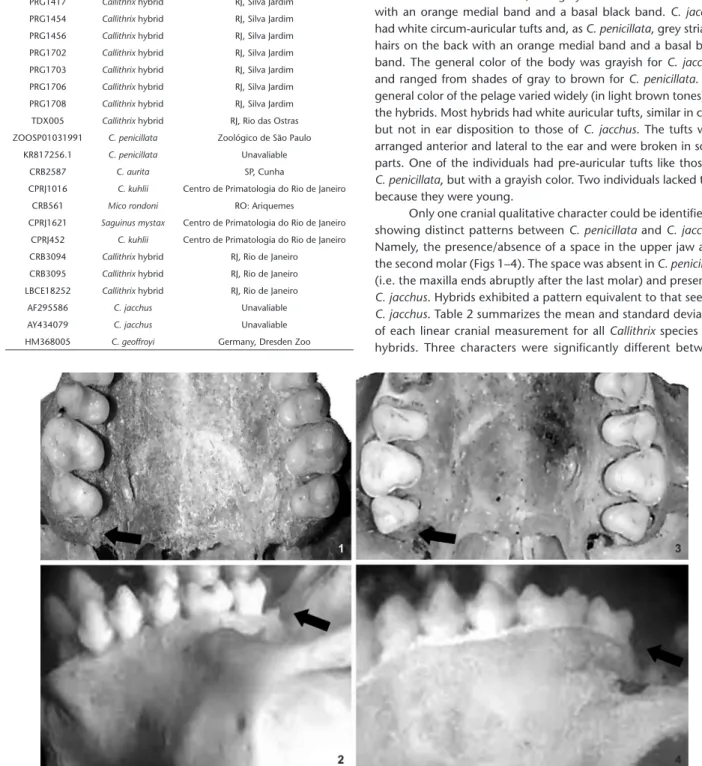
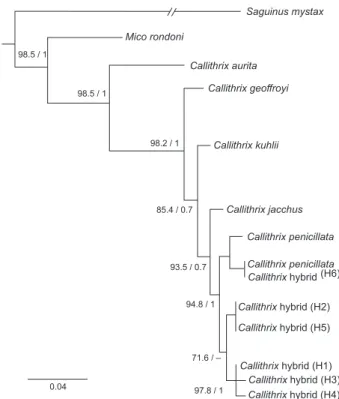
Morphological and genetic diversity in Callithrix hybrids in an anthropogenic area in southeastern Brazil (Primates: Cebidae: Callitrichinae)
Texto
Imagem

Documentos relacionados
(2004) assessed the reaction of 20 melon hybrids under field conditions in municipalities in the States of Rio Grande do Norte and Ceará and observed that the hybrids
The agronomic traits FGW and TDM had higher individual heritability estimates amongst all traits evaluated considering either the joint analysis or the seasons separately, with
In the present study, the objectives were to identify promising popcorn lines and hybrids for genetic resistance to tropical rust ( P. zeae ) in diallel crosses of inbred lines
The high genetic variability observed in the testcross hybrids, the low frequency of favorable alleles and the high concordance in lines classification with the average of other
ABSTRACT: This study aimed to evaluate the inbreeding depression and average genetic components in seven hybrids and two open pollinated cultivars for green corn production, as
In this work, inter-simple sequence repeat markers were used to determine the genetic variability and the loci segregation profiles of 84 strawberry hybrids obtained from a
et al ., 2010; Kishor et al ., 2013; Kumar & Reddy, 2016b).Thus, the evaluation of genetic diversity in this stage helps in the selection of pre-commercial hybrids and in
The purpose of this study was to evaluate hybrids obtained between temperate × tropical elite maize lines and to determine the genetic diversity and the population differentiation
