HISTÓRIA DE VIDA DE
Texto
Imagem




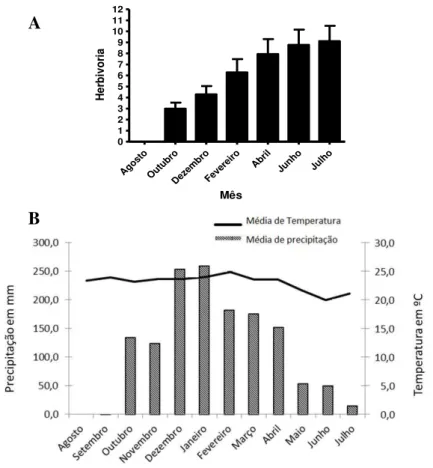




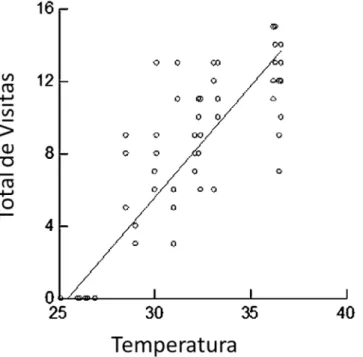
Documentos relacionados
Dentre as principais conclusões tiradas deste trabalho, destacam-se: a seqüência de mobilidade obtida para os metais pesados estudados: Mn2+>Zn2+>Cd2+>Cu2+>Pb2+>Cr3+; apesar dos
O Núcleo de Estudos Dirigidos concebe que para se desenvolver a capacidade de interpretação de textos e domínio de suas informações, permitindo sua transferência para
Foram selecionados um total de 143 prontuários consecutivos de pacientes de uma coorte observacional do Ambulatório de Hepatites Virais do Hospital das Clínicas da UFMG,
O termo extrusão do núcleo pulposo aguda e não compressiva (Enpanc) é usado aqui, pois descreve as principais características da doença e ajuda a
• O material a seguir consiste de adaptações e extensões dos originais gentilmente cedidos pelo
O presente trabalho objetiva apontar algumas influencias da atividade física na promoção da qualidade de vida e bem estar dos portadores do HIV, uma vez que o
Projetar um controlador que seja robusto a falhas no atuador utilizando a teoria de estabilidade de Lyapunov via formulação LMI; Análise e desenvolvimento do
Toque a chave de controle de volume conforme necessário para desligar ou ajustar o volume para músicas e sons da natureza. Quando o produto estiver conectado a um reprodutor de
