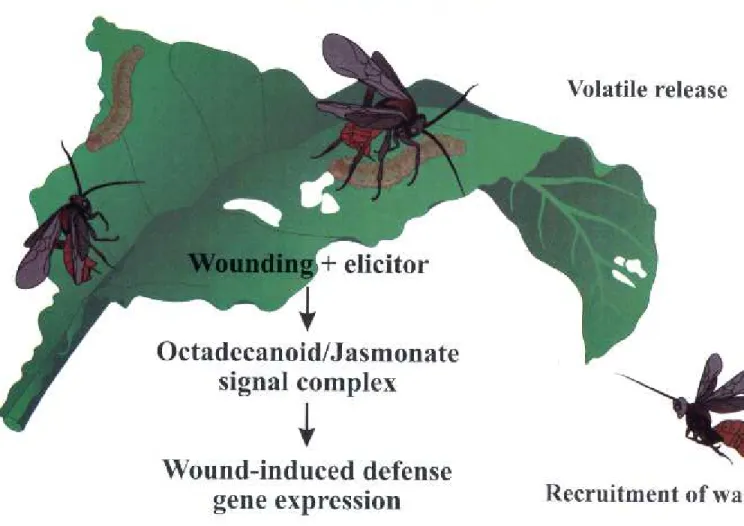
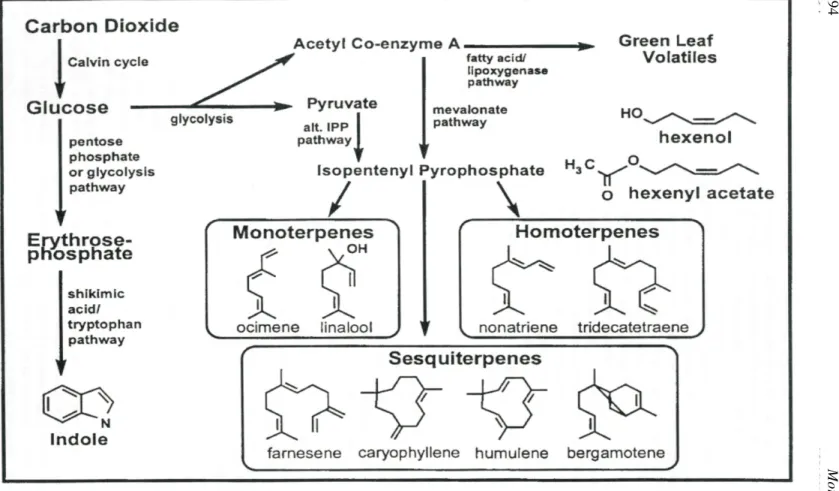
FORUM Examining Plant-Parasitoid Interactions in Tritrophic Systems
Texto
Imagem
Documentos relacionados
De seguida, a abordagem a cada fator e a sua influência ou não na web experience, e, por fim, a conclusão se os clientes concordam que a web experience influenciou ou
Auditory experimental stimuli derived from two sources: (1) environmental auditory stimuli generated by the software that managed the operant procedure, presented through
Size variation between males and females of aphidiine parasitoids are usually more conspicuous when these parasitoids are reared in smaller hosts and tends to decrease as the
The synthetic sex pheromone component methyl 2,6,10-trimethyltridecanoate is used as a kairomone during the searching behavior of Telenomus podisi ; this compound is preferred to
To elucidate the role of Os MYB1 in plant responses to environmental stimuli, we analyzed gene expression profile in rice under various abiotic stresses using
Para facilitar a comparação entre o desempenho mecânico à compressão simples entre os provetes de alvenaria normalizados e os provetes de alvenaria constituídos por 2 blocos,
Conclusions: Discrimination training with multimodal stimuli consisting of visual and odour cues leads to stimulus specific changes in the in vivo responses of the mushroom body of
In particular, semantically primed stimuli elicit an attenuated N400 response relative to unprimed stimuli (for review ref. In picture naming with distractor words, the amplitude of
