Modulation of cytokine release and gene expression by the immunosuppressive domain of gp41 of HIV-1.
Texto
Imagem
Documentos relacionados
The two points considered at the alternate sides, of the tangents through the diameter of the circle, and then the line joining these points divides the circle
In this work, we report the expression of the N-terminal domain of TPL in Pichia pastoris to study the effect of the C-terminal domain deletion on the enzyme activity, and to verify
social assistance. The protection of jobs within some enterprises, cooperatives, forms of economical associations, constitute an efficient social policy, totally different from
Tidal analysis of 29-days time series of elevations and currents for each grid point generated corange and cophase lines as well as the correspondent axes of the current ellipses
In this paper, we present a version of the Invariance of Domain Theorem for nonlinear Fredholm maps of index zero between Banach spaces (or, more generally, Banach manifolds).. To
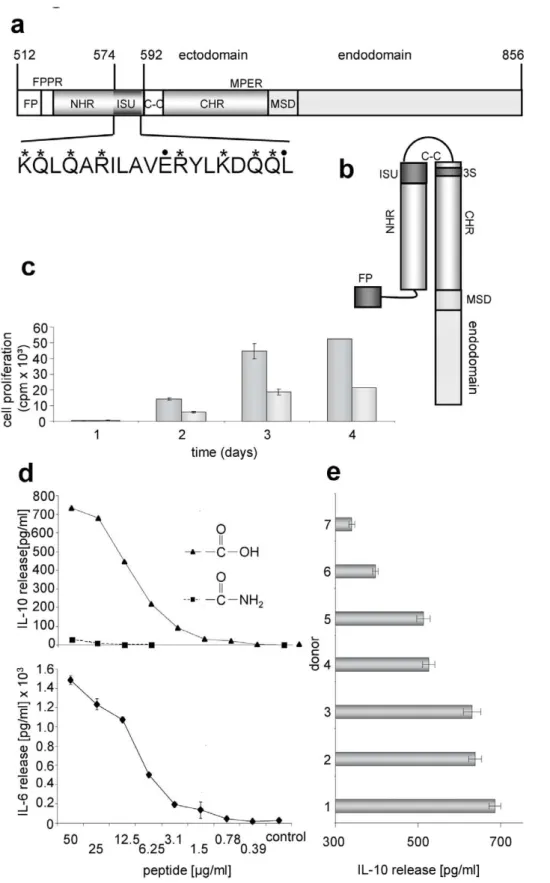
28,29 described a rapid and simple method for the identification of HIV-1 subtypes by PCR amplification of a region of gp41 using subtype-specific primers.. Therefore, our aim was
In the present study, we predicted ex- posed domains of gp120 and gp41 of Indian isolates of HIV-1 and then we used synthetic peptides mainly corresponding to the C2, V3 and CD4
Protein sequence analysis confirmed that SmPlk1 shared the common structure of Polo members com- posed of a highly conserved N-terminal kinase domain and a C-terminal polo-box
