2018
UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
Analysis of neutrophil recruitment during tissue regeneration
in zebrafish models of human disease
Mariana Gomes Ferreira
Mestrado em Biologia Humana e Ambiente
Dissertação orientada por:
Professor Doutor Ângelo Calado
Professora Doutora Deodália Dias
i
Agradecimentos
Gostaria de começar por agradecer à professora Carlota Saldanha por me receber no seu laboratório e grupo no iMM-JLA para poder realizar a minha tese de mestrado. Ao Professor Ângelo Calado pela paciente e excelente orientação durante este projeto, especialmente quando as coisas davam para o torto e eu ficava perdida ele conseguia sempre ajudar-me a voltar ao rumo certo. À Doutora Patrícia Napoleão por estar sempre disponível para me ajudar e pela ideia do projeto secundário da minha tese em que consegui obter resultados. E também à Doutora Ana, o último membro do laboratório da Professora Carlota, pelo apoio ao longo da tese.
Esta tese não teria chegado tão longe sem o apoio e fé incondicionais das meninas da Fish Facility, a Lara e a Aida. Sempre que tinha um mau resultado ou um protocolo teimoso que não queria funcionar vocês apoiaram-me e lutaram comigo até à última, e quando tive que trocar de projeto continuaram a fazê-lo. Deram-me um canto onde escrever a tese, um ouvido amigo para reclamar com protocolos e papers, e muitas muitas gargalhadas sempre que precisei. Sem vocês esta tese seria completamente diferente! E claro, até as raparigas mais recentes da Fish Facility, a Filipa e a Carolina, merecem um obrigado por me aturarem durante esta fase final da escrita da tese.
Não podia faltar o agradecimento à excelentíssima equipa do bioimaging do iMM, a Ana e o António! Correram comigo a maior parte dos microscópios do instituto e deram-me formação em quase todos, sempre que tive um problema ou uma dúvida por mais descabida que fosse disponibilizaram-se a ajudar. E o mais importante de tudo deixaram-me seguir-vos para aprender mais sobre bioimaging mesmo quando já não era relacionado com a minha tese, alimentando a minha paixão pelo bioimaging. Poder distrair-me convosco e com o bioimaging ajudou a passar aqueles intermináveis tempos de incubação ou de aquisição de imagem que eram simultaneamente demasiado longos e demasiado curtos para trabalhar noutras coisas.
Tenho também que agradecer à Doutora Leonor Saúde e ao seu grupo do iMM por me acolherem e darem uma secretária onde pudesse escrever a tese, quando a professora Carlota se reformou e eu fiquei “sem casa” no instituto. E claro pelos convites para os convívios do grupo e da fish facility que foram sempre uma óptima maneira de arejar e distrair da pressão de escrever a tese. Com um especial obrigado a Guida, a lab mãenager do grupo, por ser a pessoa mais querida de sempre e ter sempre um bocadinho para me aturar.
Quero agradecer também à minha querida amiga Cristina Torcato que já me atura desde os tempos idos da licenciatura em biologia por continuar a aturar-me e ser sempre um ombro amigo, apesar da distância física!
Tenho que agradecer à Inês Antunes, a minha tecnologicamente inábil amiga, por também me aturar desde a licenciatura e aturar as minhas nerdisses que se estavam sempre a manifestar mesmo quando não percebia o que se estava a passar.
À Cláudia Cavacas, amiga de mestrado sempre disponível para almoçarmos no iMM e beber um cafezinho depois.
E claro à mais recente, mas não menos importante, amiga a Mariana Costa, que sempre me deixou segui-la e ajuda-la enquanto utilizava o lightsheet que eu nunca pude usar, também ela apoiando o meu amor pelo bioimaging. E claro por me aturar o tempo todo desde que chegou ao iMM.
Agradeço também ao Luís e ao Manuel, colegas e amigos que mesmo que se passem meses parece que ainda nos tínhamos vito no dia anterior. Ao João, ao David, ao Mauro, ao Tiago e à Cristina, novamente, por serem um óptimo grupo de nerd-friends para jogos de tabuleiro, cartas e Pathfinder.
ii
Como não pode faltar tenho que agradecer a toda a minha família pelo apoio incondicional e por acreditarem sempre nas minhas escolhas mesmo quando achavam que não eram as melhores. E obviamente aos meus pais por financiarem toda esta aventura que foi o meu mestrado.
Em último quero agradecer às minhas caturras, o Benjamim e a Ofélia, por serem os meus bebés e os mais fofinhos do mundo mesmo quando gritam o suficiente para me magoar os ouvidos!
Se me esqueci de agradecer a alguém não foi de propósito, e mesmo aos que aqui menciono posso não parecer agradecida no dia-a-dia, mas estou mais do que vocês imaginam!
OBRIGADA A TODOS! ESTA TESE NÃO TERIA ACONTECIDO SEM VOCÊS! ADORO-VOS!
iii
Abstract
Inflammation is an essential process of an organisms’ response to assault, preventing infection and promoting tissue repair. Neutrophils are the first leukocytes to be recruited, arriving in large numbers a few hours after injury. Having been considered both indispensable for proper resolution of inflammation but a hindrance to wound healing, their role requires further study.
Diabetes mellitus (DM) is a worldwide public health concern that is characterized by chronic elevated fasting blood glucose levels (hyperglycemia), usually caused by deficient production of insulin or developed resistance to insulin. Prolonged hyperglycemia has been associated with many disease complications like the diabetic foot, which is in part caused by impaired wound healing associated with chronic inflammation. DM models in zebrafish (Danio rerio), an animal model renowned for its regenerative capabilities, display impaired regeneration of the caudal fin (Olsen et al., 2010). Therefore, we aimed first to study neutrophil recruitment during impaired caudal fin regeneration in a DM zebrafish model, and address whether it differed from what occurred under normal conditions.
Due to animal welfare concerns and most importantly, to the high inconsistency of the glycemic outcome among the tested animals, we were unable to establish a reliable DM model in our lab. As such, neutrophil recruitment during zebrafish caudal fin regeneration was only characterized in healthy animals of the Tg(mpx:GFP) transgenic line through widefield fluorescence microscopy. Under normal conditions, we observed an early influx of neutrophils at 6 hours after amputation. Neutrophil recruitment was further reduced by 1 day after amputation, when the number of neutrophils at injured fin returned to basal levels. This recruitment pattern was consistent with the early role of neutrophils during inflammation. However, our data contrasted with the findings by Petrie et al. (2015) that only observed a peak of neutrophil recruitment at 3 days after amputation.
Diabetes mellitus has also been shown to be associated with cardiovascular diseases, such as myocardial infarction (MI). This cardiovascular pathology is a major source of mortality and disability, due to the inability of humans to regenerate the damaged heart tissues forming non-contractile scar tissue instead.
Importantly and like other pathological conditions, myocardial infarction is also associated with an inflammatory process. Little is still known about the enrollment and the role of neutrophils after MI. In mouse models of MI, neutrophils have been shown to be recruited to the infarcted area (Ma et al., 2016). As no study so far has addressed the recruitment of neutrophils in zebrafish MI models, the second aim in this work was therefore to investigate this issue in adult animals by analyzing immunolabeled heart cryosections with widefield fluorescence microscopy. From our results, a marked neutrophil recruitment to the infarcted area was observed at an early time-point of 6 hours after injury and was further maintained at high levels up to 1 day after injury. By 3 days the quantity of neutrophils at the injury seemed to have been reduced and this reduction was even more striking by 7 days after injury. As observed in inflammatory processes associated to other biological contexts, a higher influx of neutrophils to the injury was thus evidenced at early time-points after MI in adult zebrafish. Overall, these results have provided a first insight on the enrollment of neutrophils in the inflammatory and regenerative processes following MI in zebrafish, and thus contributed to a better understanding of the role of these leukocytes in these processes.
Altogether, the neutrophil recruitment patterns observed after amputation of the caudal fin as well as after myocardial infarction are consistent with the early role of neutrophils during the inflammatory phases of both zebrafish regenerative processes. This leads us to suggest that despite all controversies neutrophils should, at least in zebrafish, play a role in tissue regeneration.
v
Sumário
A resposta inflamatória é um processo essencial do organismo na sua defesa face a agressões, protegendo contra infeções e estimulando o processo de reparação dos tecidos. Quando as células imunitárias residentes nos tecidos detetam agressões ou agentes patogénicos libertam sinais que desencadeiam a resposta inflamatória, começando por aumentar o fluxo sanguíneo para a área afetada. Os neutrófilos são o primeiro tipo de leucócitos a ser recrutados, começando a acumular-se na área cerca de 6 horas após o dano. Eles fagocitam agentes patogénicos e células necróticas, libertando também substâncias antimicrobianas para o meio extracelular. Este influxo inicial de neutrófilos é posteriormente seguido pelo recrutamento de monócitos em circulação, que no foco de inflamação se diferenciam em macrófagos que apresentam maior capacidade fagocitária que os neutrófilos. A resolução da inflamação envolve o início da apoptose dos neutrófilos e sua consequente eliminação mediada por fagocitose pelos macrófagos. Estes processos contribuem para uma mudança fenotípica nos macrófagos que lhes confere uma função anti-inflamatória e de pro-resolução que estimula a reparação tecidual e a recuperação da homeostasia do organismo.
A reparação dos tecidos pode ter três resultados: reparação com cicatrização mínima, formação de cicatriz, ou regeneração dos tecidos. Todos começam com uma resposta inflamatória e formação de um tecido de granulação, rico em fibroblastos e capilares sanguíneos. Na formação de cicatriz o tecido fibrótico provisório é substituído por uma matriz permanente de colagénio, conduzindo à perda de função, total ou parcial, do tecido. Para ocorrer regeneração, é necessária a formação de um blastema, uma massa proliferativa de células indiferenciadas capaz de se re-diferenciar quando chegam ao seu local de destino.
O papel da resposta inflamatória na reparação e regeneração de tecidos ainda não está totalmente esclarecido, existindo dados contraditórios. Por um lado, a depleção de neutrófilos em ratinho leva a uma reparação mais rápida dos tecidos (Dovi, et al., 2003); por outro lado, na salamandra a resposta inflamatória é indispensável para regenerar a lente ocular após dano (Kanao and Miyachi, 2006).
O peixe-zebra (Danio rerio) constitui um modelo animal único pois permite o estudo da resposta inflamatória durante a regeneração de tecidos. O seu sistema imunitário é equivalente ao dos mamíferos, e este peixe tem capacidade de regenerar quase todas as partes do seu corpo como barbatanas, o coração e a espinal-medula. O peixe-zebra é também um modelo animal para o qual um grande número de linhas transgénicas têm vindo a ser criadas ao longo dos últimos anos nas quais diversos tipos celulares são especificamente marcados com proteínas fluorescentes, como por exemplo neutrófilos, macrófagos e cardiomiócitos.
A diabetes mellitus é uma doença que afetava cerca de 366 milhões de adultos mundialmente em 2011. É caracterizada por uma elevação crónica dos níveis de glicose no sangue (hiperglicemia) em jejum. Pode ser causada quer por produção insuficiente de insulina (Tipo 1) quer por desenvolvimento de resistência à insulina (Tipo 2). Independentemente da causa, a hiperglicemia prolongada pode levar a danos a longo prazo mesmo após reestabelecer níveis normais de glicémia. Uma das várias complicações patológicas secundárias é por exemplo o pé diabético, para a qual contribuem uma deficiente capacidade de reparação e cicatrização tecidual e a inflamação crónica evidenciada em indivíduos diabéticos. Da mesma forma já foi demonstrado que modelos de diabetes em peixe-zebra apresentam um efeito secundário semelhante, evidenciando por exemplo problemas na regeneração da barbatana caudal (Olsen et al., 2010). Com o objetivo de analisar o recrutamento de neutrófilos durante a regeneração comprometida da barbatana caudal num modelo de diabetes de peixe-zebra, seria primeiro necessário estabelecer o modelo diabético na Zebrafish facility no iMM-JLA. Dois protocolos diferentes foram testados para este fim: injeção da droga diabetogénica estreptozocina, que especificamente destrói as células produtoras de insulina; e imersão dos peixes numa solução de glicose, para indução de hiperglicemia por absorção de glicose do meio. Mas nenhum dos protocolos apresentou resultados
vi
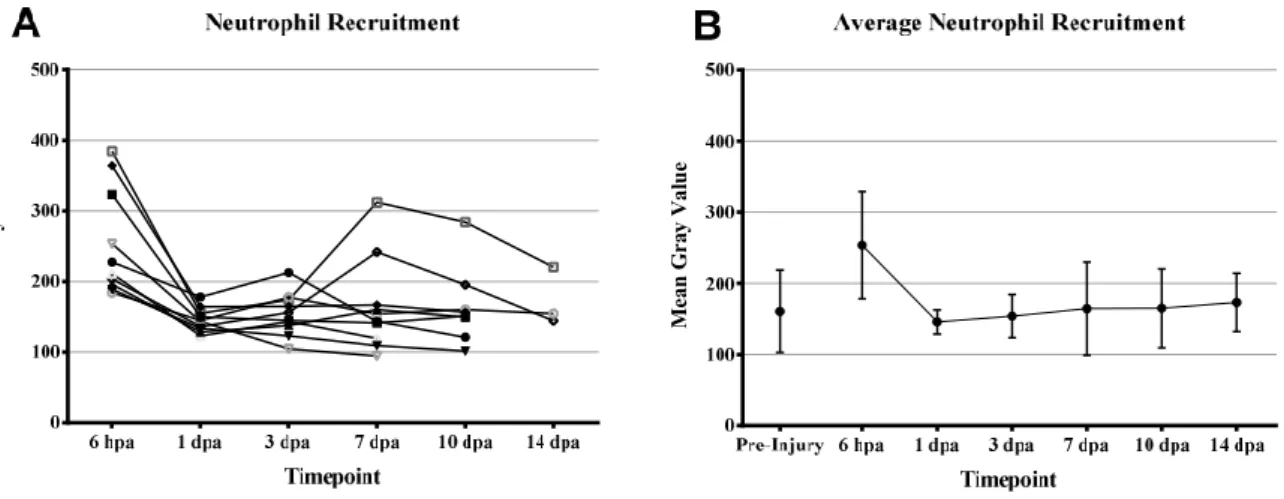
suficientemente consistentes para poderem ser considerados fiáveis na indução de hiperglicemia nos nossos peixes-zebra. Para além disso, muitos dos animais injetados com estreptozocina apresentaram efeitos secundários graves. Assim, o recrutamento de neutrófilos durante a regeneração da barbatana caudal foi apenas analisado em animais saudáveis pertencentes à linha transgénica Tg(mpx:GFP) por microscopia de fluorescência de campo largo. Os peixes desta linha apresentam neutrófilos que expressam especificamente a proteína GFP (Proteína Fluorescente Verde) e permitindo a sua observação específica com recurso a microscopia de fluorescência. Uma análise semelhante havia sido previamente publicada por Petrie et al. (2014), em que se observou um crescente recrutamento de neutrófilos que atingia o seu pico aos 3 dias após a amputação. É de salientar que os nossos resultados diferem destes, uma vez que observámos um influxo inicial de neutrófilos para a ferida às 6 horas após a amputação, com a densidade de neutrófilos na ferida e na zona regenerada a voltar a níveis fisiologicamente normais ao fim de 1 dia após a amputação.
A diabetes está também intimamente associada ao desenvolvimento de doenças cardiovasculares, como os enfartes do miocárdio. Esta patologia cardíaca em particular é uma das maiores causas de mortalidade e incapacidade no mundo, face à incapacidade dos humanos em regenerar o tecido necrótico formando antes uma cicatriz não contráctil no tecido cardíaco. Como qualquer outro estímulo danoso ao organismo o enfarte desencadeia uma resposta inflamatória. Esta é seguida pela formação de um tecido granuloso fibrótico que nos mamíferos é posteriormente substituída por uma cicatriz rica em colagénio, e no peixe-zebra dá lugar a novo tecido cardíaco. Em relação à resposta inflamatória associada, modelos de enfarte do miocárdio em ratinho permitiram anteriormente observar um significativo recrutamento de neutrófilos para a zona de enfarte, leucócitos que habitualmente estão ausentes do tecido cardíaco (Ma et al., 2016). Neste estudo observou-se uma concentração máxima de neutrófilos ao final de um dia após lesão, que embora diminuindo a partir desta altura, se manteve em níveis ainda significativamente altos aos 7 dias após a lesão. Para além deste estudo a literatura publicada sobre o recrutamento de neutrófilos após enfarte do miocárdio é impressionantemente escassa. Como tal, o nosso segundo objetivo foi analisar o recrutamento dos neutrófilos durante a resposta inflamatória induzida pelo enfarte do miocárdio em peixe-zebra.
Neste trabalho, o enfarte foi induzido na linha transgénica Tg(mpx:GFP) por crio-lesão após exposição do ventrículo. Para se determinar se a incisão efetuada para a exposição do ventrículo influenciava o recrutamento de neutrófilos foram utilizados peixes controlo da ferida, nos quais apenas se procedeu à exposição do ventrículo. Para aquisição dos dados de recrutamento recorreu-se ao tratamento histológico dos corações, os quais foram incluídos em Optimal cutting medium (OCT), congelados, cortados a frio e montados em lâminas de vidro. Os cortes obtidos foram posteriormente submetidos a diferentes procedimentos. Parte destes foram corados com hematoxilina e eosina para visualização da estrutura tecidual por microscopia de campo claro. Com os restantes a serem submetidos a marcação por imunohistoquímica com anticorpos específicos para a deteção do GFP presente nos neutrófilos e outros componentes cardíacos sendo as imagens dos mesmos adquiridas por microscopia de fluorescência de campo-largo para posterior análise. Um script de Matlab foi posteriormente utilizado para analisar o recrutamento de neutrófilos observado nas imagens de fluorescência adquiridas. Globalmente, a análise das imagens adquiridas revelou um recrutamento de neutrófilos mais pronunciado às 6 horas e 1 dia após o enfarte, que diminuiu significativamente aos 3 e 7 dias após lesão. Contrariamente ao observado em ratinho, observámos um número substancialmente reduzido de neutrófilos presentes na área afetada nos dias mais tardios. A reduzida presença de neutrófilos nos corações dos peixes de controlo da ferida permitiu ainda determinar que o recrutamento observado nos corações submetidos à crio-lesão era devido este dano e não à lesão efetuada para expor do ventrículo. No geral, os resultados preliminares obtidos neste trabalho permitiram confirmar o recrutamento inicial de neutrófilos durante a regeneração do coração após enfarte do miocárdio em peixe-zebra.
vii
Em suma no presente trabalho foi possível concluir que no peixe-zebra, tanto na resposta regenerativa à amputação da barbatana caudal como no enfarte do miocárdio, se observaram padrões de recrutamento de neutrófilos para as lesões consistentes com a função destes leucócitos no início dos processos inflamatórios associados. Estes resultados levam-nos a sugerir que pese embora a controvérsia que envolve a sua atuação na regeneração os neutrófilos devam, pelo menos no peixe-zebra, desempenhar uma função relevante neste processo fisiológico.
viii
Table of Contents
Agradecimentos i
Abstract iii
Sumário v
List of Figures and Tables x
List of abbreviations and acronyms xii
Chapter 1 - Introduction 1
1.1 - Inflammation 1
1.1.1 - Neutrophils and Macrophages 1
1.1.2 - Inflammatory Response 2
1.1.3 – Leukocyte recruitment 3
1.2 - Wound healing and Regeneration 4
1.2.1 - Is Inflammation necessary for Regeneration? 4
1.2.2 - Zebrafish, an animal model for Regeneration and Inflammation 5
1.3 - Diabetes Mellitus 6
1.3.1 - Zebrafish models of Diabetes Mellitus 8
1.4 - Myocardial Infarction 9
1.4.1 - Zebrafish as a model of cardiac regeneration 9
1.4.2 - Neutrophil recruitment in Animal models of Myocardial Infarction 11
1.5 – Objectives 11
Chapter 2 – Materials and Methods 12
2.1 - Animal Model 12
2.1.1 - Zebrafish Husbandry 12
2.1.2 - Transgenic Lines 12
2.1.3 - Anesthetics & Recovery 12
2.2 - Models and procedures for studies on Diabetes Mellitus 13
2.2.1 - Streptozotocin-induced Diabetes Mellitus model 13
2.2.2 - Glucose immersion induced Diabetes Mellitus model 14
2.2.3 - Blood collection and determination of fasting blood glucose levels 14 2.2.4 - Caudal Fin Regeneration and Imaging with Inverted Widefield Microscopy 15 2.3 - Model and procedures for studies of Myocardial Infarction Regeneration 15
2.3.1 - Cryoinjury induced Myocardial Infarction 15
2.3.2 - Heart Collection 15
2.3.3 - Heart preparation and Imaging with Lightsheet Fluorescence Microscopy 16
2.3.4 - Heart Cryoembedding 16
2.3.5 - Heart Cryosectioning 16
ix
2.3.7 - Immunohistochemistry and Imaging with Widefield Fluorescence Microscopy 16
2.4 - Neutrophil Recruitment Data Processing and Analysis 17
2.4.1 - Caudal Fin Regeneration Data Processing and Analysis 17
2.4.2 - Myocardial Infarction Data Processing and Analysis 17
Chapter 3 - Results 19
3.1 - Diabetes Models 19
3.1.1 - Streptozotocin (STZ) Intraperitoneal Injection 19
3.1.2 - Glucose Immersion 21
3.1.3 - Neutrophil Recruitment in Caudal Fin Regeneration 23
3.2 - Myocardial Infarction 26
3.2.1 - Zebrafish Heart LSFM Imaging 26
3.2.2 - Analysis of Neutrophil recruitment after MI by immunohistochemistry on
cryosections 27
3.2.3 – Protocol Alterations 37
Chapter 4 - Discussion 39
4.1 - Analysis of leukocyte recruitment in wound healing in Diabetes mellitus 39
4.1.1 - Diabetes Mellitus Models 39
4.1.2 - Caudal Fin Regeneration 40
4.2 - Myocardial Infarction 42
4.2.1 - Zebrafish Heart LSFM Imaging 42
4.2.2 - Analysis of Neutrophil recruitment after MI by IHC on cryosections 43
4.2.3 - Final considerations on the Zebrafish MI model 45
4.3 – Conclusions 46
References 47
x
List of Figures and Tables
Figure 1.1 – Representative cartoon of a macrophage (left) and a neutrophil (right). Figure 1.2 – Mechanisms of leukocyte recruitment from the blood vessels.
Figure 1.3 – Hyperglycemic fish have a reduced rate of caudal fin regeneration. Figure 1.4 – Representative schematics about the zebrafish heart anatomy and injuries.
Figure 2.1 – Schematic representation of the anatomic landmarks for blood collection from the adult zebrafish.
Figure 2.2 - Graphic representation of the division of the radial profile executed on each heart slice. Figure 3.1 – Zebrafish injected with streptozotocin showing signs of morbidity.
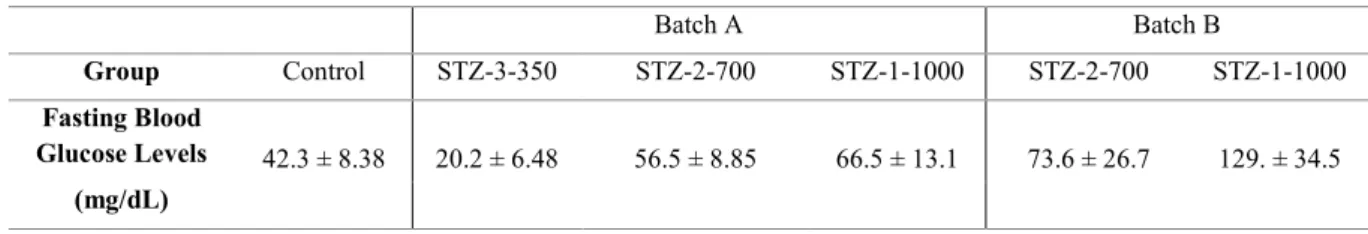
Figure 3.2 – Fasting blood glucose levels for (a) STZ batch A and (b) STZ batch B at 4 days post last injection.
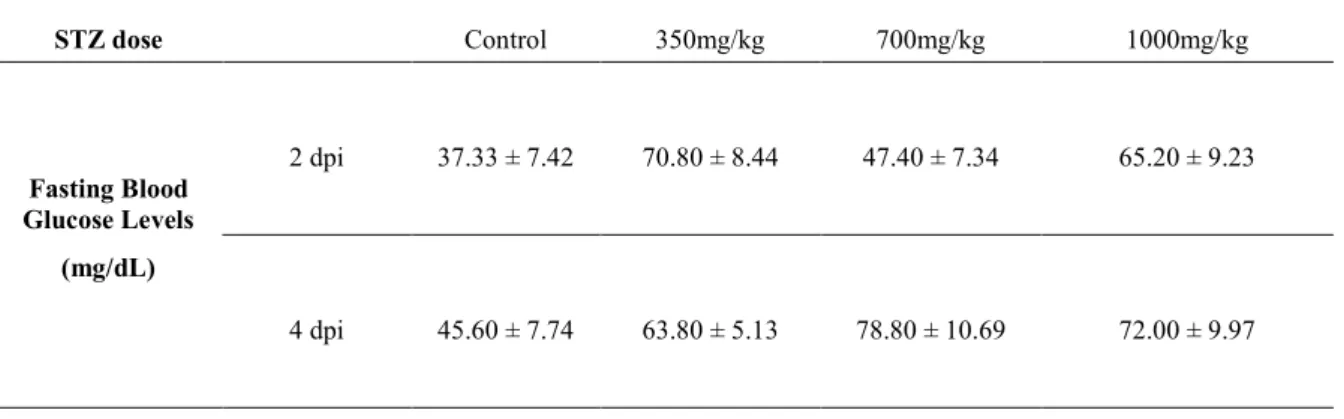
Figure 3.3 – Fasting blood glucose levels for a single injection of various STZ doses at 2 and 4 days post injection (dpi).
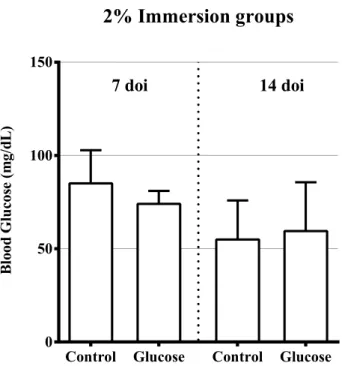
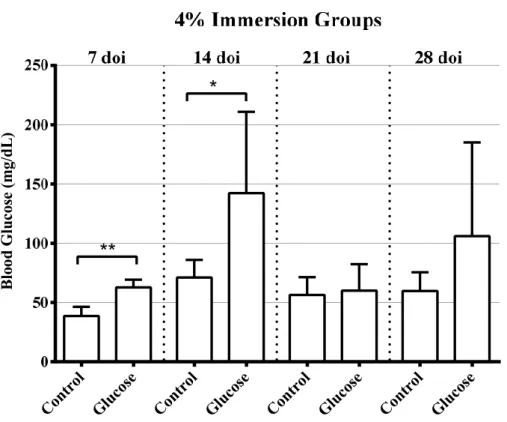
Figure 3.4 – Fasting blood glucose levels after 7 and 14 days of immersion (doi) in 2% Glucose. Figure 3.5 – Fasting Blood Glucose levels after 7, 14, 21, and 28 days of immersion (doi) in 4% Glucose. Figure 3.6 – (A) Mean Gray Value of the regenerated area for each caudal fin per time-point after amputation. (B) Average Mean Gray Value of the regenerated area in injured fish per time-point and of the similarly simulated area in uninjured controls.
Figure 3.7 – (A) Analyzed area size in µm2 for each individual caudal fin for each time-point. (B) Mean
analyzed caudal fin area in µm2 for each time-point.
Figure 3.8 – Time-lapse of a Tg(mpx:GFP) zebrafish caudal fin regeneration in transmitted light (A – F) and fluorescent imaging of the fluorescent neutrophils (A1-F1).
Figure 3.9 – Comparison between fluorescent imaging of GFP labeled cells during zebrafish caudal fin regeneration of Tg(mpeg1:EGFP) macrophages at 4 days after amputation (dpa) (A) and Tg(mpx:GFP) neutrophils at 3 dpa (B).
Figure 3.10 – Representative slices from the lightsheet fluorescence microscopy z-stack of a Tg(mpx:GFP) cleared zebrafish heart.
Figure 3.11 – Representative slices of control hearts.
Figure 3.12 – Representative slices at 6 hours post injury (hpi) of hearts submitted to myocardial infarction (MI) and SHAM procedure.
Figure 3.13 – Representative slices at 1-day post injury (dpi) of hearts submitted to myocardial infarction (MI) and SHAM procedure.
Figure 3.14 – Representative slices at 3-days post injury (dpi) of hearts submitted to myocardial infarction (MI) and SHAM procedure.
Figure 3.15 – Representative slices at 7 days post injury (dpi) of hearts submitted to myocardial infarction (MI).
Figure 3.16 – Distribution of GFP fluorescence intensity over the four defined regions of the heart slices for each heart, and their average, for Control hearts.
xi
Figure 3.17 - Distribution of GFP fluorescence intensity over the four defined regions of the heart slices for each heart, and their average, at 6-hour post injury for myocardial infarction (A) and SHAM procedure (B).
Figure 3.18 – Distribution of GFP fluorescence intensity over the four defined regions of the heart slices for each heart, and their average, at 1-day post injury for myocardial infarction (A) and SHAM procedure (B).
Figure 3.19 – Distribution of GFP fluorescence intensity over the four defined regions of the heart slices for each heart, and their average, at 3-days post injury for myocardial infarction (A) and SHAM procedure (B).
Figure 3.20 – Distribution of GFP fluorescence intensity over the four defined regions of the heart slices for each heart, and their average, at 7-days post injury.
Figure 3.21 – Average percentage of GFP fluorescence intensity in the heart region closest to the infarcted area in each time-point.
Figure 3.22 – Representative slices of hearts at 1-day post injury with myocardial infarction fixed before or after cryoembedding.
Figure 3.23 – Detail from representative slices of hearts at 1-day post injury with myocardial infarction fixed before or after cryoembedding.
Figure S.1 – Distribution of GFP fluorescence intensity over the four defined regions of the heart slices for each heart, and their average, for Control hearts.
Figure S.2 - Distribution of GFP fluorescence intensity over the four defined regions of the heart slices for each heart, and their average, at 6-hour post injury for myocardial infarction (A) and SHAM procedure (B).
Figure S.3 - Distribution of GFP fluorescence intensity over the four defined regions of the heart slices for each heart, and their average, at 1-day post injury for myocardial infarction (A) and SHAM procedure (B).
Figure S.4 - Distribution of GFP fluorescence intensity over the four defined regions of the heart slices for each heart, and their average, at 3-days post injury for myocardial infarction (A) and SHAM procedure (B).
Figure S.5 – Distribution of GFP fluorescence intensity over the four defined regions of the heart slices for each heart, and their average, at 7-days post injury.
Table 3.1 – Fasting blood glucose levels for STZ batches A and B at 4 days post-last injection (dpli). Table 3.2 – Fasting blood glucose levels for a single injection of various STZ doses at 2 and 4 days post
injection (dpi).
Table 3.3 – Fasting blood glucose levels after 7 and 14 days of immersion (doi) in 2% Glucose. Table 3.4 – Fasting Blood Glucose levels after 7, 14, 21, and 28 days of immersion (doi) in 4% Glucose. Table 3.5 - Comparison of Mean Gray Value of 6 hours post amputation (hpa) with other time-points (pre-injury, 1, 3, 7, 10, and 14 days post amputation (dpa)) using a post hoc multiple test of a one-way ANOVA.
xii
List of abbreviations and acronyms
BSA – Bovine serum albumin
DAMPs – Damage-associated molecular patterns DGAV – Direcção-Geral de Alimentação e Veterinária DM – Diabetes mellitus
DNA – Deoxyribonucleic Acid
EGFP – Enhanced Green Fluorescent Protein FBGL – Fasting blood glucose levels
GFP – Green Fluorescent Protein HE – Hematoxylin and Eosin IHC - Immunohistochemistry
iMM-JLA – Instituto de Medicina Molecular - João Lobo Antunes LSFM – Lightsheet Fluorescence Microscopy
MI – Myocardial infarction MM – Metabolic memory
mpeg1 – Macrophage expressed gene 1 mpx – Myeloperoxidase gene
OCT – Optimal Cutting Temperature
PAMPs – Pathogen-associated molecular patterns PBS – Phosphate Buffered Saline
PRRs – Pattern-recognition receptors ROI – Region of interest
ROS – Reactive oxygen species STZ – Streptozotocin
1
Chapter 1 - Introduction
1.1 - Inflammation
The inflammatory response, or inflammation, is the protective response of the innate immune system to assault, be it either damaged tissues or infectious agents, with the objective of restoring tissue homeostasis. The process involves tissue resident cells, blood vessels, proteins and other mediators, and immune cells, like neutrophils and macrophages. All these factors work together to eliminate the cause of the damage, clear any resulting dead cells and tissues, and initiate tissue reparation.
1.1.1 - Neutrophils and Macrophages
Understanding the functions and behaviors of the two main cellular types involved in inflammation, neutrophils and macrophages, is key to achieve further comprehension about the inflammatory response.
Neutrophils belong to a group of polymorphonuclear leukocytes that differentiate from the myeloid precursors by granulopoiesis in the bone marrow, being easily identified by their lobed nucleus (Figure 1.1). Neutrophils can also be distinguished from other leukocytes by the presence of myeloperoxidase in their lysosomes, an enzyme that is a powerful oxidant and antimicrobial agent. (Kumar et al., 2013) In order to keep their production balanced only a small percentage is released into circulation, where they have a short half-life due to constitutive apoptosis (Cascão et al., 2009). Neutrophils are classified as short-lived cells due to their short circulating half-life that varies from around 1.5 hours in mice to 8 hours in humans, although a more recent study has increased the estimation of neutrophil half-life in mice to 12.5 hours and 5.4 days in humans. The first methodology involved ex vivo manipulation to label the neutrophils before being evaluated in vivo, the second used deuterium-labeled water that most likely deuterium-labeled bone marrow neutrophils over estimating their half-life. (Kolaczkowska and Kubes, 2013) Neutrophils are the most abundant leukocytes in the blood and are divided into two main groups, a circulating pool mostly in large blood vessels and a marginal pool, where neutrophils accumulate transiently in organ capillaries, in the lungs and liver for example. (Cascão et al., 2009) However, when activated the neutrophil’s lifespan can increase several fold while their release and production in the bone marrow is increased. This allows them to take part in complex activities like the inflammatory response where they can eliminate pathogens by phagocytosis, release antibacterial proteins to the extracellular environment through degranulation, or through neutrophil extracellular traps made from released DNA histones and enzymes (Kolaczkowska and Kubes, 2013).
Macrophages are mononuclear leukocytes whose functions involve phagocytosis and removal of cellular debris from apoptotic cells or tissue necrosis, as well as having a role in tissue remodeling (Mosser and Edwards, 2008). They are characterized by large vacuoles often filled with cellular debris (Figure 1.1). Blood monocytes are the circulating mononuclear leukocytes. In case of inflammation these migrate into the affected area where they are able to differentiate into tissue macrophages, supplementing the resident macrophages during the inflammatory response. Like with neutrophils their production and release from the bone marrow is increased in these situations (Keightley et al., 2014).
Macrophages are usually divided into two categories, the pro-inflammatory M1 and the anti-inflammatory or tissue-repairing M2. This is an over simplistic division of macrophage phenotypes as they exist in a spectrum with the M1 and M2 states being in opposing ends (Forbes and Rosenthal, 2014). Type M1 macrophages are activated by microbial products and foreign substances, they produce enzymes to destroy phagocyted organisms and release pro-inflammatory signals. Type M2 macrophages are activated directly or by differentiation of M1 macrophages and are not actively microbicidal. Instead
2
they promote angiogenesis, activate fibroblasts and promote collagen synthesis which are indispensable for tissue repair (Kumar et al., 2013). It is also important to note that the switch from M1 to M2 macrophages is thought to be associated the phagocytosis of neutrophils during resolution of the inflammatory response (Keightley et al., 2014).
1.1.2 - Inflammatory Response
Whenever there is an assault to the organism an inflammatory response is triggered. It is present in every process from a limb amputation to hitting your hand on a desk, with an important role in the control of infection. The first visual signs of inflammation in humans were described more than 2000 years ago. Currently they are believed to be heat, redness, swelling, pain, and loss of function in the affected area (Kumar et al., 2013).
Pattern-recognition receptors (PRRs) are able to identify small molecular “danger sequences” and trigger the inflammatory response. The danger sequences can be pathogen-associated molecular patterns (PAMPs) like viral RNA or bacterial toxins from invading pathogens, or danger-associated molecular patterns (DAMPs) from endogenous danger or stress signals from tissue damage (Eming et al., 2009). The PRRs are expressed by several immune cells, like tissue resident macrophages, which when activated by PAMPs or DAMPs can secrete a plethora of inflammatory cytokines and/or reactive oxygen species (ROS). The first detectable physical reaction of this process is increased blood flow to the affected area, with vasodilation and increased vascular permeability that allow the passage of fluids and later immune cells into the tissues.
At 6 hours after stimuli detection neutrophils are the main type of leukocyte present at the inflammation site due to their abundance in the blood and being faster than monocytes at infiltrating tissues. There neutrophils phagocyte cell debris and invading microorganisms but can also release substances to extracellularly destroy the pathogens preventing infection. After a few hours, when monocytes start arriving, neutrophils begin undergoing apoptosis (Kumar et al., 2013). However, this dogma has been recently challenged as recent data suggests some neutrophils instead of suffering apoptosis just migrate away during resolution of inflammation (Kolaczkowska and Kubes, 2013).
Blood monocytes infiltrate the tissues and then begin differentiating into M1 tissue macrophages. These are much more efficient at phagocyting cellular debris and microorganisms than neutrophils, even phagocyting the latter when these begin their apoptotic processes preventing the release of their dangerous cellular content into the tissues. Phagocytosis of neutrophils is thought to be an important factor in the shift of macrophages into M2 type, which promote resolution of the inflammatory response and are associated with tissue repair. When inflammation resolves macrophages die or just leave through the lymphatic system. The normal acute inflammatory response is fast, lasting from a few minutes to a
Figure 1.1 - Representative cartoon of a macrophage (left) and a neutrophil (right). The macrophage is a large cell capable of forming pseudopods with many vacuoles filled with phagocyted cellular debris. The neutrophil’s main characteristic is its lobed nucleus and many lysosomes. Adapted from Martin and Leibovich (2005).
3
few days until resolution. Inflammation is a very controlled process, but if left unchecked the same recruited cells and molecules that are so beneficial when fighting infection and removing necrotic tissues can begin damaging the surrounding healthy tissue. This is when the acute response becomes chronic. Chronic inflammation can last for years, differencing from the acute by having mononuclear cells, like macrophages, instead of neutrophils as the main cellular players, tissue destruction from recruited cell byproducts and new blood vessel formation (Kumar et al., 2013).
1.1.3 – Leukocyte recruitment
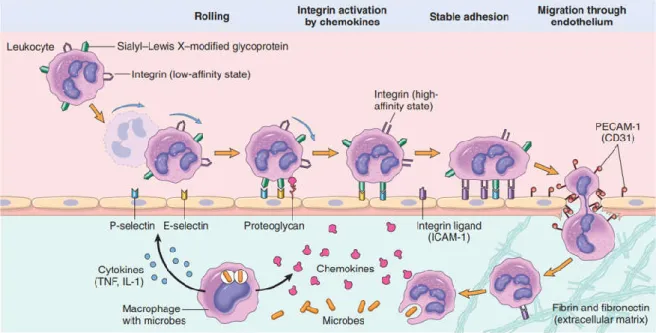
When the inflammatory response is triggered the freely circulating leukocytes have to be activated and recruited to the origin of the inflammatory signals. Leukocyte recruitment follow a sequence of several important steps: 1) Margination and rolling against the vessel walls; 2) Adhesion to the endothelium; 3) Transmigration between the endothelial cells; and 4) Migration along the interstitial tissues along a chemotactic gradient to its source (Figure 1.2).
Margination occurs naturally as the laminar blood flow pushes the bigger and heavier leukocytes to the walls of the vessels while the smaller erythrocytes can move faster in the center of the vessel. This gives leukocytes an opportunity to interact with the lining endothelial cells. As endothelial cells are locally activated by the inflammatory cytokines and other mediators, expression of adhesion molecules from the selectin family and integrin ligands is up-regulated. These adhesion molecules create loose connections with the ones present on the surface of leukocytes, mediating the light binding of leukocytes to the endothelium and their detachment from it, and in consequence, slowing the leukocytes down and making them tumble along the vessel wall in a process called rolling.
The endothelial cells also display chemoattractant cytokines (chemokines) secreted from cells at the inflammation site. These chemokines activate the rolling leukocytes which convert their low-affinity surface integrins into high-affinity integrins. These high-affinity integrins are then able to interact the integrin ligands expressed by the endothelial cells, leading to the stable adhesion of the leukocytes to the blood vessel wall.
Figure 1.2 – Mechanisms of leukocyte recruitment from the blood vessels. Here neutrophils are shown rolling, being activated, adhering to the endothelium, transmigrating through the endothelium and migrating towards the chemoattractants in the tissues. Taken from Kumar et al. (2013).
4
Transmigration of leukocytes is driven by chemokines and other chemoattractants that stimulate the passage of leukocytes between the endothelial cells at intercellular junctions, in a process called diapedesis. Once out of the blood the leukocytes follow a chemotactic gradient toward the site of infection or injury. Such gradient can be composed of bacterial products, chemokines, cytokines, or even ROS (Kumar et al., 2013).
1.2 - Wound healing and Regeneration
The inflammatory response is usually followed by wound healing, which can be divided into healing and scarring, or regeneration. Depending on the extension and nature of the stimuli that triggered it, there can be a few outcomes after resolution: 1) Healing, when the tissue is repaired with minimal scarring; 2) Scarring, if there is more extensive damage leaving extensive scar tissue; 3) Regeneration, the old damaged tissue is completely replaced by new fully functional tissue. As such all these outcomes begin very similarly, overlapping the resolution of inflammation, and after temporary tissue integrity is restored diverging and achieving very different results (Kumar et al., 2013).
When an injury occurs, the inflammatory response is triggered, and platelets are the first blood constituents to arrive at the site, forming clots that restore tissue homeostasis, provide a matrix to aid the recruitment of other circulating cells and release pro-inflammatory signals. Neutrophils are the first leukocytes to arrive, phagocyting pathogens and cells debris. Then blood monocytes invade the tissues differentiating into type M1 macrophages which take over phagocytosis of remaining pathogens and debridement of the wound, while neutrophils begin leaving the area or undergo apoptosis. Macrophages phagocyte the apoptotic neutrophils triggering their change into type M2. Endothelial cells, activated fibroblasts, and M2 macrophages form a provisional extracellular matrix called granulation tissue due to the large number of new capillary vessels resulting from the promotion of angiogenesis by the M2 macrophages. (Eming et al., 2009; Martin and Leibovich, 2005)
It is after the formation of the granulation tissue that differences between wound healing and regeneration arise. In wound healing this provisional matrix is replaced by a more permanent collagenous matrix, originating the scar tissue and often leading to loss of function in the affected area. In general, when regeneration occurs there has been a much smaller amount of granulation tissue formed. But the biggest difference in regeneration is the formation of a blastema, a mass of de-differentiated highly proliferative cells that are able to re-differentiate into mature tissues after migrating to their destination. As the damaged tissue is completely replaced by new equivalent tissue there is total recovery of function (Keightley et al., 2014).
1.2.1 - Is Inflammation necessary for Regeneration?
Regeneration differs from wound healing as there is no formation of permanent scar tissue or partial loss of organ function. Among vertebrates only amphibians and fish retain this ability as adults. There is no current consensus on how essential the inflammatory response is for regeneration. A few examples of either case will be briefly presented next.
Most mammalian models lose all their regenerative capabilities during post-natal life, except for full digit tip regeneration in adult mice and in young human children. Interestingly, the most prominent feature of fetal regeneration is the lack of inflammatory response, but while the pre-natal environment is mostly sterile an external wound is vulnerable to pathogens (Eming et al., 2009).
In sterile conditions, it has been shown that neutrophil depletion leads to much faster healing in mice when compared to their wild-type siblings (Dovi et al., 2003). Xenopus laevis larvae are capable of fully regenerating whole limbs, but as they begin undergoing metamorphosis their regenerative capability becomes increasingly impaired until the adult stage that cannot regenerate limbs. The causes
5
of this loss are mostly unknown, but it is thought to be related to the maturation of the immune system during metamorphosis, as the immune systems of larvae and adults are so different their tissues become histo-incompatible (Harty et al., 2003; Mescher and Neff, 2005). In an opposing position the regeneration of a damaged eye lens in a species of salamander requires an inflammatory response. After the damage occurs the inflammatory response is necessary to trigger the destruction of the damaged lens to allow for the regeneration of a new lens from the dorsal iris (Kanao and Miyachi, 2006).
1.2.2 - Zebrafish, an animal model for Regeneration and Inflammation
Zebrafish (Danio rerio) are fresh water fish native to the Indian sub-continent that began being used for the study of vertebrate embryogenesis in the 1930s. Transparent embryos, optically clear larvae, large clutch sizes and external fertilization allowed researchers to follow embryogenesis and development at the single cell stage, making the zebrafish uniquely suited for these studies. By the 1980s with the rise of genetic tools zebrafish mutants and transgenic constructs became increasingly common. In the last decade there has been an ever-increasing number of zebrafish mutants of genetic human diseases. For instance, acquired diseases like leukemia and melanoma can also be studied in zebrafish, which in conjunction with fluorescently labeled cell types allow for easier tracking of tumor onset, location and size. Transgenic zebrafish with labeled immune cell lineages have also allowed the analysis of immune cell behavior and recruitment during inflammation. Importantly, large clutch sizes can be used for high-throughput drug screenings, giving relevance to the model in the pharmaceutical field as well (Lieschke and Currie, 2007).
Zebrafish are capable of regenerating their kidney, liver, pancreas, jaw, spinal cord, heart and fins, with the caudal fin being one of the most studied and reliable examples of regeneration. Zebrafish fins are complex appendages. In particular, the caudal fin is composed of multiple lepidotrichia, bony fin rays that can bifurcate at the ends, each being composed by two concave hemirays joined by ligaments. In mesenchymal compartments between the rays and inside the rays there are blood vessels, nerves, pigment cells and fibroblast like cells (Poss et al., 2003). When the fin is amputated the wound surface is covered with epidermis within hours and as early as 12 hours after injury the essential blastema starts to take form. This mass of highly proliferative mesenchymal cells is formed through migration and disorganization of fibroblasts and osteoblasts. Once the blastema is formed the regenerative outgrowth begins and this lasts until the caudal fin regains its original form. This is achieved with the formation of two distinct areas in the blastema, a distal highly proliferative compartment that steadily replaces the more proximal cells as they differentiate (Gemberling et al., 2013).
Unlike in mammals the main hematopoietic organ in adult zebrafish is the kidney, not the bone marrow. Cells from the myeloid line can be found in all stages of development in the kidney, with heterophil granulocytes being the most abundant. These heterophil granulocytes show strong myeloperoxidase activity in their granules, a characteristic enzyme of mammalian neutrophil granules. All evidence points to the zebrafish heterophil granulocyte being the cellular orthologue of the mammalian neutrophilic granulocyte both in form and function. Therefore, studying zebrafish neutrophils can lead to insights on human neutrophil function and behavior during inflammation (Lieschke et al., 2001). Zebrafish monocytes and macrophages are both also morphologically equivalent to their mammalian counterparts, being large cells with large phagosomes (Menke et al., 2011).
As myeloperoxidase is specific to fish neutrophils and their precursors it was possible to use the isolated myeloid-specific peroxidase (mpx or mpo) gene to create a stable transgenic zebrafish with EGFP (Enhanced Green Fluorescent Protein) labeled neutrophils by inserting the EGFP sequence downstream of the mpx promoter. (Renshaw et al., 2006) In a similar process the promoter of the
6
macrophage expressed gene 1 (mpeg1) orthologue in zebrafish was used to insert either EGFP or mcherry fluorescent proteins so these would be expressed specifically in macrophages labeling them (Ellet et al., 2011).
Recruitment of neutrophils and macrophages after amputation of the caudal fin on adult zebrafish has been characterized by Petrie et al. (2015). They observed neutrophil accumulation at the injury site starting at 6 hours post amputation (hpa), with peak accumulation at 3 days post amputation (dpa) with return to pre-amputation density levels by 7 dpa. Macrophages were observed arriving at the injury site much later than neutrophils at 3-4 dpa, achieving peak numbers at 6-8 dpa and then, while the numbers were gradually decreasing, at 14 dpa there was still a high amount of macrophages present, mostly in the newly formed tissues (Petrie et al., 2015).
However, while neutrophils are still the first myeloid cells activated and recruited to the wound upon tail fin amputation in zebrafish larvae, they begin gradually accumulating as soon as the injury occurs until reaching a peak at 6 hpa, after which they begin leaving the injury site. Then, the slower macrophages begin arriving in larger numbers at the injury and when at 12 hpa the tissue cells begin to proliferate, ending the inflammatory phase and beginning regeneration, macrophages stay in the newly formed tissues (de Oliveira et al., 2013; Li, L. et al., 2012).
While the timing in recruitment between adults and larvae has been observed to be quite different and in both cases it macrophages were showed to be essential for the proper regeneration of the amputated tissue (Li, L. et al., 2012; Petrie et al., 2015). And while inhibition of neutrophil recruitment after fin amputation in larvae affected resolution of the inflammatory response no data was found on the effects on the regenerative capacity (de Oliveira et al., 2013). Still, while neutrophil inhibition in adult zebrafish did not appear to have an effect on the fin regeneration rate their role in the inflammatory response cannot be discarded (Petrie et al., 2015).
1.3 - Diabetes Mellitus
The International Diabetes Federation estimates diabetes mellitus affected up to 366 million adults in 2011, and the number is expected to rise to 552 million by 2030. Factors for the expected rise in incidence of diabetes are the ageing population, rise of risk factors like obesity and sedentary lifestyles, and better health care extending the life expectancy of diabetes patients (Whiting et al., 2011). The currently published diagnostic criteria for diabetes from the World Health Organization (WHO) are: 1) a fasting plasma glucose ≥ 126 mg/dL on two or more occasions. 2) A 2-hour plasma glucose ≥ 200 mg/dL after ingesting a 75 g glucose load, called the oral glucose tolerance test. 3) A random plasma glucose of ≥ 200 mg/dL (WHO, 2006).
Diabetes mellitus (DM) is a metabolic disease characterized by chronic hyperglycemia in a fasting state resulting from chronic insufficiency of insulin, insensibility to insulin, or both. Insulin is a hormone whose main objective is the regulation of blood glucose levels. When blood glucose levels rise, like after a meal, it is produced and released from the pancreas and targets skeletal muscle, adipose tissue and the liver to maximize their capture and usage of the circulating glucose and thus contributing to the normalization of the blood glucose levels. Cases of diabetes typically fall under one of two categories type 1 or type 2 DM, although there are a few rare cases that do not fit into these categories. Diabetes may also manifest during pregnancy, known as gestational diabetes, and as a consequence of drug or chemical toxicity, or genetic disorders (American Diabetes Association, 2015).
Type 1 DM is caused by absence of insulin production usually due to an autoimmune disorder associated with the chronic inflammation of the pancreatic Langerhans islets, where the insulin producing β-cells reside. Due to its autoimmune nature, type 1 DM has been historically mostly diagnosed in children and teenagers, but it can also be diagnosed in adults. Type 1 DM is often called
7
insulin-dependent diabetes as the most common therapy for it is insulin injections. Nowadays there are automated insulin pumps and glucose monitors that allow for better and finer control of glycemic levels, therefore reducing the chance of over-dosage of insulin that could lead to insulin resistance and type 2 DM. Also, novel therapies like islet-cell transplantation are being tested in order to develop a cure for type 1 DM (Atkinson et al., 2014).
Type 2 diabetes is currently the most common form of diabetes, comprising around 90% to 95% of diagnosed cases. It stems from acquired peripheral insulin resistance, which means higher amounts of the hormone are required to activate the same level of cell response in the tissues. This leads to compensatory insulin hypersecretion from the pancreatic islets, which eventually leads to a decline of the secretory function due to overexertion. Therapy for this type of DM usually comprises of oral medication that reduces glucose absorption or that increases peripheral sensitivity to insulin and it can be complemented with insulin injections if necessary. But the main therapy to manage type 2 DM is lifestyle changes as the majority of these patients are obese. The expanding adipocytes create regions of micro-hypoxia that continuously trigger the inflammatory response leading to chronic inflammation (Forbes and Cooper, 2013; American Diabetes Association, 2015).
Overall and independently of type, DM is associated with a chronic inflammatory response that is currently believed to be further potentiated by a prolonged exposure to hyperglycemia. Beyond this, DM has been shown to be associated with several secondary disease complications. Among these, there are acute and severe ones that can rapidly compromise the patient’s health and result in their death, like diabetic ketoacidosis from high blood glucose concentrations (hyperglycemia) or a diabetic coma due to low blood glucose (hypoglycemia). But apart from those, the most devastating consequences of DM are its long-term vascular complications whose onset results from the chronic elevation of blood glucose levels and are also closely associated with the chronic inflammation evidenced in DM, among other factors. These complications can be divided into micro- and macrovascular. In the latter, vascular damage affects large caliber blood vessels such as arteries, leading to accelerated cardiovascular disease that can result in myocardial infarctions and cerebrovascular disease that can manifest as ischaemic strokes. The microvascular complications in turn are caused by damage inflicted onto small caliber blood vessels and comprehend pathologies such as retinopathy (eye disease), nephropathy (kidney disease) and neuropathy (neural damage) (Forbes and Cooper, 2013).
Diabetic retinopathy is associated with changes in vascular permeability, capillary degeneration, capillary micro aneurysms, and excessive formation of new blood vessels. This develops over many years with almost all type 1 and most type 2 patients exhibiting retinal lesions after 20 years of the disease (Frank, 2004).
Diabetic nephropathy is one of the major causes of end-stage renal failure characterized by a decline in the glomerular filtration rate. Duration of DM, high blood pressure, and inadequate glycemic control are associated with development and progression of diabetic nephropathy (Tziomalos and Athyros, 2015).
Diabetic neuropathy is a syndrome that affects both somatic and autonomic divisions of the peripheral nervous system. The diabetic peripheral neuropathy usually manifests by loss of sensation in the lower limbs, but in severe cases it can also affect the fingers and hands. There has been some debate over the inclusion of diabetic neuropathy with other microvascular complications as recent evidence shows it selectively targets sensory and autonomic neurons with little vascular involvement (Forbes and Cooper, 2013). Alongside other risk factors, such as impaired wound healing, cardiovascular complications and mechanical trauma, diabetic neuropathy is a major cause of another clinically relevant complication, the diabetic foot ulcer. In fact, over 25% of diabetes patients eventually develop a foot ulcer during their lifetime, with 85% of amputations being preceded by them (Boulton, 2014).
8
Importantly, an impaired wound healing response has been observed in diabetic patients and can be partially attributed to the dysregulation of inflammation in DM, that can translate into functional defects in the leukocytes relevant to the inflammatory response. For instance, neutrophils from diabetic patients and animal models have been shown to exhibit defective chemotaxis, phagocytosis and bactericidal activity. In experimental diabetic mouse models an impairment in the initial recruitment of neutrophils to the wounds as also been repeatedly reported. However, despite the initial impairment, at later time-points both neutrophils and macrophages have been observed to persist far longer and at higher concentrations than normal in diabetic wounds. With the deregulation of the switch of M1 phenotype macrophages to the M2 phenotype being associated with the retention of the wound in a pro-inflammatory state (Ochoa et al. 2007; Forbes and Cooper, 2013). In a related note, neutrophil depletion in diabetic mice has been shown to accelerate wound healing (Dovi et al., 2003)
The most effective way to reduce the risk of these serious secondary complications is to attain normoglycemia as soon as possible, as the main cause of complication is the prolonged exposure of tissues to hyperglycemia. Prolonged exposure to hyperglycemia leads to permanent epigenetic alterations such as changes in histone methylation in the genomic DNA, usually referred to as metabolic memory (MM). (Forbes and Cooper, 2013; Sarras et al., 2015).
1.3.1 - Zebrafish models of Diabetes Mellitus
When submitted to long periods of hyperglycemia zebrafish display similar complications to those of human patients and mammalian models, like nephropathy, retinopathy and impaired regeneration (Olsen et al., 2010). The zebrafish pancreas, unlike in other fish species, is similar to the mammalian pancreas in both anatomy and development. The zebrafish is capable of re-growing the pancreas after a pancreatectomy and regenerating the β-cells after chemical ablation, therefore regaining normal insulin production and glycemic values on its own (Moss et al., 2009). But even after regaining normal glycemic values and pancreatic function the organism retains metabolic memory of the exposure to hyperglycemia. This combined with the benefits of the model alluded in the earlier section means zebrafish should be a viable model to address diabetic complications, which like in humans, are still present even after normal glycemic values are restored (Sarras et al., 2015).
The most used way to induce diabetes in mouse and rat models is the injection of the diabetogenic drug streptozotocin (STZ). The drug specifically targets the pancreatic β-cells by entering them through the specific Glut2 transporter, causing DNA damage and triggering apoptosis, what that means is that injection of STZ should mimic type 1 DM. Initial findings about the effectiveness of this drug on adult zebrafish suggested a single intraperitoneal injection of 1g/kg dose, which yielded higher fasting blood glucose levels (FBGL) after only 3 days. However, after a week the animals appeared to be regenerating the damaged cells and FBGL were returning to normal (Moss et al., 2009). Further studies used multiple
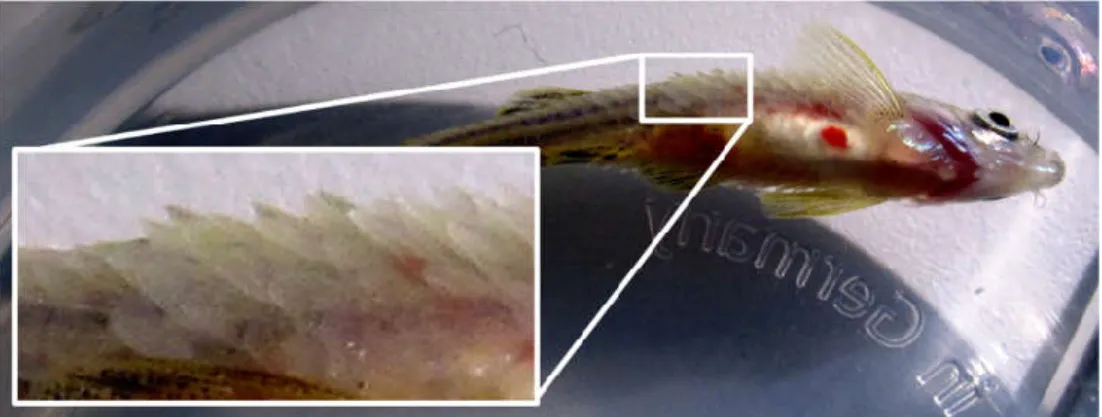
Figure 1.3 – Hyperglycemic fish have a reduced rate of caudal fin regeneration. Zebrafish caudal fin regeneration at 72 hours after amputation for a control injected fish (A) and a streptozotocin injected fish (B). Taken from Olsen et al. (2010).
9
intraperitoneal injections of a smaller 350mg/kg STZ dose and kept the animals at a lower temperature, 23°C instead of the usual 28°C, to slow fish metabolism and delay the regeneration of the damaged β-cells. When combined with booster injections this allowed for long term studies of the effects of hyperglycemia in the fish’s regenerative abilities. They also observed that upon amputation of the caudal fin hyperglycemic fish showed an impaired regeneration rate (Figure 1.3). Unfortunately, as it happens with rodents some animals could be inherently resistant to the effects of the drug (Olsen et al., 2010).
A different approach for the induction of hyperglycemia in zebrafish is immersion in a glucose solution so the animals passively absorb the glucose from their environment. This method resembles the onset of type 2 diabetes since the insulin production is not impaired but there still is hyperglycemia. There are several published variations for this method: 1) the longest employs alternating immersion with the fish being 24h in a glucose solution and then 24h in regular water (Connaughton et al., 2016); 2) in others, fish are submitted to continuous immersion in a glucose solution for up to 30 days (Capiotti et al., 2014; Carnovali et al., 2016). Concentration of the glucose solution used also varies, with a stepwise concentration increase from 1% to 3% throughout 8 weeks of alternating immersion protocol (Connaughton et al., 2016), or a constant concentration no higher than 4% to avoid lethality (Carnovali et al., 2016).
In addition, a mutant line of zebrafish that displays impaired pancreatic development and mature β-cell function and survival has also been established. The pdx1 null homozygote mutants present reduced insulin production and viability but can survive until adulthood. In the meantime, these mutant fish exhibit hyperglycemia from early larval stages allowing for long term studies of diabetic complications (Kimmel et al., 2015).
1.4 - Myocardial Infarction
Myocardial infarction (MI) is a cardiovascular disease and a major cause of mortality and disability world-wide, often being the manifestation of secondary cardiovascular complications of other diseases like diabetes. Pathologically MI is defined as extensive myocardial cell death due to prolonged ischemia (Thygesen et al., 2012).
Humans, like most mammals, have very little regenerative capability as adults. As such the wound healing after MI results in the formation of a non-contractile collagen-based scar tissue. Infarctions follow the same steps of wound healing as other injury, displaying the same overlapping phases: 1) Inflammatory response; 2) proliferative phase; 3) and maturation phase. In MI the main role of inflammation appears to be debridement of the infarcted area, with both neutrophils and macrophages having a role in the removal of the necrotic cardiomyocytes through phagocytosis. Simultaneously, and then as inflammation resolves, the native extracellular matrix is replaced by a provisional fibrin-rich matrix, the granulation tissue, that provides mechanical support to prevent tissue rupture while also assisting the migration of leukocytes and activated fibroblasts. There is also promotion of angiogenesis to ensure the proliferating cells are receiving the needed nutrients and oxygen. As the infarction enters the maturation phase the fibrin-rich provisional matrix is replaced by a permanent collagen matrix, originating the non-contractile scar tissue. (Frangogiannis, 2015).
1.4.1 - Zebrafish as a model of cardiac regeneration
Zebrafish have a closed single loop circulatory system, unlike mammals which have a closed double loop circulatory system. Their heart is situated in the anterior part of the body cavity, ventral to the esophagus (Figure 1.4a). Deoxygenated blood passes through the sinus venosus (thin wall made of connective tissue), before entering the atrium (thin muscular wall and thin trabeculae), which then passes to the ventricle (thick muscular wall and a spongy inner layer with many trabeculae), and finally through
10
the bulbous arteriosus (thick wall of fibro-elastic tissue and smooth muscle cells) from there it goes into the ventral aorta and then brachial arteries in the gills where it is re-oxygenated (Menke et al., 2011).
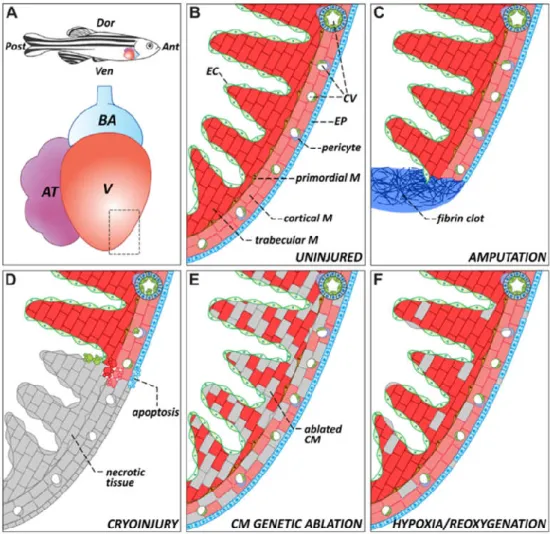
As expanded upon earlier, the zebrafish show amazing regenerative capabilities that include complete heart regeneration after damage. There are several different methods that have been used to induce injury in order to study heart regeneration: amputation of the ventricular apex, cryoinjury, genetic ablation, and hypoxia and re-oxygenation (Figure 1.4c-f) (González-Rosa et al., 2017).
Cryoinjury has long been used to induce MI in mammal models and is a procedure that allows to replicate mammal MI in zebrafish than the other methods. Cryoinjury causes massive localized cardiomyocyte death, just like what is seen in MI, allowing to more accurately analyze the processes that lead to heart regeneration after massive cell death (Chablais et al., 2011), also being considered an equivalent of the arterial ligation method also commonly used in mouse models (van den Bos et al., 2005). In the first few days after induction of MI by cryoinjury increased blood flow to the area is observed. By 24 hours after the MI fibrotic tissue has begun to replace the necrotic myocardium, but it is at 3 days that the full extent of the injury can be measured as the fibrotic tissue fully replaced the necrotic tissues. Beyond this point a collagen scar can also be seen in the injury and surrounding areas.
Figure 1.4 - Representative schematics about the zebrafish heart anatomy and injuries. (A) Heart position and anatomy in zebrafish. (B) Close up of normal zebrafish ventricle wall anatomy. (C-F) Different types of injury that can be induced on the zebrafish heart to study its regeneration: amputation, cryoinjury, genetic ablation and hypoxia/reoxygenation. Key: Ant, anterior; AT, atrium; BA, bulbus arteriosus; CM, cardiomyocyte; CV, coronary vasculature; Dor, dorsal; EC, endocardium; EP, epicardium; M, myocardium; Post, posterior; V, ventricle; Ven, ventral. Taken from González-Rosa et al., (2017).
11
The MI heals from the borders of the injury leading to a reduction of its size over time, with the collagen and fibrotic scar tissues being replaced by new cardiomyocytes resulting of the proliferation of pre-existing ones (González-Rosa et al., 2011).
1.4.2 - Neutrophil recruitment in Animal models of Myocardial Infarction
Neutrophil recruitment upon MI has been scarcely address so far. In mouse models, it has been observed that under normal conditions no neutrophils can be detected in the heart tissue (Li, W. et al., 2012). After induction of MI by ligation of the left anterior coronary artery, a peak of neutrophil recruitment was observed between 1 and 3 days after infarction. By 5 days after MI the quantity of neutrophils had dropped considerably but even after 7 days it was still significantly higher than what was observed in the uninjured hearts (Ma et al., 2016).
As for the zebrafish model, very little work has been published so far about neutrophil recruitment after heart injury. Neutrophil recruitment has been tracked for the first 24 hours after induction of cardiac inflammation via injection of nanoparticles. Significant increases in relation to the control of the quantity of neutrophils in the heart were detected at 2, 4 and 24 hours after induction (Duan et al., 2016).
1.5 – Objectives
The first objective of this master’s work was to analyze the recruitment of neutrophils and macrophages during the inflammatory response associated with the caudal fin regeneration in a diabetes mellitus zebrafish model. As outlined previously, the zebrafish has been shown to exhibit impaired tissue regeneration in a hyperglycemic condition. Here, our aim was to address whether such impairment could be correlated with an altered leukocyte recruitment during the inflammatory phase of the regenerative process.
Diabetes mellitus has been further demonstrated to underlie several cardiovascular complications, such as myocardial infarction among others. At the time this work was performed, little was known on the enrollment and the role of neutrophils in myocardial infarction in humans as well as in animal models, and more precisely, in the zebrafish. As such, our second objective was to analyze neutrophil recruitment during the inflammatory response and tissue regeneration following the induction of myocardial infarction in zebrafish.
12
Chapter 2 – Materials and Methods
2.1 - Animal Model
2.1.1 - Zebrafish Husbandry
Zebrafish were kept in the Zebrafish Facility at IMM-JLA standard Tecniplast rack system with 28ºC water temperature and optimal pH and salinity at a density of 3 fish per liter until needed. During workdays fish were fed two meals, the morning meal always being dry food and the afternoon meal being either dry food or live feed, whenever available. On weekends and other non-work days they were fed a single dry food meal. During assays the feeding schedule was kept as close to the facility standard as possible and the water of tanks not connected to the system was exchanged every two days unless otherwise stated.
Preliminary tests and optimization for each method were made with seemingly healthy facility zebrafish of similar age to the test fish to be used which were meant to be sacrificed, respecting the 3Rs rule. The project was approved by the ethical committee and DGAV.
2.1.2 - Transgenic Lines
Several different transgenic lines were used for the experimental procedures described below. For neutrophil recruitment analysis the transgenic Tg(mpx:GFP) was used. In this line, the GFP gene was placed under the control of the promotor of the mpx gene, which encodes the myeloperoxidase protein, so that the GFP protein is only expressed in zebrafish neutrophils (Renshaw et al., 2006). Macrophage recruitment analysis was attempted using Tg(mpeg1:EGFP). The enhanced GFP (EGFP) gene was placed under the promotor of the macrophage expressed gene 1 (mpeg1) (Ellet et al., 2011).
Before the start of any imaging assay the Tg(mpx:GFP) and Tg(mpeg1:EGFP) fish were screened to discard unhealthy animals that presented a notorious increase of labeled immune cells throughout their bodies in comparison to what is normally observed in these transgenic zebrafish lines.
2.1.3 - Anesthetics & Recovery
In most of the experiments and procedures performed in this work, fish were anesthetized with a 0.16 g/L solution of Tricaine-S (Western Chemical, Inc.), also referred to as MS-222, that is the standard anesthetic in the facility. This solution has been shown to alter blood glucose readouts, and therefore it was not employed in procedures and experiments performed for the Diabetes Mellitus model (Eames et al., 2010). For these a 0.1% 2-Phenoxyethanol (Sigma-Aldrich) solution was used instead as published by Intine et al. (2013).
To anesthetize the fish, they were placed in one of the anesthetic solutions and left in it until unresponsive with little gill movement. After the procedures the fish were placed in 1L breeding boxes with fresh system water to recover normal natatory capacity. For longer procedures recovery of gill movement was stimulated by gently creating water flow over the gills using a plastic Pasteur pipette. When fully recovered the fish were kept isolated in breeding boxes (maximum two per box) or on a 3.5L tank (up to 10 fish of the same test condition) isolated from the rack system.