Genetic diversity in Mexican Creole pigs with candidate genes associated with productive characters
Texto
Imagem
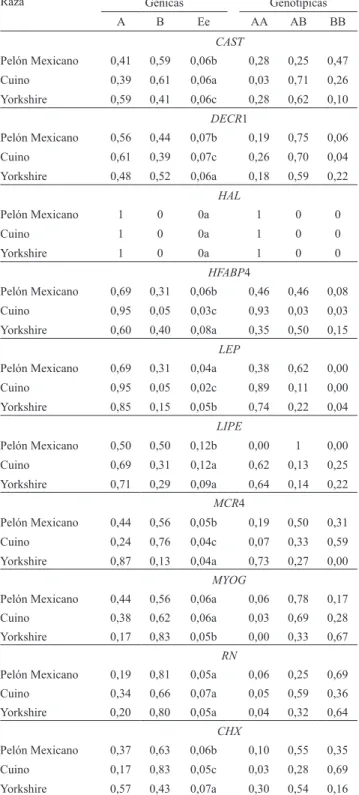
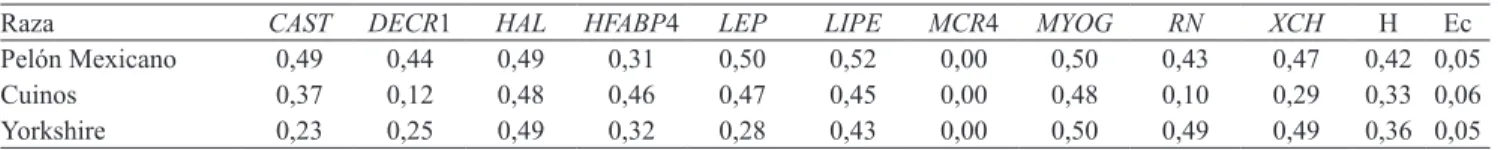
Documentos relacionados
mayor frecuencia en los pacientes fueron: medición. de señales vitales, evaluación de
En general, las diferencias numéricas presentadas entre los valores registrados y los establecidos en promedio para la nomenclatura en sistemas tradicionales,
EN ZONAS INUNDABLES Y NO INUNDABLES Los adultos fueron observados durante todo el año, encontrándose con mayor frecuencia entre los meses de enero a marzo en restinga
Así, pues, cuando los representantes de las Américas se reunieron en la Ciudad de México en diciembre de 1907 para celebrar la Tercera Convención Sa- nitaria Internacional,
Los análisis de diferencias de medias, como se observa en la Tabla 2, manifiestan diferencias significa- tivas en todas las estrategias de aprendizaje en función de los
En el análisis de la frecuencia de los marcadores de infec- ción según el sexo, se hallaron diferencias estadísticas en las pruebas del banco de sangre y del centro de referencia
Sin embargo, la presencia de pocos alelos en el lote (2 y 3 alelos por locus), los alelos de baja frecuencia, observados en los parentales y en las larvas, y el alelo raro,
es de 1945 que exigió del escritor una actitud. El manifiesto que resulta del Congreso de la ABDE se obtuvo como un consenso final en. torno al fin de la dictadura Vargas. El
