declínio do pH post-mortem resulta, numa carne PSE e com reduzida capacidade de retenção de água (Pietrzak, Greaser & Sosnicki, 1997; Owens et al., 2000). A presença de um pH elevado no post-mortem dá origem, a uma carne DFD e com baixa qualidade de armazenamento (Allen, Russel & Fletcher, 1997). Um estudo efectuado em codornizes demonstrou uma heritabilidade moderada a elevada entre os indicadores finais de pH e a cor da carne (Oguz et al., 2004).
A produção de codornizes a nível mundial representa, um pequeno sector no grupo avícola mas revela uma importância acrescida num determinado nicho de mercado e com um impacto significante em determinados países, existindo ainda grandes lacunas que podem melhorar a sua produção (Minvielle, 2004; Móri et al., 2005).
De acordo com Barros (comunicação pessoal, Abril 4, 2019), e no que respeita a produção de carne, esta é feita com aves de ambos os sexos e a eficiência produtiva da codorniz assentam num ciclo de vida curto (35 dias), um ganho médio diário (GMD) de 6,7 g/dia e um índice de conversão de 2,59. As codornizes usadas na produção de carne são abatidas com um peso-vivo a rondar as 220 g e apresentam um rendimento de carcaça de 65,2%.
3.5.1.2. Produção de ovos
A criação de codornizes orientada para a produção de ovos é um aspecto de grande interesse, oferecendo perspectivas tão importantes, quanto a criação destas aves para a produção de carne (Wilson et al., 1961). O fundamento deste interesse prende-se com o seu valor nutritivo, com a digestibilidade e palatibilidade do ovo de codorniz (Sundaram, 1988; Narinc et al., 2010). A produção de ovos de codorniz não se destina única e exclusivamente ao consumo de ovos frescos, como também apresentam interesse para a indústria conserveira (Pérez, 1978).
Os ovos de codorniz são caracterizados pelo seu pequeno tamanho e pela panóplia de padrões de cores de casca, variando de castanho escuro a azul, branco ou salpicado (Panda & Singh, 1990). A produção de ovos de codorniz apresenta valores muito variáveis, as poedeiras são capazes de pôr dois ovos por dia, com um intervalo de 12 horas, podendo alcançar-se os 500 ovos por ano. A qualidade dos ovos de codorniz é influenciada por factores genéticos e ambientais, durante o período de postura (Pérez, 1978).
O peso do ovo varia durante a estação do ano, sendo estes maiores no inverno e mais pequenos no verão (Panda & Sigh, 1987), sendo justificado pela reduzida ingestão de alimento, com temperaturas iguais ou superiores a 30º C, o que condiciona menor disponibilidade de proteína para a formação do albúmen (Marsden et al., 1987).
A selecção de codornizes poedeiras baseia-se, no peso dos animais no início da postura e o peso da produção de ovos. As fêmeas que normalmente pesam mais de 90 gramas aos 30
18
dias de idade e iniciam a sua postura até aos 40 dias de idade são consideradas excelentes poedeiras (Pérez, 1978).
Para a obtenção de linhas poedeiras de alta produção foram realizados, cruzamentos entre as espécies Coturnix coturnix e a Coturnix japonica, melhorando a resistência às condições ambientais e às doenças (Narinc et al., 2010). De acordo com Barros(comunicação pessoal, Abril 4, 2019), na produção de ovos para consumo (ovos não fecundados) é usada uma genética selecionada para o efeito (130 g de média e menor consumo de alimento), a postura é iniciada aos 42 dias de vida e mantem-se ao longo de 364 dias (52 semanas), representando um único ciclo de postura. No final da postura, as aves são abatidas e a carcaça destas aves é exportada.
3.5.1.3. Comparação da codorniz com outras aves de capoeira usadas tradicionalmente para a produção de carne e ovo
No que respeita à produção de carne, a dimensão da ave e por consequência da respectiva carcaça não é representativa do seu potencial produtivo. A comparação da carne de codorniz com as carnes de frango e pato revela que esta apresenta um maior teor de proteína (19,6 g/100 g de carne) que o frango (18,6 g/100 g de carne) e o pato (11,5 g/100 g de carne), apresentando um teor de lípidos totais mais baixo (12,1 g/100 g de carne) que o frango (15,1 g/100 g de carne) e que o pato (39,3 g/100 g de carne), o que faz da codorniz uma fonte dietética de proteína pois a sua carne apresenta uma carga calórica inferior (804kJ), comparativamente com a carne de frango (900 kJ) e pato (1691 kJ). A carne de codorniz apresenta maior concentração de vitamina A, vitamina C, vitamina B2, Vitamina B3, Vitamina B6, Vitamina B12, ferro, magnésio, cálcio, fósforo, potássio, cobre, zinco e selénio (Ioniţă, Popescu-Micloşanu, Roibu & Custură, 2010).
O ovo de codorniz apresenta em média 17,5% do peso do ovo de galinha (11 g versus 63 g, para o ovo de codorniz e galinha, respectivamente) e o peso da gema representa na codorniz 19,3% da gema do ovo de galinha (Bertechini, 2012). No que respeita à composição do ovo, a gema do ovo de codorniz apresenta uma concentração mais elevada de ácido linolénico e de ácido docosahexaenóico, uma concentração inferior de ácido linoleico, apresentando por isso um rácio n-6/n-3 mais favorável que a gema do ovo de galinha e uma concentração de colesterol idêntica (Kazmierska, Jaroz, Korzeniowska, Trziszka & Dobrzanski, 2005).
Em termos produtivos, a precocidade da codorniz, com o início da postura a começar aos 42 dias (enquanto as galinhas poedeiras iniciam a sua postura, em média, pelo dia 126) e o seu menor peso corporal no início da postura (120 versus 1350 g) implica um menor custo em alimentação durante o período de desenvolvimento (418 versus 6500 g). Contudo, uma mortalidade elevada durante o período de postura diminui a eficiência produtiva da codorniz.
__________________________
19
Revisão bibliográfica
A galinha apresenta uma maior eficiência alimentar que a codorniz, o que resulta num menor custo de produção quando se considera a produção de 1 kg de ovo, no entanto, se considerarmos o custo de produção de uma dúzia de ovos, a codorniz revela ser mais lucrativa (Bertechini, 2012).
3.5.1.4. Produção da codorniz como animal de caça e desporto
A coturnicultura, além de um meio de produção de carne e ovos, a codorniz comum pode ainda ser usada para repovoamento da caça (Pérez, 1978).
A actividade cinegética é mundialmente reconhecida, tendo um valor turístico e económico elevado. Em Portugal este sector está em franca expansão, o seu potencial cinegético é reconhecido (Erena, 1993). No Alentejo existem 863 zonas de caça associativa, 631 turísticas e 318 municipais, que se distribuem por uma área correspondente a 77% da área total do Alentejo (Sousa, 2014). A caça é uma actividade cada vez mais procurada pelas populações dos grandes centros urbanos, sendo desta forma apontada como uma estratégia produtiva e económica de diversificação agrária de grande importância. Dentre as seis espécies cinegéticas mais importantes de toda a Península Ibérica, veado (Cervus elaphus), javali (Sus scrofa), lebre (Lepus granatensis), coelho (Oryctogalus cuniculus), codorniz (Coturnix coturnix) e a perdiz (Alectoris rufa), a codorniz em 1990 ocupava o terceiro lugar das espécies mais caçadas, tornando-a numa espécie de indiscutível valor cinegético e económico (Rodríguez-Teijeiro et al., 1996).
A codorniz, C. japonica, é uma espécie avícola doméstica e devido à sua anatomia tanto externa (desenvolvimento das asas e penas essenciais para o voo) como interna (desenvolvimento dos sacos aéreos) demonstram, que esta não dispõe de possibilidades como ave de voo, o que a torna inabilitada para integrar o repovoamento de animais aptos, para o desporto de caça (Puigcerver et al., 2007). A codorniz de caça caracteriza-se por um arranque rápido, elevação do solo quase vertical e voo rápido mas relativamente curto. Esta forma de voo caracteriza a Coturnix coturnix, uma autêntica espécie cinegética que proporciona um alvo fácil para o caçador (Pérez, 1978).
Em Portugal não é permitida a utilização de codornizes da espécie Coturnix japonica e cruzamentos derivados da mesma, para fins cinegéticos, de repovoação ou a sua utilização em campos de treino de caça ao abrigo do Decreto-lei nº 2/2011. O mesmo se verificou em França desde 2002 e em Espanha desde 2007 (Sousa, 2014).
A redução das extensões cultivares provocada, pelo desaparecimento da agricultura extensiva que vai sendo substituída pela agricultura industrial e pela repovoação florestal, constituem a razão pelo qual a caça vai desaparecendo na sua modalidade livre, e vai sendo substituída por caça controlada de reservas ou cotadas, onde se produzem as espécies cinegéticas mais convenientes (Pérez, 1978; Amaral et al., 2007; Puigcerver et al., 2012). A Península Ibérica graças à sua luminosidade particular, reduzida humidade e
20
ausência de núcleos industriais de grande dimensão, reúne as condições ideais, para o repovoamento de codornizes (Rodríguez-Teijeiro et al., 1996).
3.5.1.5. Produção da codorniz como animal ornamental
A codorniz como animal ornamental também tem interesse, sobretudo quando se trata de parques e jardins recreativos. A codorniz japonesa e os animais resultantes do cruzamento entre esta espécie e a codorniz comum adaptam-se perfeitamente a estes ambientes. Por outro lado, o seu carácter dócil permite a sua adaptação a recintos familiares e jaulas, que para além do seu valor ornamental e canto, podem também constituir um rendimento de uso familiar pela sua postura de ovos (Pérez, 1978).
3.5.1.6. Comercialização
A comercialização deve ser precedida por um estudo da carne, que engloba a avaliação do rendimento de carcaça da codorniz, sua qualidade nutricional e gastronómica, e conhecer os factores que modificam a qualidade da carne (Pérez, 1978). Em geral, o rendimento cárnico da codorniz corresponde entre 70 a 75 gramas, por cada 100 gramas de carcaça (Móri et al., 2005; Khaldari et al., 2010). A carne das codornizes criadas em cativeiro caracteriza-se pela sua tenrura, apetecibilidade, suculência e digestibilidade (Móri et al., 2005).
3.5.2. Potencial de investigação
A Coturnix japonica é uma espécie comumente utilizada, como modelo ou fins comparativos para estudos de comportamento, bases genéticas, neurofisiológicas e neuroendócrinas (Sato, Sato & Ino, 1984). O seu pequeno porte, as baixas exigências económicas na sua criação e a sua alta adaptabilidade a um amplo espectro de condições de criação, são características que justificam a sua utilização como animal de laboratório (Padgett & Ivey, 1959; Reese & Reese, 1962, Mills et al., 1997). Esta espécie é utilizada como animal de laboratório, desde 1945 no desenvolvimento de vacinas nomeadamente para a Doença de Newcastle (Padgett & Ivey, 1959; Islam et al, 1994; Lima et al., 2004). Muitos genes da C. japonica têm particular interesse para a medicina humana, o gene albino presente nesta espécie induz glaucoma na codorniz à semelhança com o glaucoma humano (Dkhissi et al., 1999). A codorniz japonesa foi escolhida em 2004, como o animal a utilizar num programa de pesquisa espacial com base na lua (Young, 1968; Minvielle, 2004).
__________________________
21
Revisão bibliográfica
Fig. 6- Produtores mundiais de carne de codorniz (adaptado de
Poultry World, 2018).
3.6. Enquadramento Geral da Produção
A produção de codornizes tem como objectivo final a obtenção de animais saudáveis, com peso final ao abate adequado às exigências do consumidor e enquadradas num custo orçamental justificável (Wilson et al., 1961).
As codornizes criadas para a produção de carne são criadas até aos 35 dias de idade em pavilhões climatizados, usando-se as aparas de madeira ou a casca de arroz para a “cama” das codornizes. A climatização dos pavilhões é essencial pois durante os três primeiros dias após a eclosão necessitam de uma temperatura ambiente de 35º C, devendo a temperatura ambiente sofrer uma progressiva redução ao longo do tempo (0.5º C/dia), a partir do 28º dia de vida a temperatura deverá estabilizar entre os 21 e os 23º C (Shanawany, 1994).
Enquanto a produção de codorniz para carne é realizada em pavilhão, para a produção de ovo, as codornizes estão em jaulas, podendo uma jaula albergar 60 codornizes.
Os maiores produtores mundiais de carne de codorniz (Fig. 6) são a China (168 000 toneladas), Espanha (9 300 toneladas), França (8 200 toneladas) seguidas de Itália, Estados Unidos da América, Austrália, Portugal, Brasil e Japão (Minvielle, 2004; Sartowska, Korwin-Kossakowska, Polawska, Lipinska & Sender, 2014).
Na Europa, os maiores consumidores desta carne é a Itália, França e Espanha (Tserveni-Gousi & Yannakopoulos, 1986). Em Portugal, registou-se um maior volume de abate de galináceos (+15,3%) em Janeiro de 2016 relativamente ao mesmo período do ano anterior, embora as codornizes tenham registado um decréscimo de 11,7% (Quadro 5). Relativamente às cabeças abatidas, verificaram-se igualmente acréscimos no número de galináceos (+8,9%) sendo que as codornizes registaram um decréscimo de 7,2% (Instituto Nacional de Estatística, INE, 2018).
22
A criação de codornizes restringe-se a Portugal continental e, em específico, à área metropolitana de Lisboa, sendo que o número de cabeças e o número de toneladas se têm mantido relativamente constantes (Quadro 6).
3.7. Composição da carne
A carne desempenha um papel muito importante na nutrição humana, graças ao seu valor nutritivo. A importância da carne é avaliada pelo teor e relação dos seus principais componentes químicos como: a proteína, a gordura, os minerais e as vitaminas (Genchev et al., 2008).
O aumento do interesse do consumidor pela saúde tem sido revelado, pela prática de uma alimentação e um estilo de vida saudável (Mastrosimone, Marsico & De-Gennaro, 1998; Sadler, 2004; Karakok et al., 2010). Esta preocupação dos consumidores, pelos alimentos saudáveis e/ou biológicos mostrou um aumento tendencial do consumo de carne de caça. Este tipo de carne apresenta um valor nutricional benéfico para a saúde, principalmente quando comparado com outros tipos de carne (Amaral et al., 2015). O típico paladar, as propriedades dietéticas e o alto valor nutritivo da carne de codorniz são fundamentais para o crescente interesse dos consumidores por este produto (Genchev et al., 2008; Kokosszynski, Bernacki, Korytkowska, Wilkanowska & Frieske, 2013). A carne de codorniz é
Espécie Codornizes
Ano Nº Cabeças Toneladas
Portugal
2012 9 769 109 1 370
2013 8 828 443 1 236
2014 10 210 208 1 459
Quadro 6 - Efectivo de codornizes em cabeças e toneladas em Portugal
(adaptado de Instituto Nacional de Estatística, 2016).
Quadro 5 - Aves abatidas e aprovadas para consumo público (adaptado de Instituto Nacional
de Estatística, 2016).