A new genus and species of leaf miner (Lepidoptera, Gracillariidae) for Chile associated to the native tree Lithraea caustica
Texto
Imagem

Documentos relacionados
A metodologia de delimitação automática das áreas de preservação permanente (APPs) tendo como referência legal o Código Florestal, Lei Federal 4.771/65, a Resolução
Quanto ao tipo de mapa, existem dois que são os principais para deficientes visuais que dependem do grau de deficiência do indivíduo, são: mapas coloridos para pessoas
de plântulas, condutividade elétrica, envelhecimento artificial e emergência das plântulas em campo, uti I izando-se doze lotes de sementes de ervilha, pertencentes a dois
18.14.23.5 A cabina do elevador automático de passageiros deve ter iluminação e ventilação natural ou artificial durante o uso e indicação do número máximo de passageiros e
incorporação em subprodutos gerados nos processos industriais, seja na fase sólida, líquida ou gasosa. A mesma fornece os requisitos para o aproveitamento de água de chuva
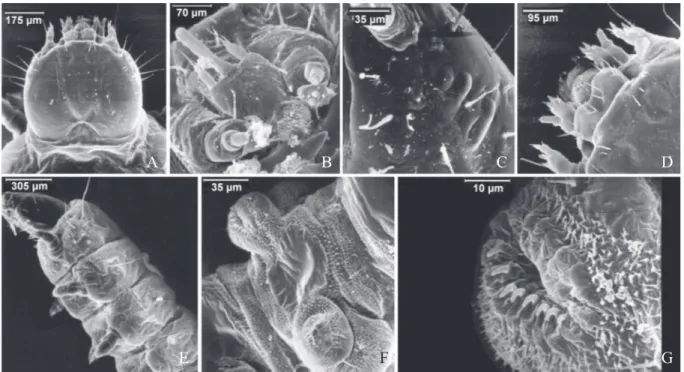
Dorsal region with blurred light area; dark patches of pigmentation absent along the body; ventral region light colored; two light colored stripes present on the caudal- fin;
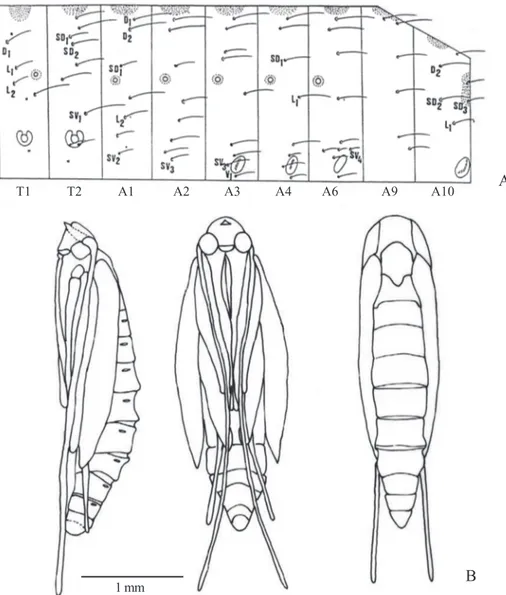
differences. Maxillary palpi with dark brown and yellowish scales; base of palpomere 2 with erect scales, basal 0.5 with patch of yellowish scales, a few yellowish scales at apex
It did not present dorsal or ventral spots, with exception of the tail which showed a few dark spots in relation to background: shades of beige (123A) and/or brown (223A,
