I]NWERSIDADE
DE
EVORA
Comunidades
de
aves
em
galerias
ripícolas mediterrânicas
do
§ul
de
Portugal
kilaMagnaGomes DuaÍe
Dissertação para obtenção do Grau
de
Meste
em Gestâo
eConservação
de Recursos
Naturais
Esta dissertação não
inclui
as críticas e zugestões feitas pelojúri.
Orientadora: Prof.
Doutoura Maria Teresa Fe'rreiraÉvona
Março
2009UE
t7t
yt
UNTVERSIDADE DE
EVORA
Comunidades
de
aves
em
galerias
ripícolas mediterrânicas
do
Sul
de
Portugal
Leila
Magrra Gomes Duarte
Dissertação para obtenção do Grau
de
Mestre em Gestão
eConservaçõo
de Recursos
Naturais
Esta dissertação não
inclui
as críticas e sugestões feitas pelojúri.
/Tr
J
ql
Orlentadore: Prof.
Doutoura Maria Teresa FerreiraÉvoru,
Março 20lD
ti
Comunidades
de aves em
galerias
ripícolas mediterrânicas
do
SUI
de
Portugal
Bird
communities in Mediterranean riparian galleries
in
southem Portugal.
Leila
Mapa
GomesDuaÉe
Cerro Álagoa F-2 8200-016
Álbufeira,
e-mail: leilamgd@gmail.comRESt'MO
Conhecer
os
reqúsitos
ambientaispor
parte das
espéciesdemonstou
ser essencial em disciplinas como a ecologia e a biologia da conservação. O presente estudoúsa
estudar as respostas e dependência das comunidades de aves em relação às galeriasripícolas
mediterrânicas. Paratal
utilizaram-se dados recolhidospelo River
tlúitat
Survey (RHS) e por censos por pontos de escuta, em três ribeiras no
sú
de Portugal. Osdados ambientais
foram
estruturadosem
matizcs de
acordocom as
característicasfisicas da ribeira
e
das margens,e
modificações antopogénicas. Enquantoos
dadosbiológicos foram
agrupadosem guildas:
alimentaçãoe
ocupaçãovertical do
habitat('estrato').
Através
de
pnálises canónicas
aos dados estuturados
obtiveram-secorrelações
válidas
ente
as
matizes
ambientaise
as guildas,
nomeadamente paraindivíduos
directamente dependentesda
água
e
planadores C'aéreas'),provando
avalidade da metodologia e o potencial da combinação destas duas técnicas.
Pahvras-chave: Ribeira
de
Seixe, Ribeira
de
Odelouca,
Ribeira
de
Arronches, Comunidades de aves, Galeriaripícol4
River Habitat Survey.ABSTRACT
Understanding species habitat requirements has pmved to be essential
in
ecologyand
conservationbiology. The
presentreport aims
to
examinethe
responses and dependenceof
bird communities in relation to Mediterranean riparian galleries. For thiswe used data collected by River Habitat Survey (RHS) and point count censuses in three
rivers
in
southem Portugal.
The
environmental
data were stuctured
in
matricesaccording
to
physical
characteristicsof
the
steam,
úe
banks
and
anthropogenicmodifications,
whilst biological
data wasgouped into
guilds: foraging and occupation(tstrato").
Through canonisal analysis to structured data we obtainedvalid
correlationsbetween the environmental variables and species guilds,
particularly for
úose
directly
dependent on water and gliders ("aéreas"),
proüng
thevalidity of
the methodology and the potential ofthese two techniques working together.Key-words:
River
Seixe,
River
Odelouca,
River
Arronches,
Bird
Communities,Riparian Gallery, River Habitat Survey.
INTRODUÇÃO
O
sul de
Portugalé
cuacterizadopor um
clima
mediterrânico(Morais eÍ
aI.2004),
e
devidoa um historial
de perturbaçãoantopogénica
e uso desolo
intensivo@écamps et
al.
1988; LepaÍt&
Debussche 1992; Stromberg 1993; Gallego-Femiíndez etal.
1999; Corbacho etaI.2003
in
Agúar
&
Ferreira 2005; Covas&
Blondel
1998), aspaisagens ripícolas mediterrânicas encontam-s€ profundamente fragm.entadas (Jansen
&
Robertson 2001).A
estrutura
das
galerias
ripícolas desempeúa
um
papel
firndamental
na: integridade ecológica dos ecossistemas ripícolas(Aguiar
&
Ferreira 2005), fixação de maÍgens econtolo
de caudal, e actua comofonte
de bio-acumulação (Saraiva 1999), determinando a composição florística e faunística do local (Aguiar&
Ferreira 2005).As
aves repÍesentamum
importante elementode
caracterização das galeriasripícolas,
visto
utilizarem-na para: alimentação, nidificação e corredor de dispersão ou migração (Masonet
a|.2005;
Décampset
al.
1987; Roché 1989;Doherty
&
GÍubb 2002), podendo algumas espécies, ainda" ser utilizadascomo
indicadores ecológicos(Fumess
et
al.
1993),
uma vez que
respondemmais
rapidamentea
perturbações tenestres que precedam as mudançasno
ecossistemaaqúüco
@ryce eraI.2002).
Os corredoresripícolas são
também importantes
para urna série
de
outras
espéciesaúfaunísticas
nos
habitats
adjacentes,funcionando
como
corredor
nâs
palsâgens2005).
Visto
permiürem a circulação entre manchas de habitat para espécies que, por exemplo, não querem sobrevoar espaços abertos, eum
suplemento alimentar (insectosaqúticos
emergentes) para outras (Mason eÍal.
1984&
Iwata etal.
2003, 12 Mason eÍal.
2006). Autores
como
Martin
eÍ
al.
Q006)
demonstramque z)nas
rib€irinhassemelhantes contêm diferentes espécies de aves, devido as influências exercidas pela palsagem
envolvente,
desempenhandoao
mesmo tempo,
um
papel
importante
na ÍeCúação dos números de aves (Forman 1997;Newton
1998; Lowrance&
Crow2002;
Smith
&
Wachob 2006).Dada
a
sua padronizaçãoe
simples aplicação,o
River Habitat
Survey (RHS), introduzido em Portugalno
âmbito daDirectiva
Quadro daÁgua
(DQA),
pÍovou
ser uma ferramenta de trabalho válida para as disciplinas de ecologia aplicada e biologia da conservação; desdeque
haja uma
combinaçãode
variáveis
ambientaise
variáveisbiológicas sobre
um
determinadotmon
§aughan
&
Ormerod
2005).
Trabalhosdesenvolüdos por autores como Buckton
&
Ormerod (1997) e Brewin er aL (1998), sãoprova disto.
No
mesmo contexto,a
Britiú
Trust
for
Omithology
@TO) no
ReinoUnido,
desenvolveuo
Waterways BreedingBird
Survey(WBBS), um
projectopiloto
em
que
utilizam
dados sobreas
comunidadesde
avesem
conjunto
com
algumas características derivadasdo
RHS,
tendo
obtido,
numa fase
preliminar,
corÍelações relevantes§ewson
er a/. 2005).Nesta perspectiv4 o objectivo
primário
deste trabalhofoi
estudar as respostas edependfucia das comunidades de aves em relação às galerias ripícolas mediterrânicas,
analisando parâmetros inerentes
à
ribeirq
ao
habitat
adjacente
e
modificações anhopogénicas, atavés da aplicação do protocolo RHS e métodos pontuais de censo deaves.
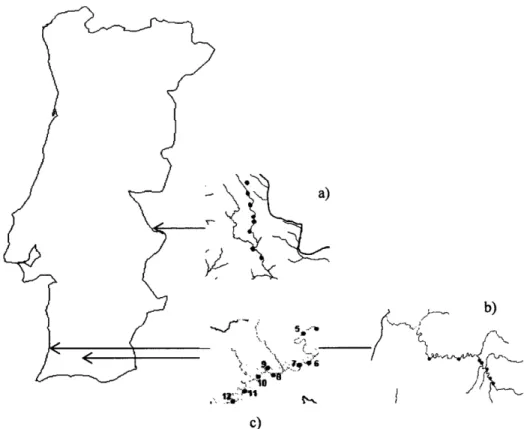
Área
deEstudo
Para amostragem foram seleccionadas
tês
ribeiras nosú
dePortugal
Ribeira de Seixe, Ribeira de Odelouca e Ribeira de Arronches(figura
l),
que, de acordo com aDQA @irectiva
QuadÍo da Água), são
cuacteizadx
pela
elevadaaltitude
e
maior disponibilidade hídrica (quadroI
-
tipo
52) e pelos depósitos calcários (quadroI
-
tipo
\L
a)
b)
c)
Figura 1
-
Localiryáo
geogrrífica dos Eoços, ern que a) Ribeira de Arronches, b) Ribeira de Seixe, e c) Ribeira de Odelouca. Para cada tsoço foram seleccionados, respectivamente: 8, 7 e 8pontos de amostsagem.
QUADRO
I
-
Atibutos climatológioos, geológicos e hidromorfológicos das Ribeiras de Seixe, Odelouca e Aronches (Morais et a/. 2004; Alves et a1.2006; Pais & Cangarato 2001; Hughes e, al.2O02;l).
Ribeira de
Seire
Ribcirr de (NelouceRiboira de Arrorchê§
Distrito Tipologia Altitude (m)
Prrcipitação média anual
(mn)
Temperatura médiar ("C)
Insolaçâo @oras por ano)
Geologia Beja e Faro Sul S2 159
-
280 600 -700 t2-18. 2600-3000 Xistos Faro Sul 54 < 300 700 - 1000 l8-2tr 280G.3000 Xistos e Tuóiditos Portalcgrc Sul S2 300-
800 600-900 l0-18. 2400-2800 Xisto c GranitoA
Ribeira de Seixe (quadrol),
apresenta um regime contínuo de caudal e visto aárea ter sido exposta a fogos, a
matiz
de vegetação envolvente, consiste, na maioria dos casos,numa
camada arbustivade
espécies resistentes, nomeadamentesilvas
(rRuázsulnifulius) e
estevas (CrsÍzs sp.).A
matriz
riprlria é
compostapor um
mosaico deAmieiros (Alnus glutinosa),
Salgueiros(Salix alba,
S.
atrocinerea,
S.
Salvifolia),
Freixos
(Frarrzus
angustifolia)
e
Choupos (génercPopulus)
(2; Vowles
&
Vowles1994).
A
Ribeira de
Arronches(quadro
l)
encontra-sêna face
sul
da
Serrade
São Mamede, e portanto a vegetação é tipicamente mediterrânica. Há dominância dos xarais mediterrânicos, substitúdos por pequenas hortas e montados de sobro (Quercus suber) e./ou azinho(Quercw rotundifolia),
intercalados por terÍenos de sequeiro, à medida quea
altitude
diminú.
A
matriz ripícola é
caracterizadapela
dominânciade Amieiros
(Alnus
gl*inosa)
(2; Pais&
Cangarato 2001).A
Ribeira
de
Odelouca (quadro
l)
apresenta
regime hidrológico
comsazonalidade tÍpicamente mediterrânica (Ferreira
&
Aguiar
2006).O
coberto vegetal édominado
por
florest4
registando-se
pÍrocessosde
substituição
das
formaçõesmediterrânicas (medronhal
e
montados de sobro),por
exploragões florestaismais
oumenos intensivas de
pinheiro bravo
(Pinuspinaster) e
eucalipto (Eucafitptus sp.).As
comunidadesripícolas
mais
característicassão
os
salgueirais(Salix salvidolia),
os amiars (Álnusglutinosa), os
freiúais
(Fraxinus angustifolia)
e
os oleandros(Neriun
oleander),
ou
sej4
formações vegetaiscom poÍte
arbóreo,arbustivo
ou
herbáceo, consoante a sua idade de formação (3; Hrtghes et al. 2007).A
selecção dostoços foi
baseada na existência de uma distllncia mínima de2km
entre locais (distância
estabelecidapelo
protocolo
RHS)
e
de
acordo
com
a acessibilidade ao local.Variíveis
ambienteis
Para a recolha da. variáveis ambientais
foi
aplicado o protocolo RHS em 2 fasesüstintas:
l)
primavera de 2005na Ribeira de
Odelouca"e 2)
primavera de2006
naRibeira de Arronches
e Ribeira de
Seixe,utilizando
o
formulário
RfVER
HABIAT
SURVEY MEDITERRANEAN
2005 Version. O protocolofoi
aplicado de acordo coma
metodologiadesenvolüda
pela Environment AgencyAgency,
1997;Brewin
eral.
1998), segundo a qual siio avaliados troços deribeira
com uma extensâostandard
de
500m,
a
uma
largurade
50m para
cada margem (RaveneÍ
al.
1998b; Davenportet
al.
2001;
RaveneÍ
al.
2002),e
sãofeitos
registosem
l0
spot-checkseqúdistantes, que incluem informaçito acerca dos atributos fisicos do canal e margens (Raven eÍ
al.
1998a; Davenport etal.
2001: Raven e,a|.2002;
Environment Agency, 2003).Do
mesmo modo faz-seum
registo sweep-up onde se registam todos as outrasvariáveis que
ooorrem
ao
longo
do
troço
de
500m,
e
nurna faixa
de
extensão perpendicular à ribeira de 50m (Brewin eraI.
1998; Fox etal.
1998; Raven er aJ. 1998b; Environment Agency 2003; Vaughan&
Ormerod 2005).Variáveis
biológicasA
amoshagem de aüfaunafoi
realizada em duas fases distintas:l)
na Ribeira de Odelouca emJuúo
e Julho dos anos de 2005 e 2006; e2)
nas ribeiras de Arronches e Seixe em Março elúaio
do ano 2006, em ambos os cas)s nos períodos de nidificação,de forma a abranger o maior número possível de especies nidificantes.
Em
cadaribeira,
foi
realizada uma amostragempontual
segundoBlbby
et al.,
1992,
com duraçãode
l0
minutos, em
cadatroço
RHS,
e
tambémde
duas formasdistintas:
l)
na ribeira de Odelouca" em üês pontos equidistantes 250m, cujolimite
dedisttincia
de
detecçãovariou com
o tipo e
extensãode habitât
associadoà faixa
de galeria ripícola; e 2) nas ribeiras de Seixe e Arronches, nos pontos corÍespondentes aos spot-checks3,6
e 9, com umlimite
de detecçãofixo
de 50m.Em ambos os casos os oensos foram conduzidos
ente
as6h-1lh
e as 17h-19h,sob
condiçõesatnosféricas
favoúveis,
e
foram
Íealizadasduas
saídasde modo
a garantir a cobertura de todo o período de nidificação (e.g. Blondel eÍa/.
l98l).
Análise
de dadosOs
dadosforam
inseridosno Microsoft Office Excel
XP
em
duas mahizesdistintas: ambiental e biológica. Nesta
ultim4
utilizou-se o valor deindiüduos
registadomais
elevado,como
sendo,de
acordocom
Bibby et
al.
Q005),
o
númerode
aves presêntes. Ambas asmatizes
foram re-arranjadas (apêndicesI
e
II),
de modo que damatriz
ambiental foram criadas 3 grupos explicativos(esfirtura
fisica, habitat e pressãoantropogénica);
e
damatriz biológica
2
guildas (alimentaçãoe
estrato). Os grupos erefer€ncia. Os grupos ambientais foram adaptados com base numa revisão
bibliogáfica
adequada ao tema (e.g. Hughes et
a|.2002:
Scott eÍ a/.2003;Masot
et al. 2006; Sievinget
al.
1999) e por análises canónicas "tentativa" aos dados. Enquanto da construção dasguildas biológicas foram excluídas as especies para as quais o método de censo não
foi
considerado adequado
(excepto aves
planadoras
das
famílias: Hirundinidae
-andorinhas,
e
Meropidae
-
abelharucos),e
aves identificadas apenas atéao
género. Cada guilda discriminou para cada classe a percentagem de ocorrência dos indivíduos(no de especies por classe/no de indivíduos por classe) (apêndice
II).
No
tratamentode
dadosutilizou-se
o
programaPRMER 5
para as anílises:MDS
(Multiple
Discriminant Analysis), CLUSTER
e
SIMPER;
e
o
programaCANOCO
4.5 para Windows, na realização das análises canónicas-
CCA
(Canonical Correspondence Analysis) (Ter Braak 1986).A
MDS
foi
realizada paraclarificação
dos dadosobtidos em
cadatoço
e
aCLUSTER para facilitar
a
sua
interpretaçâo(em particular para níveis de
stess superioresa
0,1).
Para
constução das
matrizes
de
semelhanç4
üsto
as
escalasutilizadas serem diferentes, os dados foram estabilizados com uma raiz quadrada.
As
aruílises SIMPER foram efectuadas para melhor compreender as diferenças,identificando quais as espécies mais responsáveis pelas diferenças.
A
CCA
foi
realizadapara
exposigãoe
clarificação
des associaçôesentre
ascomunidades e as variáveis ambientais €sfuturadas em guildas (Jongman et
al.
1995;Titeux
e,
al.
2004;Knutson et
al.
2005). Para análise, as variáveisforam
reduzidasutilizando a opção "manual forward selection", com permutações Monte-Carlo (999), e
ins€ridas caso fossem consideradas
sigrificativas (p<0,1).Para
colocar os dados numa mesma escala de medida" estes foram estabilizados com uma raiz quadrada(kgendre
&
Legendre 1998; Knutson et a1.2005).RESTJLTAI'OS
Mrúriz
ambienúalEm termos de estrutuÍa
fisica,
e de acordo com os dados obtidos pelo RHS, astrês ribeiras diferenciam-se clammente, sendo que Ribeira de Arronches e a Ribeira de Seixe são mais semelhantes entre si do que com a Ribeira de Odelouca.
No
entanto, emr€speito aos aspectos
que
possam interessarpara
as comrmidades avifaunísticas, oslocais não
apresentam diferenciação geográLficasignificativ4
como
representado nafigura seguinte (resultados clarificados por uma anílise CLUSTER).
slúa:0.17 CDECl c{Ât
@3
ooEC,RR3 c:E-í2 oEpr /RRí @EC6 cÀÂl ,RR' (I)Elt
cÍE-l0ogl
CÍEL7 ,RftI (IE.0 Gcs oEg, ,RRs /RRí ffit (IEsFigure 2
-
fffico
de ordenação simples (não-constrangida) ilusbando os dois primeiros eixosda Multiple{imensional Scalling (MDS), efectuada aos vários locais de amostragem, contendo
apenas informação dos dados ambientais utilizados para criar os grupos de variáveis ambientais,
para um nível de stess de 0,17.
Os substratos predominantes nos canais são a
graúlha
e os seixos, excepto paÍaa
Ribeira de
Arronches onde predomina
a
areia
e
a
rocha nua.
O
fluxo de
águapredominante
na Ribeira de
Odeloucaé o
nâo
percepdvel (no perceptible
flon),
enquanto nas outras duas são o (zpweí&'zg) e com ondulação
(rippledJlow).
Na
Ribeira de Odelouca predominao
vale emforma
deV
eo
desfiladeiro.As
margers
são
predominantemenk
de
tena,
e
sâo
as
únicas
que
apÍ€sentam resseccionamento (na parte montante do Eoço).Do mesno
modo, é a única ribeira que apÍ€s€nta margens estáveis. Nas margens há predominância de arbustos, terras lavradâse pastagens abandonadas. EsG coberto arbustivo é importante para diversas aves, como
por
exemploo
rouxinol-comum (Luscinia
megarhynchos)e o
toutinegra-dos-valados(Sylvia
melanocephala),que
são espécies generalisas associadasa
coberto denso, eestão pÍ€setrtes
em
galerias ripícolas.
Como conseqúncia
destesusos
de solo,
a extensão da vegetação varia entre a classe uniforme e contínuo.Na
Ribeira
de
Seixe predomina
o
vale
assimétrico.
As
margens
sãoáars
(apêndiceI).
Apresenta usosde solo
variadoscom Fedominlincia
úsÍvel
de pastagens abandonadas, e a extensão da vegetação nan maÍgens é predominantemente semi-contínua.As ribeiras de Seixe e Odelouca apÍesentam consideravelmente mais locais com
floresta,
o
que se
reveste
de
uma
importância
acrescidapara
as
comunidades aüfaurústicas.A
forma
do
vale
é
bastantevariável na Ribeira de
Arronches,não
havendopredominância
clara.
As
rnargens são predominantemetrtede
terra,
e
estãona
suamaioria reforçadas com muros de pedra, os quais estão associadas
poÍ
nonna a hortas. Apresenta uma clara diversidade de usos de solo, e a extensão do coberto vegetativo ébastante variável nas margens, propiciando um aumento de biodiversidade.
Comunidadeg evifaunísúices
Para análise,
de modo
a
eütff
resútados
espúrios,foram omiüdas
todas asespécies que ocorÍeram em menos de
l0%
dos locais (e.g. Titerrx et a1.2004), de modoque as abundâncias e as riquezas específicas para cada ribeira (e para o
total
dos locais de amostragem), foram de acordo com o apresentado no quadro seguinte:QUADRO 2
-
Caracterização das Ribeiras de Seixe, Odelouca e Arronches e das três ribeiras em conjunto, em termos de comunidades avifaunÍsticaq através da apÍesentação do nimero totalde indivíduos, número
toal
de esfrcies, e índices de diversidade (Margalef, Simpson,Shannon-Wiener, Divorsidade máxima, e Pielou).
Métricr
Totrl
N! total de indiüduos No total de espécies
Ínaice ae
Uargatf
Índice de Simpson
Índice Shannon Wiener Diversidade máxima Ínüce de de Pielou 1581 33 4,50 0,05 3,12 3,53 0,88
Pela análise
do
quadro,verifica-se que
as comunidâdesem
cadaribeira
sãomüto
homogéneas (suportado pelos valorcsdo
índice de Pielou),
sendo as espécies quase identicamente abundantes,e
que
ente
si
também sãomuito
semelhantes emtermos
de
riqueza
específica.
Do
mesmo
modo
4presêntam
bons níveis
de biodivcrsidade (suportado pelos valores dos índices de Margalef e Simpson).Seire ()deloucr Arrunches
569 30 4,86 0,06 3,04 3,46 0,88 169 23 429 0,064
2,U
3,13 0,91u3
3l
4,45 0,056 3,00 3,43 0,88Após
agrupamentoem
guildas estrutuadasde
acordocom
o
estrato ocupadopelas aves
e
o tipo
de alimentação de cada espécie (apêndiceII),
verifica-sê, que ascomunidades das ribeiras de Seixe e Arronches são mais semelhantes entre si, e que as
da Ribeira de Odelouca formam um grupo mais isolado
(figuÍa
3).Súess:0,08 ODELT ODE-6 @ll2
*ffiff'
ory4PEc1 ODECADECí ÁRRt ÁRR6QEH-Iz
oer_cPEr8 ODECsoEg)Ec7
ODELS ODELíO ÁRRs CDECSFigura 3
-
Gnífico de ordenação simples (não-constrangida) ilustsando os dois primeiros eixosda Multiple{imansional Scalling (MDS), efeotuada aos vírios locais de amosüagem, contendo
apenas informação das guildas de espécies utilizados para as análises canónicas, para um nível
de stsess de 0,08.
Do
mesmo modo as comunidades nas ribeiras de Seixe e Arronches pouco sediferenciam uma da outra, à excepção do
local
Caia2m
Ribeira de Arronches, que éclaramente
o
mais
diferente
do
grupo,
deüdo ao facto de ser
o
que tem
maiorabundáncia de indivíduos.
Comunidades de aves
nrs
ribeiras
Ao realizar-se as
CCA's
para cada matriz explicativa verificou-se queI
pÍesença de margens resseccionadas,explicav4
a nível de modificações antopogénicas, mais de 50% da variância total nas comunidades. Deste modo, e porque esta característica afecta directamente a estruturaÍisica
daribeira
e do habitat, estavariável
foi
utilizada como co-variável nasCCA's
realizadas ás restanês matrizes, aumentando a consistência dos resultados.Desta perspectiv4 e de acordo com as anrílises canónicas, os factores que mais
inÍluenciam as comunidades são:
QUADRO 3
-
Factòs que de amrdo com ofiltro
aplicado (permutações Monte-CaÍlo parap{,1)
às análises canónicas(CCA)
às variáveis ambientaise
comunidades avifaunísticasestruturadas ern guildas, mais influenciaram a distribuição dos grupos de aves.
Alimentrçtro Estrato
Ertrutun F'ísicr
dr
Ribeire- Altura média da margem
- Largura do leito
- Sedimento d€ areia, silte ou argila - Forma do vale
- Profimdidade do leito
- LarguÍa do leito
- Sedimento de aÍeia" silte ou argila
- Forma do vale - Presença de r4fles Estruture Flsica do
Hrbitrt
- Vegetação ripícola - Rocha nua- CobeÍto vegetativo complexo na face da
maÍgem
- Ausência de cobeÍto vegetativo na face da margem
- Uso do solo (sweep-up) PomaÍ
- Uso do solo (sweepup) Arbustos e
Matos
- Uso do solo (sweep-up) Ervas altas - Uso do solo (sweepup) Tenas cultivadas
- Uso do solo na mugem Tenas cultivadas
- Margens estáveis - Rocha nua
-
CoHo
Yegetativo complexo na face da maryern- Coberto vegetativo simples na frce da maÍgem
- Ausência de coberto vegetativo na
frce da margem
- Uso do solo (sweepup) Eucaliptal
- Uso do solo na margem Pâstagem abandonada
É
de sublinhar a pÍ€ponderância que aestrutua
vegetativa na face da maÍgem temrus
comunidades, em especial a ausência de coberto vegetativo ou a sua pÍesença numa estrutura complexa (quadro 3).De acordo com as aruírlises às gpildas de alimentação, aves que se alimentam de insectos a€Í€amente são positivamente inÍluenciadas pela larguÍa do
leito
e pelos usosde solo:
ervas altâse
terra
lavradano raio
de
50m (sweep-up).As
aves piscícolasapaÍ€ntam
evitar locais
em que
o
leito
apres€ntaalguma
profrmdidade e./ou cujosedimento seja de areia, silte ou argila" e parecem ser influenciadas positivamente pela presênça
de
vegetaçãoriparia e
uso
de
solo pomar,
aÍbustose
matos tto
s$,ee?upE ^vEo?ix a A + a Yú ,l ET IL q ? ci -.0.8 0.8 -0.8 0.8
tr'igunc 4 e 5
-
Gráficos de ordenação simples ilustando os dois prirneiros eixos da CanonicalCorrespondence Analysis (CCA), efectuada com a percentagem de ocorÉnoia dos indivíduos por cada classe de alimentação, e os grupos de variáveis ambientais "estutura ÍIsica-' (figura a)
e "habitar" (figura 5). Os dados ambientais são represeutados por linhas (ver acrónimos no
apêndice I) e as variáveis relacionadas com as guildas de alimentação são representadas por:
E
-piscícola;0-insectívorasplanadoras;O-granívorast-insectívorasfolhagom;X-inseotívoras solo
A
- insectívoras fronco e casca das áryores.De
acordoçsm
as enálisss às guildas estrato, as avesaqúticall eütâÍn
locaiscujo
sedimento é de areia, silte ou argila, e paÍêcem beneficiar da ausência de coberto arbóreo e forma do vale. Aves de vegetação estâo relacionadas com uma maior largurade leito e com
cobertovegetativo
simplesna
margem.Aves de solo
ab€Íto
estão relacionadas com locais cujo sêdimento é de areia, silte ou argila, e em que a past4gem abandonadaé
o
usode
solo na margem. Tanto as aves de copa como as aéÍeas não parecem ser influenciadas pelas características ambientais obtidas. Os outros Íesultados-a
POx3 AC a v tfi ifl
o
x
+.!
o
q
o
ê
-0.6 1.0o
-0.6 0.8Figures 6 e 7
-
Gráficos de ordenação simples ilusrando os dois primeiros eixos da CanonicalCorrespondence Analysis (CCA), efectuada com a percentagem de ocorr€ncia dos indivíduos por cada classe de ocupação vertical do habitat
('estato')
e as variáveis ambientais "estrutura Ílsica" (figura 6) e "habitaf' (figura 7). Os dados ambientais são representados por linhas (veracónimos
no
aSndice
I)
e
as
variáveis relacionadascom as grildas de
estrato sãorepresentadas:
tr
- aqúticas O - vegeaçãof
- aereas X-
copaO
- solo aberto.Apesar das variâncias totais explicadas terem sido bostarte elevadas (quadro a)
para este
tipo
de análise (Heikkinen er aL 2004), as análises não reflectiram de maneirapÍecisa
as
associaçõesentre
espéciese
as
variáveis
ambientais
(figuras
4
e
5 especialmente).QUADRO 4
-
Resultados das Canonical Correspondence Analysis realizadas, apresentadas nasfiguras 4, 5, 6 e 7 respectivamente.
Totd
Vafiance
Erplained
Áxis
I
A:clsII
(o/clAlimentaçlo/ E truturr Fúicr da Ribeira Canonical Eigoralues Sp.
-
Ewironmental Correlaliow 0.068 0.963 43.0 0.025 0.805 59.0 86.1 §p. Yüiance accounted by nes (9'o)Alimêntaçlo/ Estrnturr tr'lsico do Hrbitât Canonical Eigewalues Sp.
-
bwironmental Correlalions Sp. Vqriance accounÍed by ues (/o)0.072 0.986 45.4 0.047 0.986 74.9 82.0 Estreto/ Estrutura Físicr da Ribeire Canonical Eigooalaes Sp. - E wbonmental CoüelaÍions Sp. Yariance accouúed by ues (/o)
0.089 0.884 39.1 0.033 0.753 53.9 92.3
Estrrtd
EstrutuÍr Ffulcr doHrbitst
Canonical Eigewalues Sp.-
b»ironmental ConelationsSp. Yriance accornted by
cet
(/o)0.092 0.894 40.6 0.051 0.922 63.1 79.3
DISCUSSÃO
Apesar
dasribeiras
de
Seixe
e
Odelouca abavessaÍemari
serras Algarvias,pertenceÍem ao barlavento algarvio e à mesma região
hidÍogÍáfica (3),
aaoilise MDS
aos dados Íecolhidospelo RHS Í€vela quc
a
Ribeira de
Seixeé mais
semelhante àRibeira
de
Arronches
do
que
à Ribeira de
Odelouca
Em
relaçãoàs
comunidadesaüfaunísticas
o
mesmo
se
repete,
e
neste caso
considera-sêque
o
facto de
os observadoÍes terem sido diferentes para a amosüagem das ribeiras afectou os Í€sútâdos.Mais
ainda"poÍ
haver claradiüsão
na análiseMDS
ás comunidades avifaunísticas no que Í€speita as3
ribeiras,
considera-seque
estefacto em conjunto com
a
diferentemetodologia de censo, teve um peso significativo na qualidade dos resütados, apesar de não haver possibilidade de quantificar/qualificar a extensão desse impacte.
A
diferença na metodologia de censo de aves apesar de serligeira,
inflú
nos resultados, urna vezque, oomo indicado
na
figua
2,
os
parâmetros ambientaistidos em
oonta para
aconstruçâo das manizes ambientais, não
reflectiram
a diferença geogÍáfica dos pontos de amostragem. Sendo esteultimo
factor, um ponto favorável à utilização do protocolo RHS paraa
construção de modelos de predição da distribuição das comunidades nasaplica
à
diferençaentre
o
númerode indiüduos
e
espécies registadospara
as üês ribeiras (quadro 2).Em
termosde RHS,
comoindicado em
Duarte(2007),
a
Ribeira de
Seixe éconsiderada
no
global
"mais
natural" que
a
Ribeira
de
Arronches.Neste
cont€xto, apenas a metade montante dotoço
da Ribeira de Odelouca analisado apresenta algumgrau de
modificação antropogénicae
menor qualidade.O
que, pelosmotivos
acima referidos, não teve impacte nas análises subsequentes. Contudo, em termos de habitats eusos de solo, a Ribeira de Odelouca apÍesenta claÍamente urna menor diversidade em
comparação
com
as
duas
outas
ribeiras, que
por
sua
vez se
assemelham emdivetsidade,
e
daí
também apresêntaÍmenor
númerode
especies (quadro2).
Nesta perspectiva e tendo em conta a opinião de autores comoMaÍtin
g
ps$ingham
(2005),podemos
aferir que
as
diferençasem
relaçãoao
número
de
espéciese
indivíduosregistadas nas ribeiras (quadrc 2), pode resultar do efeito que a
matriz
de usos de soloenvolvente
tem
nas comunidadesripícolas,
como consequência de uma adaptação asmudanças
de
habitat(Ries
&
Sisk 2004;
Martin
eral.
2006). Facto zuportado pelas anrílises canónicas subsequentes que identificam o maior peso do uso de solo no sweep-zp sobre adistribúção
das comunidades (quadro 3)No
quediz
respeito a números de espécies, as comunidades aüfaunísticas dasribeims
são moderadamente homogéneas(quadro
2),
sendoque
o
elevadovalor
deindiüduos
registados na Ribeira de Arronches se deve ao facto, de se teÍem encontadomais zonas urbanas
do
que nas duasouta
rib€iras, nas quais aves conspícuas comofringilídeos (e.g. Carduelis sp.),
andorinhasdos-beirais(Delichon
wbica)
e
ptdus
(Passer
domestic,us) abundam.Para diferenciaçtto das ribeiras,
e
clarificação
dos resultados, convém referir que na Ribeira de Odelouca" aocontário
da Ribeira de Seixee
Ribeira de Arronches não foram
registadas avescomo
andorinhas(Hirundo
sp.,Delichon urbica),
ptdus
(Passer domesticu,s) e abelharucos (Merops apiaster),o
qvetenâ
contibúdo
para a diferença no número de especies eindiüduos
registados. Estas diferenças são provavelmente a causa das diferenças marcadas na análiseMD§
(figura
3)
às comunidades, contudo,e
ap€saÍdo
métodode
censonão
ser,de
acordo com Thompson(2002)
o
mais adequado para estetipo
de aves, convémreferir
que estesforam
inclúdos
para análise urnavez
que,a
sua pÍesença nas guildas,implicou
um aumentona
robustezdos
resultadosnas
análises canónicas.Outro factor
que
terácensos na Ribeira de Odelouca. Considera-se que estes deviam ter sido realizados mais cedo.
Qualquer uma das ribeiras apresenta bons níveis de biodiversidade, sem haver clara distinção de uma gradação,
üsto
que a Ribeirade
Seixe apresenta os melhores valores para os índices de Margalef e Shannon-Wiener, a Ribeira de Odelouca apresentao
melhor
valor
parao
índice de Pielou,
e a
Ribeira de Anonches
parao
índice de Simpson (quadro 2).Em
relação à guildaestato,
há um aumento nãoJinear do número deesfrcies
por
classede
acordocom
urna gradação de vegetação:"solo
aberto", "vegetaçâo" e"copa", que por suavez é suportado por Scott et
al.
Q003).De acordo com as arálises canónicas às comunidades, a existência das margens resseccionadas
em
Odelouca explica maisde
50Yoü
variância em ambas as guildas (alimentaçãoe estato). Isto
prende-secom
o
facto da
ressecção das margens estar associada a drenagens e reperfilamento mecânico dorio,
o que resulta numa diminuiçãodo
número
depoint bars e riffles,
cobertura arbórea (semi-contínuae
contínua) eaumenta
o
substrato
de
silte
(Raven
er
al.
1998), factos
que
se
verificaramefectivamente, e que, de acordo com as anáises canónicas envolvendo a estrutura
fisica
das ribeiras, têm influência significativa nas comunidades. Contudo e por esta alteração
ter sido apenas registada em Odelouca, há que ter alguma cautela na lincarização deste
resútado, errbora as análises subsequentes imiscuam este factor com os oubos.
No
global
os resútados obtidos para cadaguilda foram
consistentes, tendo asCCA's
explicado uma boa pqcentagem da variação nas comunidades (quadro4). Do
mesmo modo
os
dados obtidos
em
separadopara
as
duas guildas
oferecem interpretações consistentes quando interpretados em conjunto, por exemplo: em relaçãoa
aves
piscícolas
e
aqúticas
(guildas
constituídas
sensivelmentepelas
mesmas especies), as aves piscícolas são influenciadas positivamente pela presença de vegetaçãoripícola
(indicativo
da melhor qualidade da galeria" e portanto maior disponibilidade dealimento) mas negativamente inÍluenciadas pela presença de substato de silte, areia ou argila" do mesmo modo que as
aqúticas.
Estas sÍlo ainda influenciadas positivamentepela presença de
rffies,
por
suavez
inversamente correlacionado coma
presença doanterior.
À
semelhança, aves classificadas como aéreas (andorinhas e abelharuco) quepor sua vez alimentam-se de insectos no ar, sâo positivamente influenciadas pela maior
largura
do leito
e
por um
coberto arbóreo complexo
na face da
margem.
Avessweep-up, .uana vez que nestas ráreas,
deüdo
à escassez de alimento, as aves retiram a maioriado
seualimento
das galeriasripícolas
(Gray
1993).Assim,
prova-se ainda,que
asribeiras
são importantesfonGs
secundáriasde alimento
para aves insectívoras, factotambém suportado
poÍ
autores como Mason etal.
(1984),ls;elta etal.
(2003)iz
Masonet aL (2006), e
Mwakami &
Nakano (2002).Aves de solo aberto são positivamente influenciadas pela presença de subshato de silte, areia ou argila pois este
implica
urna menor cobertura vegetativa na margem, epelo uso de solo pastagens abandonadas nas maÍgens, duas condições consistentemente
aliadas. Em oposição aves de copa são negativaÍnente influenciadas pela presença deste
substato
e
são
apaÍentemente influenciadaspela
presençade
Eucaliptal
nos
50m (sweepup). Isto resulta do aumento das explorações florestais desta árvore, e apesar daanálise
SIMPER identificar
o
Chapim-azú (Parus
caeruleus), como sendoa
especie responsávelpor
estadiferenç4
não existe na
bibliografia
dadosque
suportem estacorrelação,
à
par da espécie ser considerada generalista(Rufino
1989;Telleúa et
al. 1999).O
facto
das avesaqúticas
aparcntemente serem inÍluenciadaspela
forrna dovale
(figura 4),
prende-secom
o
facto
desteter
sido
contabilizadopor
nri,rneros de ordem (seguindo a ordem doformulário),
sendo que a Ribeira de Arronches e a Ribeira de Seixe apres€ntam os maiores valores de soma,e
são também as que apresêntam amaioria das aves aquáticas registadas. Este resultado é portanto espúrio.
Apesar dos resultados obtidos pelas análises canónicas terem sido consistentes
com a
bibliografia,
em termos gráficos não clarificaram adisnibúção
de algumas aves(figuras
a
a
7),
pelo que
haverá
ouhos
factores
mascarandoe
influenciando
os resultados que não foram incluídos nas análises. Como os dados analisados não foram inicialmente registados como
objectivo
de produção deste estudo, não se recolheram dados que de acordo com abibliografia
também poderiam causar diferenças espaciais, nomeadamente: especies de árvores dominantes na galeria ripícola (e.9.Stong
&
Bock
1990)
(factor
ausente nos dadosdo RHS
da Ribeira de Odelouca), densidadedo
sub-coberto arbustivo(Blb
et a1.2004), vale de cheia (e.g. Suong&
Bock
1990,HuMlek
1999), latitude, altitude,produtiüdade primária
e
complexidade geomorfológica (e.g.Buckton
&
Ormerod 2002). Estes factores, à excepção dos últimos citados em Buckton&
Ormerod
(2002),
são
contempladospelo formulário RHS,
contudo
não
foram utilizados neste estudo por: ou não terem sido inseridos discriminadamente nas bases de dados, ou não terem sido recolhidos no campo.Contudo as correlações obtidas pelas
CCA's
corroboram a hipótese de que asaves
utilizarn
aribeira
como conedor de dispersão (Mason eral.l984
&
lwalÀet
al.2003
iz
Mason eta|.2006,
Woinarski etaI.2000;
Groom&
Grubb 2002; Mason er aL 2005, Décamps etal.
1987; Roché 1989), urna vez que são os usos de solo na banda dos50m
no
sweep-up quena
suamaioria
são consideradossignificativos,
e
não
os
daÍrargem.
Para supoÍtar esta hipótese, foram também realizadas, à par destas anflises,aniílises canónicas enü€ os dados ambientais recolhidos eÍlr cÂÃà spot-chec& e os censos realizados nos mesmos, para as quais não se obtiveram resultados significativos. Nesta perspectiva também se reforça a premissa de que as aves que
utilizam
a galeria ripícola,estarão dependentes também de outros factores
tal
oomo setem
úndo 3
analissÍ ao longo deste trabalho. Contudo não se pode retirar uma conclusãodefinitiva
acerca desteponto, dado o peso que o coberto vegetativo na face das nrârgens tem sobre as guildas
(quadro
3), e
porque, precisamente, as comunidadesforam
eshuturadasem
guildas; separando,por
exemplo,
avescomo
o
guarda-rios(Alcedo
atthis), rouxinol-comr.rm(Lwcinia
megarhynchos)e
felosa-poliglota
(Hippolais polyglotta), que, caso
ascondições sejam
Fopícias
(existência
de
sub.coberto
arbustivo
súciente),
estão dependentes da galeriaripícola.
Todavia as análises às espécies em separado não sãoconclusivas, moüvo pelo qual não foram incluídas neste relatório. Nesta
ópticg
Shirley(2006), zugere uma interacçâo entre as comunidades de aves ripícolas e âs comuridades que
utilizam
a
matriz
envolvente,facto
que deverá serüdo
em
contapara
estudos futuros desta índole.Pela natureza da metodologia aplicad4 potencial dos resultados e dos objectivos propostos
pam
estetipo
de
trabalho, convém fazer
algumas sugestõese propôr
a modelaçâo das técnicas utilizadas. Para efeitos de constriçãodo tempo
dispendido a recolher dados no carnpo,visto
ser utilizado para monitorização dasliúas
defuua
noâmbito da
DQA,
o
RHS
é
uma tecnica perfeitamenteadeqúvel
quepode
fomecerresultados consistentes, caso haja uma adaptação das técnicas de inventariação de aves (Malllel et
aI.2000)
ao tipo de estudo pretendido. Do mesmo modo, e como sugeridopor
Buclrton
&
Ormerod (1997) eBrewin
eÍal.
(1998), é importante a inclusão de outrosdados nas análises, como
já
foi
preüamente referido
nesterelatório.
Em
relação à metodologiade
censode
aves,é
aconselluívela
suarealiz:ção ern
duasvoltas:
aprimeira em
Abril
e
a
segundaem Maio (com um
espaçamentonão inferior
a
4semanas), de modo a garantir a detecção das espécies residentes, que em geral
nidificam
mais cedo,e
das migradoras estivais,incluindo
as tardias(4).
O
método pontualpor
pontos de escuta aplicado segundo
Bibby
etal.
(1992),dwe
ser mantido estabelecendo duas bandas de censo: uma associada à galeriaripícola
para dados associados a spoÍ-checks,e
outÍa com
limitação
de 50m,
a
acompaúar
a
distância
estabelecida noprotocolo RHS para o sweep-up. Recomenda-se ainda, o registo nos pontos: 3, 6 e 9 do
RHS, de modo a cobrir a quase totalidade do troço RHS. Sublinha-se a necessidade de:
a) manter o mesmo observador para os pontos de escuta, para calibração dos dados, e, b)
regisar
todos osindiüduos
detectados auditivamente, paraoptimizaÍ
a
caÍacterizÂ+ão específica dos locais, como sugerido por Bryce etal.
Q002).A
estruturação rtes soÍnunidadss em guildas revelou-se extremamenteútil
paraanalisar
os
dados, mas estas, contudo devem ser estruturadascom
rigor, e
portantoconstuídas de acordo com a matriz biológica
obüda
Para a elaboração desterelaório,
esta
foi
estruturada de acordo com a gradação de vegetação ocupada pelas aves (estrato)e
alimentação, tendosido
tambemutilizada
para análise índices de biodiversidade eabundânci4 como sugerido por Hughes et
al.
Q002), e por Krater etal. Q00l).
Powell
&
Steidl (2000) sugerern ainda a análise do ponto deüsta
de caracterísúcas essenciais para a reprodução das aves ripícolas. Conhrdo, para esteultimo
factor não sê obtiveram resútados consistentes,motivo
pelo qual não foram aquie:çlicitados
esses resultados.Bryce et
al.
(2002), sublinham ainda que classificaçõesestitamente
funcionais (e.g.alimentação
e
nidificação)
por
vezes podemnão
produzir
sinaisclaros,
quando os habitâts são muito variáveis,motivo
que também podeÍá estâr por detnás dos resultadospouco precisos
(figuras
4 a 7)
obtidos, e reforçandoa ideia
de quea elahração
dasguildas deve ser ponderada.
A
criação de uma guilda contendo apenas aves ripícolas,poderá também ser
útil
paraclarificar
a associação dcstas à galeria ripícola, everificaÍ
seh4
e nesse caso qual,o
impacte que a esüutura destâtem
sob,re a comunidade de aves. Da qualidade destes resultados pode advir a possibiüdade de desenvolümento de modelos de predição do efeito de impactes ambientais sobre as comunidades, ou ainda o desenvolvimento de índices de qualidade (e.g. Oliveira&
Cortes 2006).Por
ú[timo
deixa-seem
abertoa
hipótese deutilizar a
extensão da vegetaçiioripária como co-variável nas análises, paÍa testaÍ a dependência das aves em relação a esta.
Este
parâmetronão
foi
analisado nesterelatorio
üsto
a
basede
dadosnilo
discriminar esta extensllo de modo aproveitrível para as aruálises.coNSIDERAÇôBs
uxlrs
n
coNcLUsôEs
Em conclusÍlo, apesar dos dados não terem sido recolhidos com esta orientaçâo
foi
possível estudaras
respostasdas
comunidadesde
avesem
relaçãoàs
galeriasripícolas mediterrânicas, através
de
dados recolhidospelo protocolo RHS e
metodospontuais
de
censode
aves, demonstrandoo
potencial
do
uso
destas técnicas emconjunto.
Do
mesmomodo,
demonstra-sea
vantâgemda
estruturação dos dados em gurldas, que poderão ser utilizadas para:a)
estudar as comunidades de aves,b)
criarmodelos de predição do efeito de impactes ambientais sobre as comunidades ou índices de qualidade.
Gostaria
apenas
de
agradecer
à
Prof. Maria
Teresa Ferreira
pela
suadisponibilidade, profissionalismo e excelente orientaçiio; e à doutouranda Anâ Mendes
pela
cedênciados
seus dados acercada Ribeira de
Odeloucae
Íesposta as minhas questões.RET'ERÊNCIAS
BIBLIOGRÁX'ICAS
Aguiar,
F. C.
&
Feneira"
M. T.
(2005).
Human-disturbed landscapes:effects
oncomposition and
integrity
of
riparian
woody
vegetationin
the
TagusRiver
basirq Por1atgal.Ewirownental
Consemation 32(l):
3041
.Alves,
M.
H.,
Bemardo, J.M.,
Cortes,R.
V.,
Feio,M.
J., FerreirA J., Ferreira,M.
T., Figueiredo,H.,
Fomrigo,N.,
Ilhéu,
M.,
Morais,
M.,
Pinto,
P.&
Rafael,M. T.
2006.Tipologia
dc rios em Portugal Continental no âmbito daDirectiva
Quadro daÁgua
8" Congresso da Água.Bibby, C., Burguess,
N.
D.& Hill,
D.A.
(1992)Bird
Census TechniErcs, Acadernic Press,Bibby,
C.J., Burgess,N.D., Hill, D.A.
&
Mustoe, S.H. (2005)Bird
census techniques.2nd ed. Elsevier Academic Press,
London
Blondel,
J.,
Ferry,
C.
&
Frochot,
B.
l98l
-
Point
countswiú
unlimited
distance. Studies in AvianBiologt, 6:
414-420.Brewfuu
P.A., Buckton, S.
T.
&
Ormerod,
S.
J.
1998.
River
habitat surveys
andbiodiversity
in
acid-sensitiveivers. Aquatic
Conserlratior.
Marine
and
FreslmaterEcosyrtems E: 501-514
Bryce, S.
A.,
HughesR.
M.
&
Kauftrann,
P.R.
2002. Developmentof
abird intesnty
index: Using
bird
assemblagesas
indicators
of
riparian condition.
EnvironmentalManagement 30 :294-310.
Bub,
8.R.,
Flaspolúer, D.J.&
Huckins, D.J. (2004) Riparian and upland breeding-birdassemblages
along
headwater streamsin
Michigan's Upper
Peninsula-Journal
of
Wildl ife Manage ment, 6E Qr, 3 83 -392.
Buckton,
S.T.
&
Ormerod, S.J.
1997. Useof
anew
standardizedhúitat
surveyfor
assessing the habitat preferences and
distribution
of
uplandriver
birds.Bird
Study 44:127-337.
Buckton, S. T.
&
Ormerod, S. J. 2002. Global pattemsof
diversity among the specialist birds of riverine landscapes.Freslwater
Biologt
47: 695-709.Buffagni,
A.
&
Kemp,
l.
2002.l,ooking
beyondthe
shoresof
theUniked
Kingdom:addenda
for
the
application
of
River Habitat
Survey
in
Southern European rivers.Jownal of
Limnologt.612:
199-214.Covas, R.
&
Blondel, J. (1998) Biogeography and historyof
Mediterraneanbird
fauna. Ibis, 140:395-407.Davenport,
A.
J.,
Gumell,
A.
M.
&
Armitage,
P.D.2001. Classi$ing
urban rivers.ir:
..Décamps,
H.,
Joachim, J.&
LaWa"J.
1987.The
importancefor
birdsof
the riparian woodlandsúthin
the
alluüal
corridor
of
the River
Garonne,SW
France. RegulatedRivers, 1,301-316.
Décamps
H,
FoÍtuneM,
Gazelle F, Patou G (1988)Historical
influenceof
man on theriparian dynamics of a
fluüal
landscape. LandscapeEcologt,
l,
163-173.Doherty, P. F., and T. C. Grubb. 2002. Survivorship ofpennanent-resident birds in a
frap.ented
forested landscape.Ecologt
83:8M-857 .Drurte,
L.
2007.Bird
communitiesin
riparian
areas andRiver Habitat
Surveyin
the South of Portugal. Trabalho Apresentado para a Conclusão da Licenciatura emBiologia
-
Variante de Biologia. Universidade deÉvorg
Évora. Portugal.Enúronment Agency
(1997)Introductory
Gaidanceon
the Agency'sContribution
to Suslainable Development, Bristol,UK,
Environment Agency.Enüronment
Agency,2003.
River Habitat&ntey
inBritain
andlreland.
SEPA.Forman,
RT.T.
(1997) Land
mosaics:
the
ecology
of
landscapesand
regions. Conservation B iologt,
E, 50-59.Ferreir4 M.
T.&
Aguiar, F. C. 2006. Riparian and aquatic vege{ntionin
Mediterranean-type shearrs (Westem Ibéria).Iiznetica,2í(l-2)i
411424
Fox, P. J.
A.,
Naura,M.
&
Scarlett, P. 1998.An
accountof
the derivationof
a standardfield
method,River Habitat Swrey. Aquatic
Consemation:Marine and
FreshwaterEcosystems
8:
455-475.Fumess,
RW.,
Greenwood J.J.D.&
Jarvis P.J. 1993. Can birds be usedto
monitorúe
enúronment?,p. l-41.
In:, R.W. Fumess&
J.J.D Greenwood @ds). Birds as monitorsof
enúronmental change. London, Chapman&
Hall,
325p.Gray,
L. J.
1993. Responseof
InsectivorousBirds
to
Emerging
Aquatic
Insectsin
Riparian Habitatsof
a Tallgrass Prairie Steam. ÁmericanMidland Naturalist
129:288-300.
Groom, J.D.
&
Grubb Jr., T.C. (2002)Bird
species associatedwith
riparian woodlandin
frap.ented,
temperaE-deciduous forest. ConservattonBiologt,ló(3):
832-836.Heikkinen,
R
K.,
Luoto,M.,
Virkkala
R
&
Rainio,K.
(2004) Effectsof
habitat cover, landscapestructue
and spatial variables on the abundanceof
birds
in
anagricultural-forest mosaic.
"/ownal of
AppliedEcologt,4l,824-835.
Hubaleh
Z.
1999. Seasonal changes of bird communities in a managed lowland riverineecosystem.
Folia
Zoologica 48:203-210.Hughes,
S.
J.,
Ferreira,T.
&
Cortes,R.
V.
2007. Hierarchical
spatial pattems anddrivers
of
chânge
in
benúic
macroinvertebraê communities
in
an
intermittentMediterranean river. Aquatic Consemation: Marine and
Freslu'dter
Ecosystems866i
l-19.
JanseÍL
A.
&
Robertson,A.
I.
(2001). Riparianbird
communitiesin
Íelation
to
landmanagement practices
in
floodplain
woodlandsof
south-eastemAustralia. Biological
Conservation, 100(2): I 73-l 85.
Jongmarl R.H.G., C.J.F.
Ter
Braak,&
O.F.R.Van
Tongeren. 1995. DataAnalysis
in
Community and Landscape Ecology. Cambridge Universityhess,
Cambridge,U.K.
Knutson,
M.
G., and E. E. Klaas. 1997. Declinesin
abundance and species richnessof
birdsfollowing
a major flood on the UpperMississippiRiver. Auk
114:,367 -380.Kratt€r,
A. W.,
Steadman,D.
W.,
Smittr,C. E., Filardi, C. E.
&
Webb,H.
P. (2001).Knutson,
M.
G., McColl,
L.
E.
&
Suarez,§.
A.
2005.
BreedingBird
AssemblagesAssociated
With
Stagesof
Forest
Successionin
Large
River
Floodplains.
Natwal
Areas
Journal25:55-70
Legendre,
P.,
&
L.
Legendre. 1998.Nunerical Ecologt
(2ndedition).
Elsevier,New
York.
Iowrance,
R.
&
Crow,
S.R. (2002)
Implementationof
riparian
buffer
systemsfor
landscape management.
Ecologt
landscapein
AgroecosystemsMatagemenÍ (ed. L.
Ryszkowski), pp . 145-157 . CRC Press, Boca Raton,Florida USA
Iúanel,
S.D.,
Buckton, S.
&
Ormerod,S.
1999.Altemative
methodsfor
predictingspecies
distribution:
an
illustration
with
Himalayan
river
bnds.
Journal
of
Applied
Ecologt
S6:734-747.Manel, S., Buckton, S.
T.
&
Ormerod, S.I.
2000. Testing large-scale hypoüeses using surveys: the effectsof
land useon
the habitats, invertebrates andbirds
of
Himalayanlivers. Journal
ofApplied Ecologt
ST: 756.MartiÍl
T. G., Mclntyre,
S.,
Cathrall, C.
P., Possingharl
H.
P. 2006.
Is
LandscapeContext
Important
for
Riparian
Conservation?Birds
in
Grassy Woodland.Biolo§cal
C ons emation 127
Q):
201 -21 4MaÍtin,
T. G.&
Possingham, H. P. (2005). Predicting the impactof
livestock grazing onbirds using foraging height data Journal
ofApplied
Ecolog
42(2):400408.
Masoq C.F.,
Macdonald,
S.M.
&
Hussey,A.
(1984)
Structure, management and consenrationvalue
of
the riparian woody plant
community.Biolo§cal
Consemation,29,201-206.
Mason, C. F.,
Hoftranru
T.A.
&
Macdonald, S.M.
2006. The winter bird communityof
river
corridorsin
eastem Englandin
relation to habitat variables. Ornis FennicaMorais,
M.,
Pinto, P., Gúlherme,
P.
Rosado,J.
&
Antunes,l.
2004. AssessÍnentof
temporary streaÍns:
the
robustnessof
metric
andmultimetric
índices underdifferent
hydrological conditions. Hyfuobiologia
516: 229 -249.Murakami,
M.,
and S. Nakano. 2002.Indirect
effectof
aquatic insect emergence on a terrestrial insect population through predation by birds.Ecologt
Letters 5:333-337.Newsoq
S. E., Noble,D.
G.,&
Marchant
J. 2005. Waterways BreedingBird
Survey:preliminary analyses
of
mammal datâ 1998-2003. BTO Research ReporÍ No. 405.Newton,
l.
(1998) Populationlimitation
inbrró.
Academic PressLimited,
London.Oliveira, S.
V.
&
Cortes,R.
M. V.
2006. Enüronmental indicatoÍs
of
ecologicalintegnty
andtheir
developmentfor
running waters
in
northem
Portugal. Limnetica,25(l-2):479-498.
Pais,
M.
C.
&
Cangarato,R.
[2001].
Roteiroscom
As
avesdo Alentejo.
Cento
de Estudos deAüfauna
lbérica. Évora" Portugal.Powell,
B.
F.
&
Steidl,
R.
J.
(2000). Nesting habitat and reproductive
successof
souúwestem riparian birds. The Condor 102(4):823-831
Raven, P.
J., Fox, P.,
Everar{ M., Holnes, N. T. H.
&
Dawson,F.
H.
1997.River
habitat structure: a new systÊÍnfor
classiffing rivers
accordingto
their habitât quality. FeshwaterQtality:
DeÍin@
theIndefinable?
(eds P.J. Boon
&
D.
L.
Howell).
TheStationary
Oflice,
Edinburg[
241-250 pp.Raven, P. J., Holmes,
N.
T. H., Dawson, F. H., Fox, P. J.A.,
Everard,M.,
Fozzard,I.
R
&
Roue&
K.
J.
1998a.River
Habitat Quality. The physical
characterof
rivers
and sheams in theUK
and Isle of Man.Enirorunental
Agency, River Habitat &trvey Report No.2.Raven,
P.
J.,
Holmes,
N. T.
H.,
Dawsorl
F. H.
&
EvenÍ4
M.
1998b. Quality
assessment
using River Habitât Survey
data.Áquatic
Consemation:
Marine
andRaven, P. J., Holmes,
N. T. H.,
Charrier, P., Dawson,F.
H., Naura,M.
&
Boon, P. J.2002. Towards a harmonized approach
for
hydromorphological assessmentof
riversin
Europe:a
Qtralitative
comparisonof
three
surveymethods. Aquatic
Conservation:Marine and Freslmtüer Ecosystems 12: 405-424.
Ries,
L.
&
SislqT.D.,
2004.A
pÍedictive model
of
edge effects.Ecologt
85,2917-2926.Roché,
J.
1989.Un
gradient ecologique: la succession des oiseaux nicheurs le long descours d'eau.
lcÍa
Oecolo§ca 10: 37-50.Rufino, R.
(Coord.)
(1989)Atlas
das
ovesque nidifrcam
emPortugal
Continental.CEMPA
Serviço Nacional de Parques, Reservas e Conservação da Natureza.Lisboa
Saraiva,
M.G.M.A.N.L.
(1999) Orio
comopaisagen.
Fundação Calouste Gulbenkian, Fundação para a Ciência e Tecnologia.Lisboa-Scott,
M.L.,
Skagen, S.K.&
Merigliano, M.F.
(2003)Relating
change and grazing toaüan
communities in riparian forests . CorcemationBiologt,
l7(l):284-296.
Shirley, S.
M.
(2006). Movementof
forest
birds
acrossriver
and clearcut edgesof
varying riparian buffer strip
údths
. Forest ecologt and management223(l-3),
190-199Sieüng,
I(
E.,Willson, M.
F.&
De Santo, T.L.
(2000). Definingconidor
frrnctionsfor
enderricbirds
in
fraggrent€d south-temperate rainforest. ConsemationBiologt l4(4):
tt20-1132.
Smith, C.M.
&
Wachob, D.G. (2006) Trends associatedwith
residential developmentin
riparian breeding
bird
habitat
along the
Snakeriver
in
JacksonHole,
WY,
USA: implications for conservation planning.Biolo§cal
Conservation,12E,431446.
Strong,