cells harbouring different genetic background:
the role of the lysosome.
Dissertação para obtenção de grau de Mestre
Mestrado em Biotecnologia para as Ciências da SaúdeCAROLINA ALMEIDA MARQUES
UNIVERSIDADE DE TRÁS-OS-MONTES E ALTO DOURO
VILA REAL, 2010
ii Este trabalho foi realizado no Departamento de Biologia da Universidade do Minho, sob a supervisão da Professora Doutora Ana Preto e da Professora Doutora Olga Coutinho. A sua execução foi financiada pelo projecto PTDC/BIA-BCM/69448/2006, atribuído pela FCT.
iii
A
CKNOWLEDGEMENTSChegada ao fim mais uma importante etapa da minha vida, feita de alguns contratempos e outras tantas vitórias, há algumas pessoas que merecem o meu especial agradecimento por terem feito parte desta etapa e terem, de forma directa ou indirecta, contribuído para que mais este objectivo fosse atingido:
À Professora Doutora Ana Preto, em primeiro lugar por ter aceite ser minha orientadora. Por ter confiado em mim e me ter apoiado incondicionalmente. Por todos os ensinamentos que me transmitiu e por me ter feito crescer cientificamente, mas acima de tudo, a nível pessoal. O meu muito obrigada!
À Professora Doutora Olga Coutinho, por ter aceite ser minha co-orientadora. Obrigada por toda a disponibilidade e pelos ensinamentos transmitidos ao longo da realização deste trabalho.
À Professora Doutora Isabel Gaivão, por ter aceite ser a minha coordenadora na UTAD, por toda a disponibilidade que sempre demonstrou e por mais uma vez me acompanhar numa etapa tão importante.
Aos meus Colegas de Laboratório, por me terem recebido e por se terem mostrado sempre disponíveis para ajudar em tudo. Ao João Pedro, por me ter ajudado à integração no trabalho de laboratório. Ao Cristóvão, pelos ensinamentos e pelas dicas brilhantes que deu para a realização deste trabalho. Às meninas, em especial à Vera, à Cristina Carvalho, à Ana Oliveira, à Hanna e à Flávia, e ao Filipe, pelos bons momentos de descontracção, pela partilha de angústias e pela ajuda sempre prestada quando necessário. À Sofia, companheira nesta viagem desde Vila Real até Braga, por tudo! Obrigada a todos!
À Sara, pela preciosa ajuda nestes últimos tempos, quando tudo parecia correr mal e finalmente correu bem, graças a ti, à tua disponibilidade e incansável dedicação! Um obrigada muito especial!
Aos colegas de mestrado, ou melhor, aos amigos de Vila Real. Porque só vale a pena festejar esta vitória se o fizermos todos juntos, como o fizemos desde 2005,
iv quando éramos ―pequeninos‖ e tudo parecia mais fácil. Obrigada pelo companheirismo, mas acima de tudo pela amizade!
Ao Luís, por tudo aquilo que és para mim, por tudo o que fazes por mim e por me deixares sonhar contigo! Obrigada por me ouvires, por me aturares quando ―a vida se alagava‖, por muito me teres ajudado, de todas as formas que te foram possíveis, para que este meu objectivo fosse finalmente cumprido.
À minha Família. À minha Mãe, por me mostrar o seu amor condicional e a sua preocupação constante comigo. Ao meu Pai, por tudo aquilo que luta para me dar sempre o melhor e por ser o meu exemplo e orgulho. À Ju, pela companhia, amizade e apoio que sempre demonstrou. A todos pelas palavras de incentivo durante esta caminhada e por saber que posso sempre contar convosco!
v
A
BSTRACTColorectal carcinoma (CRC) is the third most common form of cancer in the world, with more than one million estimated new cases per year, from which roughly half resulted in death, being one of the most common causes of cancer death worldwide. In this context the finding of new therapeutic approaches is of prime importance for these types of carcinomas. Two major oncogenes are implicated in sporadic colorectal carcinogenesis, KRAS and BRAF mutations which are alternative and occur respectively, in 30% and 10% of CRC cases.
The use of dietary propionibacteria as a probiotic might be a useful strategy in CRC therapy. This bacteria produces short-chain fatty acids in the human intestine, namely propionate and acetate. Acetate can induce apoptotic cell death in colorectal carcinoma cells and not in normal colorectal cells.
Lysosomes have been implicated in cell death pathways, since selective lysosomal membrane permeabilization (LMP) and release of lysosomal enzymes, such as cathepsins to the cytosol may induce apoptosis. It has been proved that in acetic acid-treated S. cerevisiae cell death is induced through mitochondrial degradation and Pep4p, the yeast ortholog of human cathepsin D, is released from the vacuole into the cytosol.
Ceramide and related sphingolipids play an evolutionarily conserved role in the cellular response to stress by regulating cell growth, differentiation, senescence, and survival. Ceramide plays a particularly role in cancer, being able to induce apoptosis in tumour cells, including human colorectal carcinoma cells. Ceramide and its metabolism may have an important role in LMP induction. It has been shown that ceramide generated by the endolysosomal acid sphingomyelinases binds and activates the lysosomal Cat D.
The involvement of ceramide and LMP in the cell death induced by acetate in CRC cells harbouring different oncogene activation to the best of our knowledge was never investigated.
In the present study, we aimed to study the different sensitivity to acetate in CRC cells harbouring mutations in KRAS and BRAF oncogenes and to elucidate the involvement of LMP and ceramide in acetate-induced cell death in these cells. For that purpose we used two CRC derived cell lines: HCT-15 cell line, with a KRASG13D mutation and RKO cell line with a BRAFV600E mutation. The cell lines were treated with
vi sodium acetate at different concentrations and we performed studies on cell viability, cell proliferation and apoptosis. We also performed analysis of LMP induction and analysis of the role of ceramide.
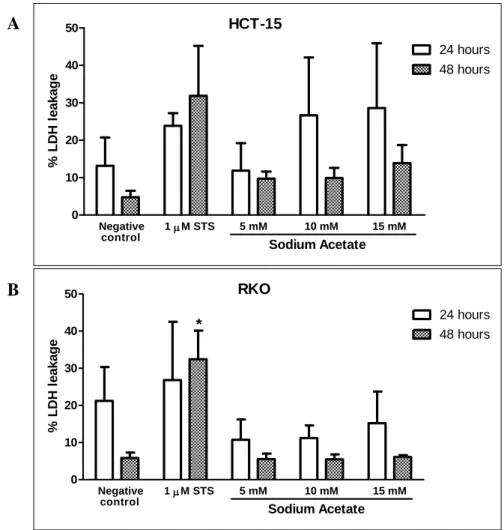
Our results confirmed that acetate induces apoptosis and showed that HCT-15 cells are more sensitive to acetate since the IC50 found for RKO cell line is almost double of
the IC50 found for HCT-15 cell line, indicating that the genetic background might have
an important role on this response. In both cell lines, acetate inhibited cell proliferation and induced apoptosis. Our results also show that cathepsin D seems to be released to the cytosol in response to acetate, in contrast with untreated cells, suggesting the involvement of LMP in acetate-induced apoptosis in CRC cells. Our results in the role of ceramide in acetate-induced apoptosis through LMP need further investigation.
In summary, our results showed that CRC cells harbouring KRASG13D mutation are more sensitive to acetate than those harbouring BRAFV600E mutation and that LMP seems to be a mechanism of acetate-induced apoptosis in CRC. Tumour cells seem to be more sensitive to lysosome-destabilizing agents, prompting the possibility of exploiting this pathway to selectively induce apoptosis in CRC cells. Understanding the mechanisms underlying acetate-induced cell death, namely the involvement of ceramide and LMP, provide the possibility of using acetate and other LMP inducers, as therapeutic agents in CRC therapy, alone or in association with conventional therapy.
vii
R
ESUMOO carcinoma colorectal é o terceiro tipo de cancro mais comum no mundo, estimando-se que existam mais de um milhão de novos casos por ano e aproximadamente metade dos casos resultam em morte, sendo uma das causas mais comuns de morte por cancro no mundo. Neste contexto, torna-se importante a descoberta de novas abordagens terapêuticas para este tipo de carcinomas. Existem dois oncogenes principais implicados no desenvolvimento do cancro colorectal esporádico, sendo as mutações KRAS e BRAF mutuamente exclusivas, ocorrendo em 30% e 10% dos casos de carcinoma colorectal, respectivamente.
O uso da propionibacteria, proveniente da dieta alimentar, como probiótico pode vir a tornar-se uma estratégia útil no combate ao carcinoma colorectal. Estas bactérias produzem ácidos gordos de cadeia curta no intestino, nomeadamente propionato e acetato. Este último parece induzir a morte por apoptose em células do carcinoma colorectal mas não em células normais.
Os lisossomas estão implicados em vias da morte celular, uma vez que a permeabilização da membrana lisossomal e a libertação de enzimas lisossomais, como as catepsinas, para o citoplasma pode induzir apoptose. O tratamento com ácido acético em S. cerevisiae induz morte celular através da degradação da mitocôndria e o Pep4p, o ortólogo da catepsina D humana nas leveduras, é libertado do vacúolo para o citoplasma.
A ceramida e os esfingolípidos relacionados têm um papel conservado evolutivamente na resposta celular ao stress, regulando o crescimento celular, a diferenciação, senescência e sobrevivência. A ceramida desempenha um papel importante no cancro, sendo capaz de induzir apoptose em células tumorais, incluindo células do carcinoma colorectal. A ceramida e o seu metabolismo podem também ter um papel importante na indução da permeabilização da membrana do lisossoma, já que a ceramida gerada pela esfingomielinase ácida endolisosomal liga-se e activa a catepsina D.
O envolvimento da ceramida e da permeabilização da membrana do lisossoma na morte celular induzida pelo acetato em células do carcinoma colorectal com diferente activação oncogénica, tanto quanto temos conhecimento, nunca foi investigada.
viii No presente estudo, o nosso objectivo era o de estudar a diferente sensibilidade ao acetato em células do carcinoma colorectal com mutações nos ongenes KRAS e BRAF e elucidar o envolvimento da permeabilização da membrana do lisossoma e da ceramida na morte celular induzida pelo acetato nestas células. Para esse fim, usámos duas linhas celulares derivadas de carcinoma colorectal: HCT-15, com a mutação KRASG13D, e RKO com a mutação a BRAFV600E. As linhas celulares foram tratadas com diferentes concentrações de acetato de sódio e foram realizados estudos sobre a viabilidade celular, proliferação celular e apoptose. Também foi analisada a indução da permeabilização da membrana do lisossoma e o papel da ceramida.
Os nossos resultados confirmam que o acetato induz apoptose e demonstram que as células da linha celular HCT-15 são mais sensíveis ao acetato uma vez que o CI50
determinado para a linha celular RKO é quase o dobro do encontrado para a linha HCT-15, indicando que o background genético pode desempenhar um papel importante nesta resposta. Em ambas as linhas celulares, o acetato inibiu a proliferação celular e induziu a apoptose. Os nossos resultados demonstram ainda que a catepsina D parece ser libertada para o citoplasma em resposta ao acetato, ao contrário do que acontece às células não tratadas (controlo), sugerindo o envolvimento da permeabilização da membrana do lisossoma na apoptose induzida pelo acetato em células do carcinoma colorectal. Os nossos resultados sobre o papel da ceramida na apoptose induzida pelo acetato através da permeabilização da membrana do lisossoma carecem de mais investigação.
Em resumo, os nossos resultados mostram que as células com a mutação KRASG13D são mais sensíveis ao acetato do que aquelas com a mutação BRAFV600E e que a permeabilização da membrana do lisossoma parece ser o mecanismo envolvido na apoptose induzida pelo acetato no carcinoma colorectal. As células tumorais parecem ser mais sensíveis a agentes destabilizadores do lisossoma, proporcionado a possibilidade de explorar esta via para induzir selectivamente a apoptose em células do carcinoma colorectal. A compreensão dos mecanismos subjacentes à morte celular induzida pelo acetato, nomeadamente o envolvimento da ceramida e da permeabilização da membrana do lisossoma oferece a possibilidade de usar o acetato e outros indutores da permeabilização da membrana do lisossoma como agentes terapêuticos no tratamento do carcinoma colorectal, por si só ou associados com outros agentes convencionais.
ix
T
ABLE OFC
ONTENTS ACKNOWLEDGEMENTS ... iii ABSTRACT ... v RESUMO ... vii TABLE OF CONTENTS ... ix ABBREVIATIONS ... xii 1 INTRODUCTION ... 1 1.1Hallmarks of cancer ... 2 1.2Colorectal carcinoma ... 31.2.1 Colorectal Carcinoma risk factors and prevention ... 4
1.2.2 Oncogenic activation in Colorectal Carcinoma: KRAS and BRAF Mutations ... 5
1.3Apoptosis ... 6
1.3.1 The role of apoptosis in normal colorectum and colorectal carcinogenesis .. 7
1.3.2 Acetic acid/acetate-induced cell death in S.cerevisiae and in colorectal carcinomas ... 9
1.4Lysosome membrane permeabilization as a lysosomal pathway of cell death... 11
1.4.1 Lysosomal enzymes ... 12
1.4.2 Mechanisms of lysosome mediated apoptosis ... 14
1.4.3 Methods for detection of lysosome membrane permeabilization ... 16
1.4.4 Lysosome membrane permeabilization and cancer ... 18
1.4.5 Lysosome membrane permeabilization and cathepsin D expression in colorectal carcinomas ... 20
1.5 Ceramide ... 21
1.5.1 Ceramide and apoptosis ... 22
x
1.5.3 The role of ceramide in colorectal carcinomas ... 25
1.6 Rationale and aim ... 27
2 MATERIALS AND METHODS ... 29
2.1 Cell lines and culture conditions ... 30
2.2 LDH leakage assay ... 30
2.3 MTT reduction assay ... 31
2.4 TUNEL assay ... 33
2.5 SRB assay ... 34
2.6 Lysosome membrane permeabilization (LMP) detection ... 36
2.6.1 Acridine Orange Staining ... 36
2.6.2 Immunoblot detection of Cathepsin D... 37
2.6.2.1Preparation of cytosolic fraction ... 37
2.6.2.2Preparation of total protein extracts ... 38
2.6.2.3Protein quantification ... 38
2.6.2.4 Protein precipitation ... 39
2.6.2.5 Western Blotting ... 39
2.6.3 Immunofluorescence detection of Cathepsin D ... 40
2.7 Statistical Analysis ... 41
3 RESULTS AND DISCUSSION ... 42
3.1 Effect of sodium acetate on cell viability of CRC cell lines ... 43
3.2 Effect of sodium acetate on extracellular pH ... 47
3.3 Effect of sodium acetate on CRC cell lines proliferation ... 48
3.4 Effect of sodium acetate on CRC cell lines apoptosis ... 50
3.5 Lysosome Membrane Permeabilization Detection ... 54
3.5.1 Acridine Orange staining and Immunofluorescence detection of Cat D ... 54
3.5.2 Immunoblot detection of Cathepsin D in subcellular fractions ... 57
xi 4 CONCLUSIONS AND FUTURE PERSPECTIVES ... 61
xii
A
BBREVIATIONSalkSMase – Alkaline sphingomyelinase ANT – Adenine nucleotide translocator AO – Acridine Orange
APC – Adenomatous polyposis coli aSMase – Acid sphingomyelinase Bak – Bcl-2-antagonist/killer
Bax – Bcl-2-associated X proteinBid – BH3 interacting domain death agonist BRAF – v-RAF murine sarcoma viral oncogene homolog B
BSA – Bovine serum albumin
CAPPs – Ceramide activated protein phosphatases Cat B – Cathepsin B
Cat D – Cathepsin D Cat L – Cathepsin L CDase – Ceramidase
CIN – chromosomal instability pathway CRC – Colorectal carcinoma
DAPI - 4',6-diamidino-2-phenylindole DMH – 1,2-dimethylhydrazine DMSO – Dimethylsulphoxide DNA – Deoxyribonucleic acid
ED50 - Half maximal effective concentration
ER – Endoplasmatic reticulum Eto – Etoposide
FAN – Factor associated with neutral SMase FAP – Familial adenomatous polyposis FITC – Fluorescein isothiocyanate GCS – Glucosylceramide synthase GlcCer – Glucosylceramide GTP – Guanosine triphosphate GTPase – Guanosine triphosphatase HMA – Human microbiota-associated
HNPCC – Hereditary non-polyposis colorectal cancer
IC50 – Concentration needed to reduce the initial response in 50 %
IF – Immunofluorescence
xiii
LAMP – Lysosome associated membrane protein LDH – Lactate dehydrogenase
LMP – Lysosome membrane permeabilization MAPK – Mitogen-activated protein kinase
MMP – Mitochondrial membrane permeabilization MSI – Microsatellite instability pathway
MTT – 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide MYH – MYH gene
nSMase – Neutral sphingomyelinase PBS – Phosphate buffer saline
PTPC – Permeability transition pore complex PVDF – Polyvinylidene difluoride
ROS – Reactive oxygen species RV – Resveratrol
SDS-PAGE – Sodium dodecyl sulfate polyacrylamide gel electrophoresis SCFA – Short chain fatty acids
SLs – Sphingolipids SMase – Sphingomyelinase SPT – Serine palmitoyltransferase SRB – Sulforhodamine B
STS – Staurosporine
TdT – Terminal deoxynucleotidyl transferase TNF – Tumour Necrosis Factor
TUNEL – Terminal dUTP Nick-End Labeling UV – Ultra-violet
1
1
I
NTRODUCTION
2
1.1 Hallmarks of cancer
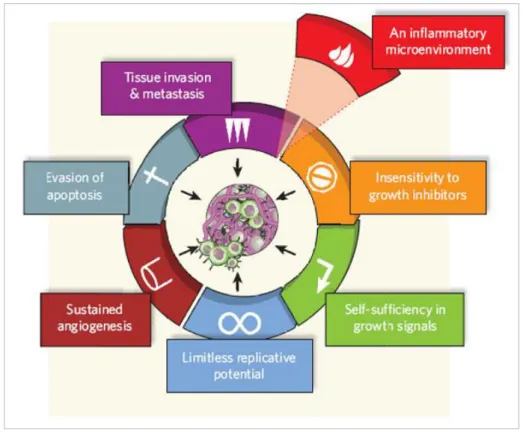
Cancer is a very complex disease, variable in its presentation, development and outcome from a patient to another, and is also a multistep process in which normal cells undergo metabolic and behavioural changes, acquiring capabilities to progressively transform into highly malignant cells. Hanahan and Weinberg (2000) described the hallmarks of cancer, which are six essential alterations in cell physiology that dictate malignant growth (Fig. 1.1): self-sufficiency in growth signals, insensitivity to growth inhibitory (antigrowth) signals, evasion of apoptosis, limitless replicative potential, sustained angiogenesis and tissue invasion and metastasis (Hanahan and Weinberg, 2000). A seventh hallmark of cancer has been added to the list: an inflammatory microenvironment, an essential component resulting from the tumour microenvironment inducing cancer related inflammation. Indeed, activation of oncogenes leads to expression of inflammation-related programs, promoting cancer development (Colotta
et al., 2009; Mantovani, 2009).
Figure 1.1 – The 6 classical hallmarks of cancer plus one, the inflammatory microenvironment: acquired
3
1.2 Colorectal carcinoma
Colorectal carcinoma (CRC) is the third most common cause of cancer death in both sexes in the western world: according to data from GLOBOCAN project (2008) there were an estimated 1 233 711 CRC cases worldwide, accounting for 9.7% of total cancer cases, resulting in more than 600 000 deaths (8% of total cancer deaths). Similar patterns of incidence and mortality are observed in both sexes, incidence is 10% in men and 9.4% in women, while mortality is 7.6% in men and 8.6% in women (Globocan project, 2008, http://www-dep.iarc.fr/). CRC has higher incidence rates in New Zealand, Australia, North America, Europe and more recently Japan, and lower rates reported in Asia and Africa (Boyle and Levin, 2008). In 2008, in Portugal, CRC was the most common form of cancer, with almost 7000 new cases and about 3700 deaths among Portuguese population. In women and men, CRC was the second cause of death by cancer, after breast and prostate cancer, respectively (Globocan project, 2008, http://www-dep.iarc.fr/).
Most of colorectal cancers are sporadic (~70%), arising from somatic mutations and clonal evolution at the tumour site. The remaining cases arise in a familial/hereditary context: 5% show an evident hereditary basis predisposing to the disease, while 25% show only an increased propensity of family members to develop CRC without truly recognized hereditary syndromes (Carli et al., 2009; Rustgi, 2007). The most common form of hereditary CRC is the hereditary non-polyposis colorectal cancer (HNPCC), accounting for 3-4% of all CRC cases, followed by familial adenomatous polyposis (FAP), representing 1% of colorectal tumours. Less than 1% of CRC arise from other syndromes (MYH-associated polyposis, hamartomatous polyposis syndromes and hyperplastic polyposis) and other high-risk conditions (inflammatory bowel diseases, previous diagnoses for cancer of the ovary, endometrium, breast, bile duct, pancreas, stomach) (Boyle and Levin, 2008; Lynch and De La Chapelle, 2003; Rustgi, 2007; Velho, 2010).
4 1.2.1 Colorectal Carcinoma risk factors and prevention
There are two main non-modifiable risk factors for CRC, genetic predisposition and age. However, modifiable risk factors should be considered, as the worldwide variations in colorectal cancer incidence suggest a contribution of dietary and lifestyle factors to the aetiology of this disease. This is supported by migration studies showing that populations moving from low- to high-risk areas rapidly reach the higher level of risk of the adopted country (Boyle and Levin, 2008). Indeed, of all common cancers, colorectal carcinoma is the most susceptible to undergo dietary influence. Higher intake of total energy (Giovannucci and Goldin, 1997), red and processed meat (Chao et al., 2005) and alcohol (Moskal et al., 2007) are associated with an increased CRC risk. On the other hand, higher intake of fruits and vegetables only exert little effect on the risk (Marques-Vidal et al., 2006) and higher intake of dietary fibre can reduce the risk of CRC (Bingham et al., 2003).
Short chain fatty acids (SCFA) are end products of the physiological bacterial fermentation, namely propionibacteria, present on dietary fibre. SCFAs are natural constituents of the colonic lumen and this feature may constitute a key probiotic potential and their potential in cancer prevention and therapy has been investigated (Ruemmele et al., 2003). Colonic endogenous bacteria also produce butyrate, a SCFA that is able to induce apoptosis in colon cancer cells but not in normal cells (Hague et
al., 1995). The use of propionibacteria as a probiotic has also been investigated, once
this genus of bacteria is found in several dairy products and its metabolism relies on anaerobic conversion of carbohydrates and lactic acid to SCFA, particularly propionate and acetate (Britz and Steyn, 1979). Propionibacteria can survive in the human intestine and was found to induce apoptosis in colorectal cancer cells but not in normal cells, at least in part, due to their specific property to produce propionate and acetate (Jan et al., 2002). Propionibacteria and SCFA potential benefits in CRC will be further discussed.
5 1.2.2 Oncogenic activation in Colorectal Carcinoma: KRAS and BRAF Mutations
The MAPK (mitogen-activated protein kinase) pathway regulates cell proliferation, differentiation, senescence and apoptosis (Oliveira et al., 2007). MAPK activation leads to increased cell proliferations and reduced apoptosis, two of the six hallmarks of cancer (Hanahan and Weinberg, 2000).
KRAS gene belongs to the RAS superfamily and is located at chromosome 12p12.1, coding a GTPase. Active RAS proteins are found at plasma membrane, acting as mediators in the transduction of extracellular stimuli, resulting in the activation of signalling cascades involved in the regulation of cell growth, proliferation, differentiation, survival and death (Aoki et al., 2008). Active form of KRAS needs to be GTP-bound, and mutations in these genes often cause RAS to accumulate in the active GTP-bound state, resulting in sustained activation of the downstream signalling pathways (Bos, 1989).
BRAF belongs to RAF family of serine/threonine protein kinases. These proteins transduce signals from activated RAS in the cell surface to the nucleus (Kolch, 2000). In human cancers, mutations in RAF genes promote the constitutive activation of RAF proteins, leading to abnormal stimulation of the MAPK cascade (Davies et al., 2002). The most common BRAF mutation, found in about 80% of the mutated cases, is a thymidine to adenosine transversion at nucleotide 1796 (T1796A) that converts valine 600 to glutamate (V600E) (Davies et al., 2002; Garnett and Marais, 2004). This mutation induces proliferation and transformation, allowing cells to grow as tumours (Ikenoue et al., 2004), showing a 138-fold transforming and oncogenic activity over wild-type BRAF (Davies et al., 2002; Velho, 2010).
It is thought that at least 50% of colorectal cancers have a deregulation of the MAPK pathway (Fang and Richardson, 2005). Two major oncogenes are implicated in sporadic colorectal carcinogenesis, KRAS and BRAF mutations, which are alternative (Oliveira et al., 2007). Mutations on KRAS and BRAF genes are frequently found in malignant and pre-malignant colorectal lesions (Rajagopalan et al., 2002). Mutations of the KRAS proto-oncogene are an early event in development of CRCs (Bos et al., 1987). KRAS oncogene is mutated in 21% of all human sporadic cancers, including in about 30% of CRCs cases (Fang and Richardson, 2005; Oliveira et al., 2007), and
6 BRAF mutations are found in 20% of all human cancers, including in about 10% CRCs cases (Ahlquist et al., 2008; Oliveira et al., 2007; Velho, 2010).
1.3 Apoptosis
Apoptosis is a mechanism of programmed cell death, which is tightly regulated under normal conditions, being essential to fundamental processes such as development, immune response and tissue homeostasis. There are two classic pathways described for apoptosis (Fig. 1.2): the intrinsic pathway and the extrinsic pathway. The intrinsic pathway is activated by many different types of cellular stress, including UV irradiation, γ-irradiation, some chemotherapeutic drugs, reactive oxygen species (ROS) and radicals generated by the cell as a by-product of normal metabolism, and controlled by members of the Bcl-2 family (pro-apoptotic and anti-apoptotic proteins). Its key points are the release of cytochrome c from mitochondria and caspase activation (Watson, 2004; Yang
et al., 2009). The extrinsic pathway of apoptosis is activated by the binding of ligands to
death receptors as Fas (APO-1/CD45) and other members of the tumour necrosis factor (TNF) receptor family, resulting in caspases-8 activation (Danial and Korsmeyer, 2004).
7 1.3.1 The role of apoptosis in normal colorectum and colorectal carcinogenesis
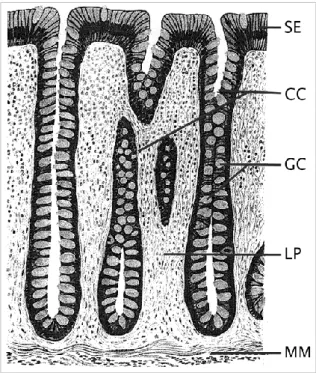
Colonic epithelial cells are configured in deep invaginations into the wall of the colon called crypts (Kinzler and Vogelstein, 2002) (Fig. 1.3), and apoptosis has an important role in determining and maintaining the normal colonic epithelia architecture (Watson, 2004; Watson and Pritchard, 2000). In normal colonic epithelia, two types of apoptosis can occur: spontaneous apoptosis that occurs continuously in normal, unstressed colon, and stress-induced apoptosis, which occurs in response to toxic stimuli (Watson and Pritchard, 2000). A delicate balance between cell proliferation at the base of the crypt and cell death at the top of the crypt, should be kept in order to maintain the normal architecture of colonic crypts and gastrointestinal homeostasis (Yang et al., 2009). The transformation of colorectal epithelium to carcinoma usually proceeds through a series of pathological progressions to produce colorectal polyps, adenomas, adenoma-containing carcinomas and adenocarcinomas (Kukitsu et al., 2008), and is associated with a progressive inhibition of apoptosis (Bedi et al., 1995).
Figure 1.3 – Morphology of normal colon tissue. SE – surface epithelium, CC – colon crypts, GC –
goblet cells, LP – lamina propria, MM – muscularis mucosa (MM). The crypts open to the surface epithelium—in this cross section, some of the crypts appear partially or below the surface. Adapted from Kinzler and Vogelstein (2002).
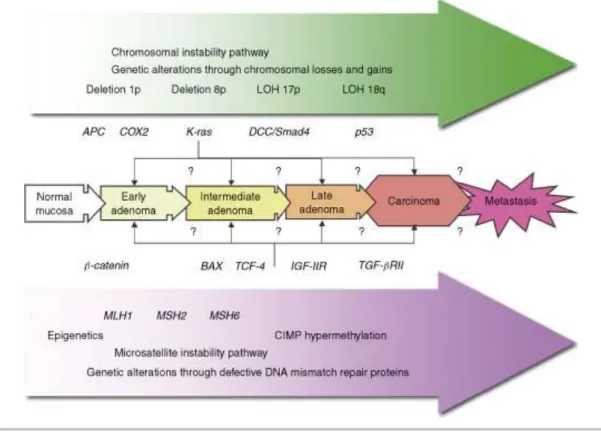
8 Initially, CRC was thought to follow an adenoma-carcinoma sequence, a multistep process involving several genetic changes (Fearon and Vogelstein, 1990). More recent studies showed that although a large number of genes are mutated in colorectal tumours, only a small proportion is responsible for driving tumour progression and growth (Wood et al., 2007). Besides, it has been suggested that most colorectal tumours develop via three distinct pathways: suppressor, mutator and methylator (Fig. 1.4) (Worthley et al., 2007). Suppressor pathway or chromosomal instability pathway (CIN) is present in approximately 70-85% of CRC (Grady, 2004; Komarova et al., 2002), and is associated with mutation in APC (adenomatous polyposis coli) gene or loss of 5q, mutation of KRAS, loss of 18q and deletion of 17p which contains p53 gene (Grady, 2004). Mutator pathway or microsatellite instability pathway (MIS) occurs both in hereditary and sporadic CRC but through a different mechanism. In HNPCC a germline mutation in a mismatch repair enzyme causes MSI, while in sporadic CRC the promoter of a mismatch repair gene undergoes epigenetic silencing by methylation (Söreide et al., 2006; Watson, 2004). In the methylator pathway, specific genes are affected by epigenetic events, namely DNA methylation (Worthley et al., 2007), resulting in CIMP (CpG Islands Methylator Phenotype) cancers (Toyota et al., 1999).
Figure 1.4 - Characteristics of the two major pathways in colorectal carcinogenesis: CIN pathway and
9 The increasing knowledge of apoptotic pathways allows investigation of these processes and their regulators as therapeutic targets in cancer, and efforts are being made to improve the effectiveness or specificity of currently available therapy. Drugs that directly induce apoptosis present several advantages when compared to those that do not directly induce it. Apoptosis results in removal of dead cells by phagocytosis without inflammatory process or damage to the surrounding normal tissue. Drugs that directly induce apoptosis are less mutagenic and can eliminate cancer cells totally, avoiding tumour recurrence, unlike cytostatics that usually leads to tumour relapse (Yang et al., 2009).
1.3.2 Acetic acid/acetate-induced cell death in S.cerevisiae and in colorectal carcinomas
Saccharomyces cerevisiae has been widely used as eukaryotic model for diverse
studies in several fields of biological sciences, including the study of human apoptosis. Acetic acid is a normal end product of the alcoholic fermentation carried out by S.
cerevisiae, and it was found to induce an apoptotic phenotype in yeast involving
mitochondria and cytochrome c release (Ludovico et al., 2002; Ludovico et al., 2001). Acetic acid was also found to induce mitochondrial outer membrane permeabilization (MOMP), which is a critical step in the apoptotic pathway and seems to require ADP/ATP carrier (AAC) proteins, which are yeast orthologues of the mammalian adenine nucleotide translocator (ANT) (Pereira et al., 2007). In mammalian cells, MOMP occurs, at least in part, due to the opening of the permeability transition pore complex (PTPC), located at the contact site between the two mitochondrial membranes, and is composed by several proteins, including voltage-dependent anion channel (VDAC) in the outer membrane, and the ANT in the inner membrane (Zoratti and Szabò, 1995). After apoptotic stimuli in mammalian cells, ANT protein forms a non-specific pore, leading to diffusion of large molecules and consequently to permeabilization of outer membrane and release of apoptogenic proteins into the cytosol (Vieira et al., 2000). Acetic acid-induced cell death in yeast was also found to trigger translocation of Pep4p, an orthologue of human cathepsin D, from the vacuole to the
10 cytosol (Pereira et al., 2010). This observation suggests that partial release of cathepsins from the lysosomes might be an early apoptotic event in mammalian cells, as it will be referred later on.
As previously mentioned, SCFA seem to have a potential role in colorectal carcinoma cells apoptosis (Jan et al., 2002). Considering the results of acetic acid-induced cell death in S. cerevisiae and the fact that propionibacteria are able to survive in human intestine and to produce acetate which induces apoptosis in CRC cells (Jan et
al., 2002), this issue should be exploited in order to provide a new therapeutic approach
for CRC. Only few works have been published in this field and the mechanisms underlying this hypothesis are not understood.
In 2002, Jan and co-workers showed that two species of propionibacteria, P.
acidipropionici and P. freudenreichii can induce cell death in HT-29 and Caco-2
carcinoma cell lines apparently via SCFA, namely propionate and acetate. They also reported that the two SCFA can induce cell death separately, with a half maximal effective concentration (ED50)of 11 mM for propionate and 20 mM for acetate, in
HT-29 cells. Cell death pathway was reported to be apoptosis, as SCFA induced cleavage of pro-caspase 3, generating the active form of caspase 3, and features of apoptotic cell death, such as nuclei shrinkage, chromatin condensation and nuclei fragmentation into apoptotic bodies, were also observed (Jan et al., 2002). In this work, it was also demonstrated the involvement of mitochondria in apoptosis, as reported by the increased mitochondrial inner membrane permeability and the enhanced generation of ROS. The use of mitochondrial inhibitors of apoptosis as Bcl-2 and vMIA (viral mitochondrial inhibitor of apoptosis, a protein encoded by human cytomegalovirus) partially prevented propionibacterial SCFA-induced cell death, suggesting the critical role of mitochondria in this death process (Jan et al., 2002). These authors also identified ANT as a potential SCFA target within the PTPC, since vMIA selectively interacts with ANT.
As the cell microenvironment may be determinant for the type of cell death induced by toxic stimuli and in the lumen of human colon pH ranges from 5.5 to 7.5, it is important to determine pH influence on propionibacterial SCFA-induced cell death. Lan and co-workers (2007) concluded that SCFA produced by P. freudenreichii trigger apoptosis at pH 7.5 while at pH 5.5 a necrotic process was observed in HT-29 cells. At pH 7.5 propionibacterial SCFA induced cell cycle arrest in the G2/M phase, and morphological characteristics of apoptotic cell death such as membrane blebbing,
11 chromatin condensation and fragmentation, phosphatidylserine exposure and formation of apoptotic bodies, were observed (Lan et al., 2007b).
Propionibacteria, via its metabolites propionate and acetate, have been proved to induce apoptosis in colorectal carcinoma cell in vitro (Jan et al., 2002; Lan et al., 2007b) but its pro-apoptotic potential in vivo, with selectivity to only malignant cells and not to healthy ones, has also been investigated (Lan et al., 2008). Colonic epithelial apoptosis and proliferation was evaluated in human microbiota-associated (HMA) rats consuming P. freudenreichii TL133 on a daily basis, 48 hours after the induction of carcinogenesis with 1,2-dimethylhydrazine (DMH). TL133 strain of P. freudenreichii is able to survive and to be metabolically active in the gastrointestinal tract of HMA rats (Lan et al., 2007a), and was proved to induce apoptosis of colon cells only in animal treated with DMH and not in healthy ones, suggesting that apoptosis induction specifically targets damaged cells. These data indicate that propionibacteria might have a protective role against colon cancer cells as it might help in the elimination of damaged cells by apoptosis in the colonic epithelium (Lan et al., 2008). Altogether, these data point to a useful role of propionibacteria as a probiotic, and their metabolites, propionate and acetate, as powerful agents for prevention and therapy of CRC.
1.4 Lysosome membrane permeabilization as a lysosomal pathway of
cell death
Apoptosis, or lack of it, is critical to tumorigenesis (Letai, 2008). Neoplastic cell expansion could be attributable to decreased cell death rather than rapid proliferation, once defects on apoptotic pathways allow neoplastic cells to survive beyond senescence, providing protection from hypoxia and oxidative stress as the tumour expands. Cell death pathways are blocked at different levels of the signalling cascade by upregulation of anti-apoptotic proteins and/or downregulation or dysfunction of pro-apoptotic molecules (Fulda, 2009). Therefore, evasion of apoptosis, as a hallmark of cancer, promotes tumour formation and progression as well as treatment resistance. Although cancer cells may block classical apoptotic pathways, cell death can still occur through the release of lysosomal enzymes (Kirkegaard and Jäättelä, 2009).
12 For many years, the role of lysosomes in cell death was thought to be exclusive of necrotic and autophagic pathways, taking part of the apoptotic process solely in digestion of engulfed apoptotic bodies, and lysosomal proteases functions were limited to nonspecific intracellular protein degradation occurring within the lysosome (Ferri and Kroemer, 2000; Leist and Jäättelä, 2001). However, the ‗lysosomal pathway of apoptosis‘ is nowadays a widely accepted concept as partial lysosomal permeabilization with subsequent release of proteolytic enzymes into the cytosol, contributing to the cellular signalling pathways, has been described in several models of apoptosis (Guicciardi et al., 2004). The discovery of lysosomal cell death pathway was a difficult task, since lysosomal ultrastructure appears intact in apoptotic cells analyzed by electron microscopy (Brunk et al., 2001). Initially, lysosomal rupture was considered as an all-or-nothing switch during late stages of uncontrolled necrosis (De Duve, 1983), but more recent studies demonstrated that lysosomes with normal ultrastructure may have leaked part of their enzymes, and that partial lysosome membrane permeabilization (LMP) can trigger apoptosis and apoptosis-like cell death (Guicciardi et al., 2004). Therefore, the magnitude of lysosomal permeabilization determines the type of cell death mediated by lysosomal enzymes: a complete breakdown of the organelle with release of high concentration of lysosomal enzymes results in unregulated necrosis, while partial, selective permeabilization triggers apoptosis (Bursch, 2001; Turk et al., 2002). Once released into the cytosol, lysosomal enzymes contribute to the execution of the apoptotic program, either by direct cleavage of key cellular substrates, or by acting together with caspases (Leist and Jäättelä, 2001).
1.4.1 Lysosomal enzymes
LMP followed by release of lysosomal contents, especially cathepsins, to the cytosol seems to be the critical step of the lysosomal death pathway. Cathepsins are the best studied lysosomal enzymes and are classified according to their active site amino acid, i.e. cysteine, aspartate and serine cathepsins (Rawlings and Barrett, 1999). There are several human cathepsins identified, but the most relevant are the cysteine cathepsins B and L (Cat B and Cat L) and the only lysosomal aspartic protease, cathepsin D (Cat D),
13 because they are the most abundant in the lysosome (Turk et al., 2002) and they remain active at neutral pH, although their optimal activity is at acidic pH (Boya and Kroemer, 2008). The processing of the proenzymes into the catalytic active form usually occurs within the lysosome (Ishidoh and Kominami, 2002) and when released into the cytosol these enzymes are in their active, monomeric form, and are stable at physiologic pH. Cat B is one of the most stable proteases at neutral pH, and is essential in different models of apoptosis. Cat L is the less stable protease at neutral pH, but is also involved in several models of apoptosis. Finally Cat D has been implicated in apoptosis induced by staurosporine (Bidere et al., 2003; Johansson et al., 2003), interferon-γ, Fas/CD95/APO-1 and TNF-α (Demoz et al., 2002), oxidative stress (Roberg et al., 1999), sphingosine (Kågedal et al., 2001) and p53 (Wu et al., 1998).
Besides lysosomal enzymes such as cathepsins, protein components of the lysosomal membrane also mediate essential functions in the lysosome, including the acidification of the lysosomal lumen, transport of amino acids, fatty acids and carbohydrates resulting from the hydrolytic degradation, as well as other nutrients generated by lysosomal hydrolases (Eskelinen et al., 2003). Lysosomal membrane proteins may also be involved in the interaction and fusion of the lysosomes with themselves and with other cell components such as endosomes, phagosomes and the plasma membrane (Fukuda, 1991). Lysosome associated membrane protein-1 (LAMP-1) and -2 (LAMP-2) are the major components of the lysosomal membrane, constituting ~50% of the membrane proteins in this compartment (Eskelinen, 2006). It has been suggested that these glycoproteins can modulate lysosomal membrane sensitivity to an apoptotic stimuli, since they act as barriers against the hydrolytic activity of the lysosomal enzymes, preventing accidental release of lysosomal constituents into the cytosol (Fehrenbacher et al., 2008; Johansson et al., 2010). Oncogene-induced reduction in LAMP-1 and -2 can be caused by Cat B overexpression, and is associated with increased susceptibility to LMP in response to photo-oxidation and the anti-cancer drug siramesine. Upregulation of lamp-1 and -2 mRNAs were observed in various human cancers (Furuta et al., 2001; Ozaki et al., 1998), and might constitute a compensatory mechanism for the deleterious effect of the reduced half-life of LAMP-1 and -2 in cancer cells overexpressing lysosomal cysteine cathepsins. In colorectal neoplastic tissues, either adenoma or cancer, LAMP molecules are highly expressed, compared to nonneoplastic counterparts, indicating that the expression of LAMP molecules is related to neoplastic progression. LAMPs may be correlated not only with
14 malignant transformation of tumours cells but also with their property of invasion (Furuta et al., 2001).
1.4.2 Mechanisms of lysosome mediated apoptosis
Although lysosomes and their enzymes are known to be associated with apoptotic cell death, the exact mechanisms and possible cross-talks with other apoptotic pathways are still unclear. Apparently, lysosomal proteases trigger apoptosis not via a single specific pathway, but rather multiple molecular pathways, including those controlled by traditional apoptosis mediators, like caspases and proteins from Bcl-2 family (Guicciardi et al., 2004). Evidences support that LMP seems to be an early event in the apoptotic cascade (Bidere et al., 2003; Boya et al., 2003a; Roberg et al., 1999), and a possible mechanism might be direct cleavage and activation of caspases by lysosome proteases released into the cytosol. Even though main effector caspases have been proved to be poor substrates for several cathepsins in vitro, it has been demonstrated that a human cathepsin L-like protease and a trypanosomal cysteine cathepsin related to cathepsin L are able to cleave procaspase-3/-7 in cell free systems and cell culture (Stoka et al., 2001). Cat B has also been proved to cleave procaspase-2 in vitro, though less effectively than when it is cleaved during auto-activation process, yet inducing significant mitochondrial cytochrome c-releasing activity (Guicciardi et al., 2004).
Direct activation of caspases by cathepsins should be considered, but growing evidences suggest that lysosome proteases rather promote apoptosis by inducing mitochondrial dysfunction, associated with release of pro-apoptotic factors (Bidere et
al., 2003; Boya et al., 2003a; Zhao et al., 2003). Following mitochondrial damage, ROS
generated and others factors could also send feedback signals to the lysosome, resulting in further lysosomal breakdown and amplification of the apoptotic cascade (Zhao et al., 2003). Mitochondrial dysfunction through lysosomal protease release can be caused directly, for instance, by activation of phospholipase A2 with production of free arachidonic acid, which are both capable of destabilizing mitochondrial membranes (Zhao et al., 2003), or indirectly by inducing Bax and Bak to adopt their apoptotic conformation and to insert into the mitochondrial membrane, triggering mitochondrial membrane permeabilization (MMP) and consequent cytochrome c release (Bidere et al.,
15 2003; Boya et al., 2003a). Indeed, is more likely that lysosomal enzymes do not directly induce mitochondrial dysfunction. This is supported by a study in human lymphocytes reporting that lysosomal proteases translocation into the cytosol precedes mitochondrial permeabilization and the release of Cat D into the cytosol triggers Bax activation and its relocation to the mitochondria, resulting in release of the pro-apoptogenic factor AIF (apoptosis-inducing factor) and the apoptotic phenotype (Bidere et al., 2003). The requirement of Bax and/or Bak was confirmed by demonstrating that mouse embryonic fibroblasts from Bax/Bak double knockout are resistant to MMP when treated with LMP inducers, suggesting that lysosome-mediated apoptosis requires the activation of a Bax/Bak-dependent pathway rather than directly induce mitochondrial dysfunction (Boya et al., 2003a; Boya et al., 2003b).
The BH3-only protein Bid might also be involved in LMP. It has been demonstrated that Bid is cleaved and translocated to mitochondria following lysosomal disruption by lysosomotropic agents (Cirman et al., 2004). Bid can be processed and activated by several cysteine cathepsins at cytosolic pH, and Cat D has been suggested to cleave and activate Bid in the acidic environment of the lysosome after TNF-R1 internalization (Heinrich et al., 2004; Schneider-Brachert et al., 2004). Cat B- or Cat D-mediated cleavage of Bid seems to be a connection between MMP and LMP, but Bid involvement in LMP is still controversial. Bid can be cleaved either by lysosomal enzymes or by non-lysosomal enzymes like caspase-8 (Guicciardi et al., 2004), and other lysosomal enzymes rather than cathepsins might be responsible for its cleavage (Reiners et al., 2002). It has also been suggested that Bid is not required for cathepsin-mediated apoptosis in an in vivo model of cell death where a cystatin B/Bid double knockout mice showed that absence of Bid did not rescue the neurological phenotype caused by constitutional activation of cathepsins, due to the lack of the endogenous inhibitor cystatin B (Houseweart et al., 2003).
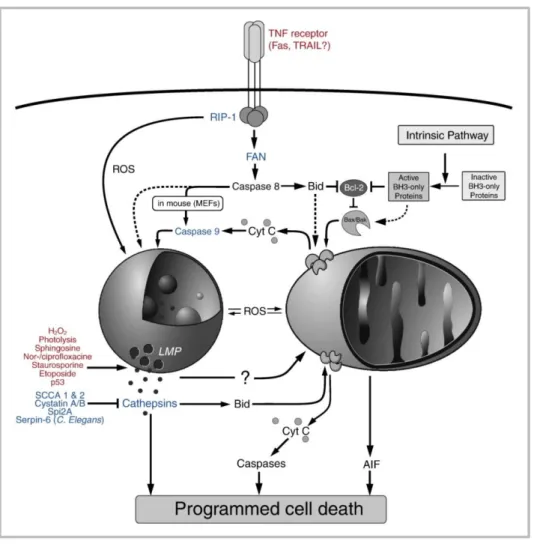
In conclusion, lysosome-mediated apoptosis may trigger the classical MMP-caspase pathway as well as the MMP- and caspase-independent pathways (Fig. 1.5), depending on the cellular context, including the cell type, the immortalization status and the genetic background (Boya and Kroemer, 2008).
16 Figure 1.5 – Lysosomal involvement in cell death signalling. Stimuli that induce LMP are printed in red.
Proteins that modulate the lysosomal death pathways are printed in blue. Adapted from Kirkegaard and Jäättelä (2009).
1.4.3 Methods for detection of lysosome membrane permeabilization
As the evidence of a ―lysosomal pathway of apoptosis‖ accumulates and becomes widely accepted, methods for studying lysosomes and lysosomal proteases in apoptosis were developed. Partial LMP seems to be the initiator and executioner of the apoptotic program (Guicciardi et al., 2004). Its distinctive signal is the translocation of soluble components (including enzymes) from the lysosomal lumen to the cytosol that can be measured by a variety of techniques (Boya and Kroemer, 2008).
Most of the methods are based on the difference in pH between lysosomes and surrounding milieu, that has a neutral pH in contrast to lysosomes which have an
17 average of pH of 4.6 to 5.0, due to the proton-pumping vacuolar V-ATPases, which produce a proton gradient over the lysosome membrane (Ivanova et al., 2008).
The simplest way to assess LMP is to use lysosomotropic fluorescent dyes which are weak bases that become charged after attracting one or more protons. This protonated form is not membrane permeable, remaining trapped in the acidic compartments (Ivanova et al., 2008). Lysotracker (a trademark of Molecular Probes) and Acridine Orange (AO) are frequently used for this purpose. AO used in the present work, is a lysosomotropic weak base that accumulates, in its protonated form, within lysosomes emiting red fluorescence under blue excitation light. When it is released to the cytosol, after lysosome membrane permeabilization, emits green fluorescence (Trincheri et al., 2007).
However, an increase in lysosomal pH might not necessarily be a consequence of LMP, but might be a consequence of specific effects of agents on V-ATPases or of an increased proton leakage (Ivanova et al., 2008), therefore, an alternative method to verify LMP is required.
An early molecular marker for lysosome integrity is to measure the activity of the lysosomal enzyme β-hexosaminidase, but it is prone to produce artifacts, once is very large and also present in the lysosome membrane-bound form. The most preferred method for LMP detection is the use of lysosome proteases as cytosolic markers, because they are smaller than β-hexosaminidase and they are often associated with cell death progression and very easy to assay (Ivanova et al., 2008).
Translocation of cathepsins from lysosomes to the cytosol can be detected by immunoblots of subcellular fractions, tipically cytosolic fraction versus heavy membrane fraction, which include mitochondria and lysosomes (Boya and Kroemer, 2008).
Immunofluorescence techniques, using antibodies against cathepsins are also able to reveal their redistribution from lysosomes to the cytosol. In untreated control cells, cathepsins-specific immunostainings show cytoplasmic punctuate structures; after LMP, immunofluorescence detection of cathepsins reveals a diffuse staining throughout the entire cell (Boya et al., 2003a).
Others methods for detection of cytosolic cathepsin translocation have been described, including: measurement of protease activities using cathepsin-specific substrates (Groth-Pedersen et al., 2007; Yin et al., 2005); transfection of cells with fusion constructs encoding cathepsins coupled to fluorescent proteins (Broker et al.,
18 2005; Werneburg et al., 2007); pulse-chase experiments, by loading lysosomes with fluorescent dextrans and observing the cytosolic release of the dye after LMP (Bidere et
al., 2003).
1.4.4 Lysosome membrane permeabilization and cancer
The lysosomal pathway of apoptosis was first identified in pathological situations, such as the cathepsins involvement in the Central Nervous System (CNS) apoptosis following ischemia and during neurodegenerative process, the contribution to TNF-α signalling in the liver, liver damage following bile duct obstruction, and cold-ischemia/warm-reperfusion injury (Guicciardi et al., 2004). Tumorigenic transformation of cells is another pathological condition favouring lysosomal pathway activation. Transformation leads to several changes in the lysosomal compartment promoting increases in lysosomal volume and total protease activity and enhanced secretion of proteases (Tardy et al., 2006), inducing changes in the trafficking and subcellular localization of the cathepsins B, D and L (Kirkegaard and Jäättelä, 2009). Such alterations become pro-oncogenic when enhanced secretion of cathepsins initiates proteolytic pathways that increase neoplastic progression (Mohamed and Sloane, 2006). Indeed, an increase in cysteine cathepsin activity during tumour development was found to be important for tumour angiogenesis, cell proliferation, tumour growth and invasion (Joyce et al., 2004). Overexpression of cathepsins often occurs in human cancers, and high levels of their expression can be associated with increased risk of relapse and poor prognosis (Palermo and Joyce, 2008).
Besides the tumour-promoting effects of cathepsins, mentioned above, there are evidences that these protease may function as tumour suppressors as well (Lopez-Otin and Matrisian, 2007). This opposing role of cathepsins depends on the context: if cathepsins are released intracellularly, they contribute to cancer cell death, but if they are released extracellularly, they are pro-oncogenic, breaking down the extracellular matrix, and stimulating angiogenesis and migration (Fig. 1.6) (Kirkegaard and Jäättelä, 2009). For example, transformation has been shown to increase mouse embryonic fibroblasts susceptibility to lysosome-dependent cell death induced by anticancer agents (Fehrenbacher et al., 2004).
19 The increased susceptibility of cancer cells lysosomes to LMP is not completely understood. There are several possibilities that still need experimental verification, but all possibilities indicate that lysosomes from cancer cells are particularly susceptible to therapeutic induction of LMP: cancer cells have larger lysosomes than normal cells that may be more fragile than normal-sized lysosomes (Glunde et al., 2003; Ono et al., 2003); higher metabolic rates and increased turnover of iron-containing proteins in cancer cells lead to the accumulation of iron in lysosomes, with consequent iron-mediated sensitization to ROS-induced LMP (Eaton and Qian, 2002); cancer cells often produce elevated ROS levels and the associated higher rate of spontaneous cathepsin release from lysosomes may promote cell death induction (Gyrd-Hansen et al., 2004).
Therefore, transformation-induced changes in lysosomal compartment might have a great potential in cancer therapy, in the context of induction of cell death. During malignant transformation, cells often acquire defects in the classical caspase-dependent apoptosis pathway, including reduced expression of pro-apoptotic signalling and overexpression of anti-apoptotic proteins. Tumour cell death becomes caspase-independent, but still can occur via lysosomal cell death, particularly during the early stages of tumorigenesis when cancer cells are more sensitized to death stimuli (Kirkegaard and Jäättelä, 2009). Targeting lysosomal cell death may be an attractive and alternative therapeutic strategy to overcome the block in classical apoptotic pathways in cancer cells.
20 1.4.5 Lysosome membrane permeabilization and cathepsin D expression in colorectal carcinomas
There are few published works concerning the role of LMP induced cell death in CRC. Trincheri and co-workers (2007) reported that Resveratrol (RV), a naturally occurring polyphenol with potential chemopreventive and chemotherapy activities in colorectal cancer, triggers the caspase-dependent intrinsic pathway of apoptosis. These authors provide evidence that Cat D, and not Cat B or Cat L, mediates RV cytotoxicity. In DLD1 and HT29 cell lines, RV promotes accumulation of mature Cat D, inducing lysosome leakage with increased cytosolic immunoreactivity of Cat D. Moreover inhibition of Cat D prevents apoptotic events such as Bax oligomerization, MMP, cytochrome c release and caspase 3 activation (Trincheri et al., 2007).
Cat D protein levels in colorectal tumours show divergent expression patterns, suggesting a complex regulation and function of this protease (Iacobuzio-Donahue et
al., 2004). Iacobuzio-Donahue and co-workers analyzed Cat D protein patterns in 59
colorectal tumours, and concluded that the average Cat D protein content was not different in colorectal tumours comparing with normal mucosa, but individual tumours demonstrated marked changes in Cat D expression levels. Measurement of protein content of tumour versus normal pairs on Western blots revealed loss of Cat D in more than 50% of colorectal tumours. However, 2-fold increases in Cat D protein levels were also observed in ~1/3 of tumours, supporting the concept that colorectal cancers develop via divergent molecular pathways and that Cat D may function differently in different cancers. These authors also showed that regarding the stage of the tumour, it has been reported that Cat D expression decreased more frequently in stage III and IV cancers (64%) than in adenomas and locally invasive carcinomas (stage I and II) (40%), suggesting that decreased levels of Cat D protein expression is common in colorectal tumours and the loss of Cat D expression may facilitates tumour development. The variable Cat D levels on CRCs may also be explained by the differential regulation of Cat D expression by mutant versus wild-type p53, since the later was shown to transactivate Cat D expression via two p53-binding sites in the Cat D promoter. The results obtained in this study, where 50% of colorectal cancers have been shown to have decreased Cat D expression, might be explained in part by the loss of normal p53, which occurs in approximately 50% of colorectal cancers. Significant loss and gain of
21 chromosome 11, where Cat D gene is located, in different colorectal tumours may also explain the observations on Cat D expression patterns (Iacobuzio-Donahue et al., 2004). Another study on surgical specimens of 100 primary colorectal adenocarcinomas with different grades of differentiation revealed that immunoreactivity for Cat D was negative in the cytoplasm of normal colorectal epithelial cells adjacent to carcinoma (Yilmaz et al., 2003). Cat D expression in carcinoma cells was present in 68% of the cases, and Cat D positivity was higher when the grade of differentiation is higher.
All these results suggest an important role of the lysosome, LMP and Cat D during colorectal carcinogenesis that need to be investigated.
1.5 Ceramide
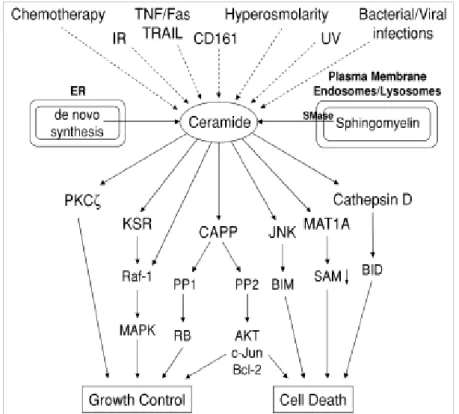
Ceramide is the metabolic and structural precursor for complex sphingolipids (SLs); its basic structure comprises a sphingoid base attached to a fatty acid chain, varying in length from C14 to C26 (Saddoughi et al., 2008). Besides, ceramide is an important second messenger, regulating vital cellular functions (Arana et al., 2010). Endogenous levels of ceramide are regulated by complex and integrated pathways involving specialized enzymes (Fig. 1.7). There are three main metabolic pathways to form ceramide: (1) the de novo pathway, which occurs in the endoplasmic reticulum (ER) and possibly in ER-associated membranes (Hannun and Obeid, 2008), where serine and palmitoyl-CoA condense, catalyzed by serine palmitoyltransferase (SPT) (Nagiec et al., 1996); (2) the sphingomyelinase pathway, consisting in the hydrolysis of sphingomyelin (SM) by activation of sphingomyelinases (SMases), producing phosphocoline and ceramide (Jones and Hannun, 2005); (3) the salvage pathway or SLs recycling pathway in which ceramide is originated from the catabolism of complex SLs that eventually break down into sphingosine in the lysosomes (Kitatani et al., 2008).
22 1.5.1 Ceramide and apoptosis
Ceramide has been suggested to have a role in apoptosis, as many mediators of apoptosis, such as chemotherapeutic agents, CD95, TNF-α, growth factor withdrawal, hypoxia, or DNA damage, are, as well, regulators of ceramide generation (Pettus et al., 2002). Several studies indicate that changes in ceramide endogenous levels in response to these agents precede the apoptotic cascade (Dbaibo et al., 1997), and alterations in ceramide metabolism enzymes that increase endogenous levels of ceramide result in apoptosis and/or growth arrest (Abe et al., 1995; Bielawska et al., 1996).
Figure 1.7 – Ceramide central role in sphingolipids metabolism. Adapted from Saddoughi et al. (2008).
The activation of SMases provides a strategy for the fast generation of ceramide in response to several stimuli, confirming the role of ceramide as an active second messenger in cell death pathways (Kolesnick and Kronke, 1998; Marchesini and Hannun, 2004). SMases can be characterized according to their optimal pH as acid SMase (aSMase), neutral SMase (nSMase) and alkaline SMase (alkSMase) (Bartke and
23 Hannun, 2009). Although some studies have reported the involvement of nSMase and alkSMase in apoptosis induction (Clarke and Hannun, 2006; Liu et al., 2006), most studies focused the role of aSMase, demonstrating its critical role in response to many apoptotic stimuli in almost any mammalian cells (Schenck et al., 2007).
The precise mechanisms by which ceramide induces apoptosis are not completely understood, but several possibilities have already been described. The regulation of biological functions occurs through sphingolipids-protein interactions (Morales et al., 2007; Snook et al., 2006), and most frequently, these targets of ceramide constitute protein phosphatases and kinases implicated on regulation of cancer signalling pathways, such as Akt, Protein Kinase C (PKC), MAP kinases or phospholipase D (Hannun and Obeid, 2002; Ogretmen and Hannun, 2004) (Fig. 1.8). Ceramide mediates the regulation of protein phosphatase-1 and -2 (PP1 and PP2A)-family enzymes, also referred to as ceramide activated protein phosphatases (CAPPs) (Wolff et al., 1994). Upon CAPPs activation by ceramide, downstream targets such as Bcl-2 family proteins, cyclin dependent kinases, Rb, and c-Myc onco-protein, are also regulated (Ogretmen and Hannun, 2004). Cathepsin D is another ceramide binding protein, which is activated via ceramide interaction (Heinrich et al., 2004).
24 1.5.2 Ceramide and lysosome membrane permeabilization
Ceramide and its metabolism may have an important role in LMP induction. It has been shown that exogenous C6 ceramide does not permeabilize lysosomes in vitro (Werneburg et al., 2002), but ceramide generated by the endolysosomal aSMase binds and activates the lysosomal Cat D, by inducing the autocatalytic proteolysis of the 52 kDa pre-pro cathepsin D to form the enzymatically active 48/32 kDa isoforms of Cat D, which, in turn, are responsible for the proteolytic activation of other lysosomal proteins (Heinrich et al., 1999). In TNF-induced cell death, Cat D is activated through direct binding to ceramide, and its activation is dependent on functional aSMase expression. Active Cat D cleaves Bid and leads to activation of caspase 9/3 (Heinrich et al., 2004). TNF stimulates ceramide generation by activating nSMase at the plasma membrane and aSMase in the lysosome, but so far only nSMase has been connected to LMP through the factor associated with neutral SMase (FAN), since FAN does not only mediate TNF-induced ceramide production but also contributes to caspase-8 processing and cell death (Segui et al., 2001).
Ceramide has also been reported to be involved in CRC cells apoptosis by targeting Cat D. It has been suggested that in HT-29 cells increased levels of ceramide, induced by C2-ceramide, inhibited the transport and processing of Cat D. An inactive analogue
of C2-ceramide had no effect on Cat D transport and maturation. The effect of ceramide
on Cat D transport and processing occurs before the appearance of classical features of apoptosis, suggesting that alterations of Cat D transport along the secretory pathway due to increased levels of ceramide is an early event in cells undergoing apoptosis (De Stefanis et al., 2002)
Sphingosine, which is generated from processing of ceramide catalyzed by the lysosomal enzyme acid CDase, has also been proposed as a mediator of LMP, since it can act as a detergent, directly destabilizing the lysosomal membrane (Andrieu-Abadie and Levade, 2002). Due to its lysosomotropic properties, sphingosine accumulates within the lysosomes where it is protonated and can permeabilize the membrane via a detergent mechanism, promoting the relocation of lysosomal enzymes to the cytosol (Kågedal et al., 2001). TNF-α also regulates intracellular levels of sphingosine, by cathepsin B-mediated downregulation of shingosine kinase, an enzymes that converts the pro-apoptotic sphingosine to an anti-apoptotic sphingosine 1-phosphate (Taha et al.,
25 2005). Cat B activity results in accumulation of sphingosine in lysosomes, particularly in the liver where it has been implicated in LMP, also explaining the requirement of Cat B for an efficient LMP in TNF-treated hepatocytes (Werneburg et al., 2002).
1.5.3 The role of ceramide in colorectal carcinomas
Ceramide has been described to induce apoptosis in tumour cells (Carpinteiro et al., 2008), including human colorectal carcinoma cells (Zhang et al., 2006).
It has been suggested that ceramide is closely related with tumorigenesis and tumour progression in digestive tract, since a decreased in ceramide levels increases the risk of developing a digestive tract tumour. Indeed, human colon carcinoma cells have lower ceramide content compared to normal colon mucosa cells (Selzner et al., 2001). Dietary SLs seem to have a role in CRC prevention, since studies with experimental animal have shown that consumption of SLs inhibits colon carcinogenesis. The possible mechanism involved in SLs suppression of colon carcinogenesis is that exogenously supplied SLs bypass a sphingolipids signalling defect important in cancer, for example, a loss of cellular sphingomyelin turnover to produce ceramide and sphingosine (Berra et
al., 2002).
Ceramide has been implicated in CRC apoptosis in several models. For instance, C2
-ceramide as well as sphingoid bases sphingosine and sphinganine, inhibit proliferation and induced apoptotic cell death in HT-29 and HCT-116 human CRC cell lines (Ahn and Schroeder, 2002). Cannabinoid receptor activation in colon cancer cell lines also seems to induce apoptosis in those cells, mediated by de novo synthesis of ceramide (Cianchi et al., 2008). Another study reports the use of curcumin, known for its anti-carcinogenic properties, to induce apoptosis in colon cancer cell lines (Moussavi et al., 2006). These authors demonstrated that curcumin apoptosis-inducing effect was accompanied by ceramide generation through de novo synthesis. Ceramide analogues had also been tested in vitro and in vivo in the induction of apoptotic cell death and prevention of tumour growth in metastic human colon cancer (Selzner et al., 2001). A ceramidase enzyme inhibitor B13, inhibits ceramide metabolism and leads to the accumulation of cellular ceramide, and was proved to be not toxic to normal cells in
26
vitro and to animals treated with B13 in vivo. This ceramidase inhibitor has been proved
to prevent the development of tumour metastases in vivo (Selzner et al., 2001).
The level of APC protein can also be associated with ceramide-induced apoptosis of colon cancer cells. APC mutations, one of the most frequent mutations occurring in CRCs, generate a truncated APC protein. Therapeutic strategies to induce apoptosis through processes that involve reduced dosage effect levels of APC were studied and treatment of colon cancer cell lines expressing wild-type APC with C2-ceramide
revealed a reduction of the APC protein levels. This effect is associated with increased apoptosis, that is also observed when colon cancer cell lines expressing the truncated from of APC protein are treated with C2-ceramide (Jaiswal and Narayan, 2004).
In summary, all the evidences show that ceramide has a role in inducing apoptosis in CRC.