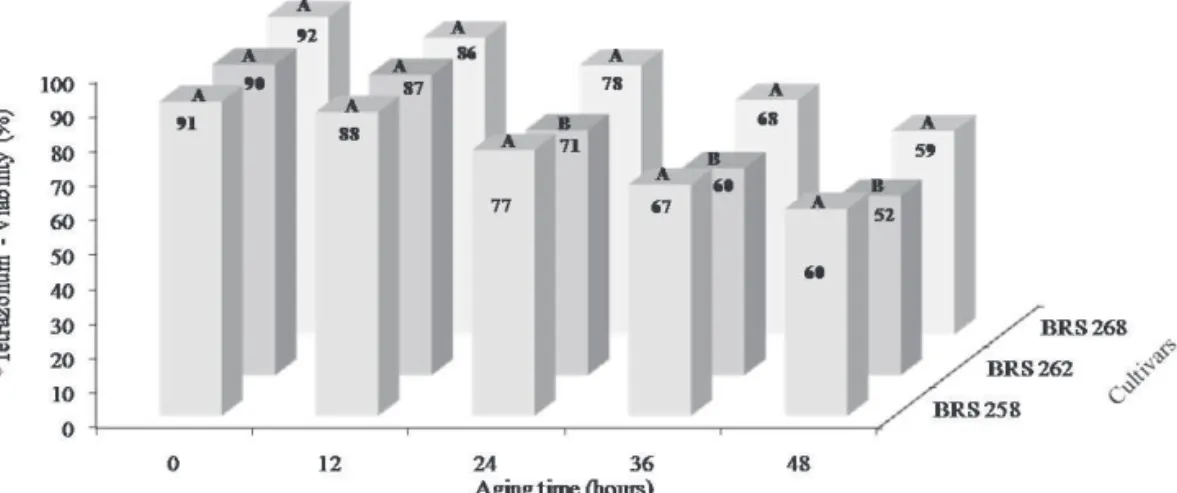
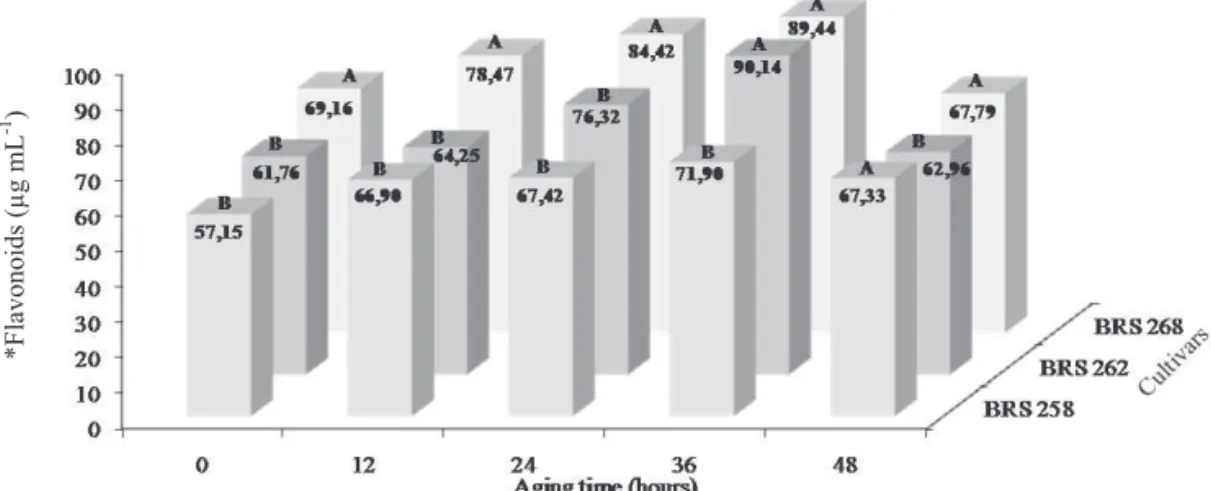
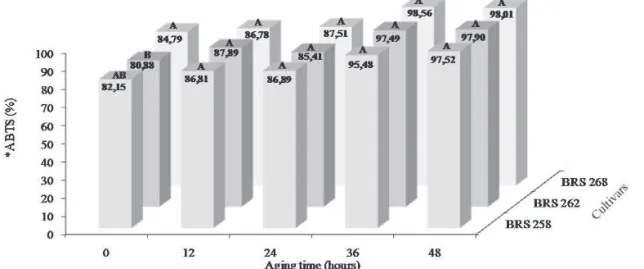
Physiological quality, content and activity of antioxidants in soybean seeds artificially aged
Texto
Imagem


Documentos relacionados
Esta pesquisa foi norteada pela revisão bibliográfica, com base em artigos e livros e pesquisas, acerca do assunto estudado: a queda da obrigatoriedade do diploma
The aim of this study was to evaluate the effect of foliar application of manganese, with different doses and stages of application, on the physiological quality of soybean seeds
Méd: de *******?, vê só ******, essa medicação que a gente vai te dar agora, que é um comprimido de dose única, tu vai pegar aqui no posto, o outro comprimido que é para
Tendo compreendido que a ACV pode auxiliar no processo de diminuição dos impactos da CPIC desde a fase de extração de matérias primas, passando pelos processos
Outra importante ação realizada pela escola foram os momentos de convivência, nos quais alunos de escolas de ensino fundamental eram convidados a fazer uma visita à
α -tocopherol contents increased (25%) for the artificially aged soybean seeds, and at 180 days of storage, the difference in α -tocopherol levels between natural and
To evaluate the effect of preconditioning of maize seeds in ozonated water on seed physiological quality, germination test (GT) was conducted and the following variables were
Ao considerar n números fuzzy, deve-se usar a adição desses números e dividí-los por um número real, a partir do que se encontra a média fuzzy BOJADZIEV, 2007 7.2 CÁLCULO
