UNIVERSIDADE FEDERAL DA PARAÍBA
CENTRO DE CIÊNCIAS EXATAS E DA NATUREZA
ADRIANO MEDEIROS DE SOUZA
AS ÁREAS DE ENDEMISMO DOS OPILIONES
(ARACHNIDA) DA FLORESTA ATLÂNTICA AO NORTE
DO RIO SÃO FRANCISCO, BRASIL.
ADRIANO MEDEIROS DE SOUZA
AS ÁREAS DE ENDEMISMO DOS OPILIONES
(ARACHNIDA) DA FLORESTA ATLÂNTICA AO NORTE
DO RIO SÃO FRANCISCO, BRASIL.
Dissertação apresentada ao Programa de Pós Graduação em Ciências Biológicas (Zoologia), da Universidade Federal da Paraíba, como parte dos requisitos à obtenção do título de Mestre em Ciências Biológicas (Zoologia) na referida instituição.
Orientador: Prof. Dr. Marcio Bernardino da Silva
A729a Souza, Adriano Medeiros de.
As áreas de endemismo dos opiliones (arachnida) da floresta atlântica ao norte do rio São Francisco, Brasil / Adriano Medeiros de Souza.- João Pessoa, 2013.
155f. : il.
Orientador: Marcio Bernardino da Silva Dissertação (Mestrado) – UFPB/CCEN
1. Zoologia. 2. Biogeografia histórica. 3. Opiliões - Brasil. 4. Endemicidade - análise. 5. Parcimônia de endemicidade - análise. 6. Elementos bióticos - análise.
ADRIANO MEDEIROS DE SOUZA
AS ÁREAS DE ENDEMISMO DOS OPILIONES
(ARACHNIDA) DA FLORESTA ATLÂNTICA AO NORTE
DO RIO SÃO FRANCISCO, BRASIL.
Dissertação apresentada ao Programa de Pós Graduação em Ciências Biológicas (Zoologia), da Universidade Federal da Paraíba, como parte dos requisitos à obtenção do título de Mestre em Ciências Biológicas (Zoologia) na referida instituição.
Data de Aprovação: _____/_____/____
Nota: ___________________________
BANCA EXAMINADORA:
_____________________________________
Prof. Dr. MARCIO BERNARDINO DA SILVA (Orientador) Departamento de Sistemática e Ecologia - UFPB
_____________________________________
Prof. Dr. ALEXANDRE VASCONCELLOS (1º Titular) Departamento de Sistemática e Ecologia - UFPB
_____________________________________
Prof. Dr. RICARDO PINTO DA ROCHA (2º Titular) Instituto de Biociências - USP
_____________________________________
Profª. Drª. MARIA REGINA DE VASCONCELLOS BARBOSA (Suplente) Departamento de Sistemática e Ecologia - UFPB
__________________________________________________ Prof. Dr. ULISSES DOS SANTOS PINHEIRO (Suplente) Departamento de Zoologia - UFPE
Agradecimentos:
Essa provavelmente é a parte do trabalho que mais me deu trabalho em redigir. Antes de qualquer coisa peço desculpas às pessoas que irei esquecer, pois a minha memória é péssima. Tentarei citar as pessoas por ordem cronológica de participação. Vamos lá: gostaria de agradecer primeiramente aos meus pais (José Domingos de Souza “Seu José” e Marinalva Maria de Medeiros) pelo apoio incondicional nesses dois anos de mestrado. Contei muito também com o apoio de meus irmãos (Flávio e Flávia) e claro dos meus sobrinhos e afilhados, Bruno e Gustavo, que muito me fizeram rir nesses dois anos de mestrado.
Queria agradecer ao parceiro de trabalho, de coleta e de muitas cervejinhas, André Felipe. Se não fosse por esse cara pálida com sua emblemática frase: “A vida é feita de emoções!” talvez eu nunca tivesse me arriscado a sair da minha zona de conforto e me aventurado com os opiliões. Valeu Zé!
Outra pessoa importante nessa etapa inicial foi o Prof. Dr. Ricardo Pinto-da-Rocha. Não posso deixar de citar o nome dele. Obrigado por ter acreditado em mim e não me deixar desistir.
Entretanto há uma pessoa que eu nem consigo definir a sua importância nesse trabalho: o Prof. Dr. Marcio Bernardino da Silva, ou simplesmente, “MBS”! O que eu posso dizer de um cara que me ajudou em tudo o que podia? Foi muito mais do que um orientador durante esses dois anos, foi um amigo, um parceiro mesmo. Simplesmente meu sincero obrigado MBS! Devo essa dissertação a você!
Também gostaria de agradecer à Coordenação do Programa de Pós Graduação em Ciências Biológicas (Zoologia) por sempre tentar me ajudar quando eu solicitei. Aproveito aqui para agradecer a todos os professores do referido programa que eu tive o prazer de assistir alguma aula. Todos vocês contribuíram muito com essa etapa da minha vida.
Agradeço também a Comissão de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) e ao REUNI pela bolsa a mim concedida. Agradeço também ao auxílio concedido pela comissão do XVII International Congress of Arachnology para a participação no III Congresso Latinoamericano de Aracnología.
“AracnoJampa”: Luiz Paulo (Buchecha), Everton (Arroz), Arthur, Paulo (Pessimista), Nathália, Simara, Diva, Karla, Géssica, Aurora e Lu. Vocês tornaram as minhas coletas super divertidas, sem contar os dias no laboratório. Cabe aqui mencionar também o pessoal do Laboratório de entomologia: Carol, Aline, Matilde, Renato, Jean, Renata, Daniel, Layla, Ana Cláudia e Wellington. Valeu pelos dias de risada galera!
Também gostaria de agradecer aos meus amigos de graduação José Renato e Walter Dennis. Vocês tornaram as minhas quintas mais divertidas com o #BeerDay.
Gostaria de agradecer também ao Programa em Biodiversidade do Semiárido, especialmente ao Prof. Leonardo de Sousa Carvalho (UFPI) por permitir que eu participasse do projeto.
Aos meus colegas de turma de pós também fica aqui o meu obrigado. Aline, Laís, Cristiano, Alvino e principalmente Rafael e Emerson (pela convivência diária), agradeço pelos momentos de descontração.
Esse trabalho também não seria possível sem a colaboração de diversas pessoas que estão ligadas as diversas unidades de conservação nas quais tive a oportunidade de trabalhar. Ficam aqui os meus agradecimentos aos chefes das seguintes UCs: Damásio (Reserva Biológica da Serra Negra), Aronílson (Reserva Municipal Mata do Mucuri) e, sobretudo a Luiz Aleixo, vulgo “Lula do Mel” (Parque Natural Municipal João Vasconcelos Sobrinho) por todo o suporte fornecido.
Agradeço também à Banca Avaliadora por aceitar o convite. Sei que todos têm suas obrigações, mas mesmo assim fizeram esse esforço a fim de contribuir com o presente trabalho.
Fica aqui registrado também a minha gratidão ao Prof. Dr. Adriano Brilhante Kury, curador da Coleção de Aracnídeos no Museu Nacional (RJ), por me receber na referida Coleção e permitir o meu trabalho.
Não poderia deixar de agradecer a senhorita Cassia Maria Rodrigues, por aguentar toda a minha chatice e mau humor. Além do que, se não fosse pelo convite que ela me fez no fim de 2006, talvez eu nunca tivesse começado a trabalhar com aracnídeos. Eu te amo princesa, nunca se esqueça disso!
LISTA DE TABELAS
LISTA DE FIGURAS
SUMÁRIO
Resumo... 11
Abstract... 12
Apresentação... 14
1.
INTRODUÇÃO...
151.1 ÁREAS DE ENDEMISMO: A UNIDADE BÁSICA DA BIOGEOGRAFIA HISTÓRICA... 15
1.2 AS PROPOSTAS DE REGIONALIZAÇÃO BIOGEOGRÁFICA DA MATA ATLÂNTICA BRASILEIRA... 17
1.3 A MATA ATLÂNTICA NORDESTINA... 19
1.4 OS BREJOS DE ALTITUDE... 20
1.5 OS OPILIÕES DA MATA ATLÂNTICA... 22
2.
JUSTIFICATIVA...
243.
OBJETIVOS...
254.
MATERIAL E MÉTODOS...
264.1 TÁXONS UTILIZADOS... 26
4.2 COMPILAÇÃO DAS OCORRÊNCIAS... 26
4.2.1 Coletas... 27
4.2.2 Compilação dos dados da literatura... 28
4.2.3 Visita aos Museus... 28
4.2.4 Georreferenciamento... 28
4.3 ANÁLISES... 29
4.3.1. Análise de Endemicidade (NDM)... 29
4.3.2. Análise de Parcimônia de Endemismo (PAE)... 30
4.3.3. Análise de Elementos Bióticos (BEA)... 31
4.3.4. Critérios combinados... 32
4.4 PROTOCOLO PARA DELIMITAÇÃO DAS ÁREAS DE ENDEMISMO... 33
5.
RESULTADOS...
365.1 NDM... 36
5.2 PAE... 37
5.4 ASSOCIANDO OS MÉTODOS NUMÉRICOS AOS CRITÉRIOS
COMBINADOS... 39
6.
DISCUSSÃO
... 426.1 AVALIAÇÃO DOS MÉTODOS: NDMD, PAE E BEA... 42
6.1.1 O tamanho da grade... 42
6.1.2. Avaliando o desempenho dos métodos numéricos... 43
6.2 AS ÁREAS DE ENDEMISMO DO NORDESTE: ORIGENS E RELAÇÕES HISTÓRICAS... 45
7.
CONCLUSÃO
... 518.
REFERÊNCIAS
... 52Resumo
A Mata Atlântica é uma das regiões mais ricas do mundo, tanto em diversidade de espécies quanto em endemismo. Devido a isso e ao grau de devastação desse bioma, há alguns anos ela foi enquadrada como um dos 25 “hotspots” mundias de biodiversidade. Entretanto, as relações históricas entre os diferentes setores da Mata Atlântica são pouco conhecido. Uma etapa fundamental é a delimitação das áreas de endemismo, que são unidades básicas para análises biogeográficas. Os estudos nessa área avançaram bastante na Mata Atlântica do sul e sudeste com o uso das distribuições dos opiliões que ocorrem nessa região, entretanto, permanece uma lacuna com relação à Mata Atlântica nordestina. Diante disso, o objetivo do presente estudo foi delimitar as áreas de endemismo da Mata Atlântica localizada ao norte do rio São Francisco, utilizando as distribuições de espécies de Opiliones, além de comparar os resultados obtidos pelas diferentes metodologias utilizadas e avaliar a influência do tamanho das células sobre os resultados. Ao todo, foram utilizadas 1581 ocorrências de 224 espécies, sendo que, dessas, 18 ainda não estão descritas. As ocorrências foram obtidas por coletas, revisão de literatura e visita a museus. Foram utilizados três métodos numéricos para busca de áreas de endemismo que trabalham usando a ocorrência das espécies num conjunto de células de uma grade: a Análise de Endemicidade (NDM), a Análise de Parcimônia de Endemicidade (PAE) e a Análise de Elementos Bióticos (BEA). Para os três métodos foram aplicados três grades: uma grade menos detalhada (2° X 2°), uma grade intermediária (1° X 1°) e uma grade mais detalhada (com células de 0,5° X 0,5°). Em seguida, as áreas de endemismo foram delimitadas aplicando, sobre os resultados das análises numéricas, um protocolo baseado numa série de critérios combinados que, por sua vez, derivam de conceitos de áreas de endemismo descritos na literatura. Ao todo, foram delimitadas treze áreas de endemismo para a Mata Atlântica, sendo que, dessas, três correspondem à Mata Atlântica nordestina: a Área de Endemismo Bahia (BA), a Área de Endemismo Brejos Cearenses (BCE) e a Área de Endemismo Pernambuco (PE). Os resultados obtidos pela NDM e pela PAE foram similares, enquanto que a BEA apresentou resultados completamente diferentes e arbitrários. O tamanho das células utilizadas teve influência nas análises, tanto na quantidade de áreas encontradas, quanto na quantidade de células incluída em cada área. A maior quantidade de dados utilizada tornou áreas delimitadas mais robustas, sobretudo aquelas da Mata Atlântica nordestina, quando comparada a trabalhos anteriores. Essas áreas, assim como aquelas da Mata Atlântica sul e sudeste, provavelmente correspondem às áreas de refúgios florestais pleistocênicos, quando a Mata Atlântica inteira experimentou ciclos de expansão e retração. Essas oscilações estão associadas aos ciclos de glaciação e aquecimento que ocorreram durante o Quaternário, sendo que, na Mata Atlântica nordestina, a expansão de seus limites provavelmente ocasionou um contato com a Floresta Amazônica. Outras causas prováveis foram às transgressões marinhas ou tectonismo, no caso, aplicados a região da Baía de Todos os Santos.
Abstract
The Atlantic Forest is one of the richest regions of the world, both in species diversity, as in endemism. Due to this and to the degree of devastation, a few years ago, this biome was considered one of the 25 worldwide biodiversity hotspots. However, the historical relationships between different sectors of the Atlantic Forest are poorly understood. A critical step in that knowledge is the delimitation of the areas of endemism, which are basic units for biogeographic analyzes. Studies performed in this subject have advanced enough, both south and southeastern Atlantic Forest, by using the distribution patterns of harvestmen which occur into this region, nevertheless, a gap remains regarding to the northeastern Atlantic Forest. Therefore, this study aims to delimit the areas of endemism in Atlantic Forest north São Francisco river, by using Opiliones species distributions, also comparing the obtained results by different methodologies and finally evaluating the influence of the size of the cells on the results. In total, we used 1581 occurrences of 224 species, and of these, 18 are not yet described. The occurrences were obtained of field collecting, literature and museums. We used three numeric methods to search for areas of endemism that work using the occurrence of species in a set of cells on a grid: Endemicity Analysis (NDM), Parsimony Analysis of Endemicity (PAE) and Biotic Element Analysis (BEA). For the three methods were applied three grids: a less detailed grid (2° X 2°), a intermediated grid (1° X 1°) and one more detailed grid (with 0,5° X 0,5° cells). Afterwards, the areas of endemism were delimited by applying on the results of the numeric analyzes a protocol based on a number of combined criteria derived from areas of endemic concepts described in the literature. Altogether, thirteen endemism areas were delimited for Atlantic Forest, and, three of these correspond to the northeastern Atlantic Forest: Area of Endemism Bahia (BA), Area of Endemism Brejos Cearenses (BCE) and Area of Endemism Pernambuco (PE). The results from NDM and PAE were similar, whereas BEA results were entirely different and arbitrary. The size of cells had influences in the analysis, both on the number of areas found, as the number of cells included in each area. The largest amount of data used, turned limited areas into more robust, especially those of the northeastern Atlantic Forest, if compared to previous work. These areas, as well as those from south and southeastern Atlantic Forest probably correspond to areas of Pleistocene forest refuge, when the entire Atlantic Forest experienced cycles of expansion and contraction. These oscillations are associated with the cycles of glaciation and warming that occurred during the Quaternary, where, the expansion of northeastern Atlantic Forest boundaries probably led it to a contact with the Amazon Rainforest. Other likely causes were marine transgressions or tectonism, in this case, applied to the region of Baía de Todos os Santos.
AS ÁREAS DE ENDEMISMO DOS OPILIONES (ARACHNIDA) DA
FLORESTA ATLÂNTICA AO NORTE DO RIO SÃO FRANCISCO,
Apresentação
A Mata Atlântica, que constitui um dos 25 hotspots mundiais de biodiversidade, é uma das áreas mais ricas em número de espécies, sendo que uma grande parte dessa biota não existe em mais nenhum lugar do globo. Entretanto, as causas de tamanho grau de diversidade e endemismo ainda são pouco compreendidas.
Os métodos da Biogeografia Histórica são uma poderosa ferramenta no auxílio da elucidação e compreensão de tais processos. Uma das primeiras etapas em estudos de cunho biogeográfico é a delimitação de áreas de endemismo. Entretanto, os métodos numéricos para essa delimitação foram pouco usados, carecendo de bancos de dados abrangentes e detalhados, o que não era possível no passado. Mas com o desenvolvimento das grandes coleções de organismos disponíveis nos museus, formadas por diversas gerações de pesquisadores, uma grande quantidade de dados de ocorrência mais detalhados encontram-se a disposição para a realização de tais análises, o que assegura resultados mais robustos. Diversos táxons já foram estudados usando essa abordagem, dentre eles os opiliões. As primeiras pesquisas com o grupo nesse âmbito iniciaram no início dos anos 2000 e mostram resultados cada vez mais promissores. Entretanto, um setor da Mata Atlântica permanece como uma lacuna nos trabalhos anteriores: a Mata Atlântica ao norte do Rio São Francisco.
Esse setor da Mata Atlântica é de extrema importância por ser uma das regiões mais devastadas desse bioma. Além disso, essa região apresenta uma série de encraves localizados no meio da vegetação de Caatinga: os Brejos de Altitude. Esses encraves são pouco compreendidos acerca de suas origens e relações com a Mata Atlântica costeira, o que só reforça a necessidade de mais pesquisas nessa região.
1.
INTRODUÇÃO
1.1. ÁREAS DE ENDEMISMO: A UNIDADE BÁSICA DA BIOGEOGRAFIA HISTÓRICA.
Dentre as diversas áreas da Biologia, a Biogeografia é aquela que tem como interesse o estudo dos padrões distribucionais dos seres vivos, no âmbito ecológico e histórico (Brown & Lomolino, 1998). É uma ciência que tem por essência a interação com outras áreas de conhecimento, como a paleontologia, genética, ecologia, etc. Os estudos em biogeografia basicamente resumem-se em tentar explicar porque as espécies estão onde estão, o que na prática é a síntese do processo evolutivo, levando em conta o tempo, forma e espaço (Crisci et al., 2003).
Os padrões de distribuição dos organismos pelo globo são motivo de interesse de diversos pesquisadores desde o século XVIII (Brown & Lomolino, 1998). O botânico sueco e pai da taxonomia Carl Von Linné (1707-1776) já se preocupava em tentar explicar a distribuição dos organismos, a partir da ideia criacionista de que Deus criou as espécies em um determinado local e ali seria o seu centro de origem. Outros pesquisadores posteriores a Linné também se debruçaram sobre o tema, como o também botânico suíço Augustin Pyrame de Candolle (1778-1841). A este último, é atribuída a autoria dos termos “endemismo” e “regiões biogeográficas”, que é hoje em dia o foco de diversas pesquisas em biogeografia histórica (um dos ramos da Biogeografia). A partir dos padrões de distribuição de plantas, de Candolle desenvolveu, em 1840 uma proposta de divisão do globo em regiões biogeográficas (Nelson & Platnick, 1981).
Tão importante quanto a contribuição feita por de Candolle, foram as pesquisas do naturalista britânico Alfred Russel Wallace (1823-1913). Seguindo a divisão proposta por Sclater em1858, com base na distribuição de aves, Wallace formalizou, em 1876, uma divisão biogeográfica do globo em seis regiões zoogeográficas (Neártica, Neotropical, Paleártica, Etiópica, Oriental e Australiana). Essa divisão foi aceita por diversos pesquisadores que trabalharam com o tema posteriormente e é aceita até os dias atuais. Até esse momento, as pesquisas eram mais de caráter especulativo, sem métodos claros e objetivos.
estavam em constante movimento e teve um profundo impacto na biogeografia histórica, ao dar fim ao pensamento dispersionista corrente até então (Crisci et al., 2003). O segundo foi o surgimento da sistemática filogenética, desenvolvida pelo entomólogo alemão Willi Hennig em 1955, que forneceu todo o arcabouço teórico e metodológico para a inferência de relações entre as espécies (Wiley, 1988). Todavia, os novos métodos propostos esbarravam num problema: a unidade biogeográfica a ser usada durante as análises.
Os primeiros trabalhos usando a nova abordagem embasada nos conceitos propostos por Hennig sugeriam que deveriam ser usadas como unidade básica para as análises biogeográficas as áreas de endemismo, que seriam as menores unidades históricas (Nelson & Platnick, 1981). Entretanto, o conceito de área de endemismo é debatido até hoje (Nelson & Platnick, 1981; Axelius, 1991; Platnick, 1991; Morrone, 1994; Harold & Mooi, 1994; Hausdorf, 2002; Crother & Murray, 2011).
Como já foi dito, o primeiro a mencionar o termo endemismo foi Candolle em seu livro “Statistique de la Familie des Composées” de 1838. Embora não tenha deixado explícita a definição, uma característica importante para o reconhecimento de uma área de endemismo foi citada pelo mesmo: a ocorrência de espécies que não existem em mais nenhum outro lugar (Espinosa-Organista & Llorente-Busquets, 1993; Crisci et al., 2003). Para Nelson & Platnick (1981), uma área de endemismo é uma pequena região que tem um número significante de espécies que não ocorrem em mais nenhum outro lugar.
Baseado no conceito proposto por Nelson & Platnick (1981), Platnick (1991) definiu área de endemismo como a congruência distribucional de duas ou mais espécies. Essa definição também é seguida por Morrone (1994) e por Humpries & Parenti (1999). A importância de se usar áreas de endemismo como unidade básica para análises também é citada por Axelius (1991). Segundo esse autor, uma análise biogeográfica que não utilizasse as áreas de endemismo como unidade básica poderia gerar resultados arbitrários.
sugerindo então o uso de “Elementos bióticos”, que seriam um grupo de táxons cujas distribuições são significativamente mais similares entre sim do que com qualquer outro grupo.
Alguns autores também já discutiram uma forma objetiva de se delimitar as áreas de endemismo. Alguns trabalhos sugerem a sobreposição de uma grade sobre um mapa com as distribuições individuais de cada espécie (Morrone, 1994; Morrone & Escalante, 2002; Szumik et al., 2002; Hausdorf & Hennig, 2003; Hennig & Hausdorf, 2004; Szumik & Goloboff, 2004). A unidade básica para as análises passa a ser cada célula da grade, sendo que agrupamentos dessas células podem ser interpretadas como áreas de endemismo. Foram propostos como métodos de busca por áreas de endemismo, a utilização da máxima parcimônia (Morrone, 1994), de um teste de agrupamento (Hausdorf & Hennig, 2003; Hennig & Hausdorf, 2004) e a utilização de critérios de otimização (Szumik et al., 2002; Szumik & Goloboff, 2004).
Entretanto, um fator muito importante a todos os métodos que utilizam células como base para a busca de áreas é o tamanho da grade utilizada (Morrone & Escalante, 2002). Essa característica foi pouco discutida, apesar de ter enorme importância sobre os resultados, já que, dependendo do tamanho e da posição da grade utilizada, alguns padrões de endemismo podem não ser reconhecidos pelas análises (Morrone & Escalante, 2002).
1.2. AS PROPOSTAS DE REGIONALIZAÇÃO BIOGEOGRÁFICA DA MATA ATLÂNTICA BRASILEIRA.
Várias propostas foram realizadas com o intuito de setorizar a Mata Atlântica. Uma das primeiras propostas foi a de Cabrera & Willink (1973) baseada em dados de distribuição de animais e vegetais. Segundo os autores, esse domínio fitogeográfico estaria dividido em duas províncias biogeográficas: “Província Paranaense” (abrangendo desde o extremo sul do Brasil, a oeste da Serra do Mar e centro do Rio Grande do Sul, até o extremo nordeste da Argentina e Paraguai) e “Bosque Atlântico” (toda a costa brasileira, estendendo-se da latitude 7º S até 30ºS). Entretanto, os autores não sugerem áreas de endemismo em sua divisão, funcionando mais como unidades ecológicas ou morfoclimáticas.
desde o estado da Bahia até o norte do estado do Espírito Santo; “Rio de Janeiro”, localizado desde o sul do Rio Doce no Espírito Santo até o estado do Rio de Janeiro. Um quarto centro de endemismo indicado pelo autor corresponde a uma área de encrave de floresta no meio de uma vegetação típica de cerrado, localizada ao norte da cidade de Brasília, que segundo o próprio autor estaria relacionada à Mata Atlântica.
Utilizando as ocorrências de alguns grupos de mamíferos, Costa et al. (2000) conseguiram reconhecer três áreas de endemismo: “Pernambuco”, “SE coast” e “Paulista”. Entretanto, os autores ressaltam a possibilidade de não terem sido reconhecidas algumas áreas devido ao tamanho das células da grade empregada. Já Morrone (2001) propôs dividir a Mata Atlântica em três áreas biogeográficas: “Bosque Paranaense” (localizado no Sudeste do Brasil, noroeste da Argentina e oeste do Paraguai), “Bosque de Araucaria angustifolia” (localizado no Sul do Brasil) e “Bosque Atlântico Brasileño” (que seria toda a Mata Atlântica costeira, nunca excedendo de 100 km do litoral).
Silva et al. (2004) utilizaram 104 espécies de aves da Mata Atlântica para a delimitação de áreas de endemismo. Nesse estudo, os autores conseguiram reconhecer quatro áreas de endemismo: Pernambuco, Bahia Central, Costa da Bahia e Serra do Mar. Com base nas distribuições de aves, primatas e borboletas, Silva & Casteleti (2005) propuseram a divisão da Mata Atlântica brasileira em cinco áreas de endemismo: quatro localizadas na Região Nordeste (Brejos Nordestinos, Pernambuco, Diamantina e Bahia) e uma localizada na Região Sudeste (Serra do Mar). Entretanto, este trabalho não usou nenhum método numérico explícito para propor esta divisão.
Entretanto, os autores ressaltam que essa área carece de mais dados para a correta delimitação dos seus limites.
Apesar de algumas poucas diferenças, os vários estudos convergem na divisão biogeográfica da Mata Atlântica em quatro grandes blocos. O primeiro está relacionado à floresta de araucárias mais ao sul. Outro bloco seria mais relacionado às florestas localizadas na Serra do Mar, sendo que, a depender do táxon utilizado e do método, esse bloco pode surgir como uma única área de endemismo (Costa et al., 2000; Silva et al., 2004) ou fragmentado em áreas de endemismo menores (Pinto-da-Rocha et al., 2005; DaSilva & Pinto-da-Rocha, 2011). Os dois últimos estão relacionados às florestas costeiras ao sul da Bahia e ao norte do Rio São Francisco. Entretanto, o limite entre as áreas varia muito, dependendo do grupo abordado e da metodologia utilizada.
1.3. A MATA ATLÂNTICA NORDESTINA.
A Mata Atlântica constitui uma das áreas com maior biodiversidade do mundo (Galindo-Leal & Câmara, 2005). Entretanto, devido à forte pressão antrópica, esse ecossistema encontra-se entre os mais ameaçados do planeta. A área de abrangência inicial desse ecossistema foi estimado em cerca de 1 a 1,5 milhão de km2, sendo que atualmente restam pouco mais de 10% da área original (Ribeiro et al., 2009). Mesmo apresentando tamanho grau de devastação, a Mata Atlântica ainda abriga grande diversidade biológica, possuindo boa parte de sua fauna e flora endêmica. Esta característica peculiar desse ecossistema fez com que o mesmo fosse incluído em uma das 25 prioridades mundiais para a conservação da biodiversidade mundial (Pôrto et al., 2005). O conceito de áreas prioritárias ganhou força nas últimas décadas e o termo “hotspot” passou a ser utilizado para designar ecossistemas que se encontram extremamente ameaçados e que abrigam uma alta diversidade biológica e endemismo. Embora representem cerca de 1,4% da superfície terrestre, essas áreas abrigam mais de 60% da biodiversidade do globo (Myers et al., 2000).
Embora considerada um hotspot de biodiversidade mundial, boa parte da fauna e flora da Mata Atlântica permanece desconhecida (Silva & Casteleti, 2005; Conservation International do Brasil et al., 2000). A situação agrava-se ainda mais no que diz respeito à Mata Atlântica localizada ao norte do rio São Francisco (Pôrto et al. 2005). Esta, provavelmente, é a região desse bioma que se encontra mais fragmentada.
e biogeografia, pois apresenta uma grande quantidade de animais e plantas endêmicos (Tabarelli & Santos, 2004). Devido a essa biota peculiar, essa região já foi mencionada por diversas vezes como uma área de endemismo da Mata Atlântica (Prance, 1982; Costa et al., 2000; Silva et al., 2004; Silva & Casteleti, 2005; DaSilva & Pinto-da-Rocha, 2011), o que só ressalta a sua importância. Também há um interesse nessa região pelo fato de haver vários encraves de floresta em meio à vegetação de Caatinga. Esses encraves são denominados de “Brejos de Altitude”.
1.4. OS BREJOS DE ALTITUDE.
A palavra “brejo” tem diversos significados no Brasil, a depender do estado ou região. De uma forma geral, o termo “brejo” refere-se a terrenos alagadiços com saturação de água no solo (Luft, 1993). Entretanto, na Região Nordeste, esse termo ganha um novo significado: passa a caracterizar espaços situados na Caatinga hipo e hiperxerófila, sob o domínio da umidade e influenciados pelo relevo, pelas massas de ar e pelo clima, o que propicia uma região de maior regime pluviométrico, com a ocorrência de chuvas orográficas. Elas garantem temperaturas mais amenas nos brejos em relação à Caatinga que está ao redor (Andrade-Lima, 1982). Essas regiões são localmente conhecidas por “Brejos”, “Brejos de Altitude”, “Brejos Nordestinos”, “matas serranas” (PB, PE) ou “matas úmidas” (CE).
Porém, há algumas nuances em torno do significado do termo, o que gerou diversas definições pelos pesquisadores que se debruçaram sobre o tema. Para Andrade & Lins (1964), a definição de Brejo de Altitude deveria levar em conta principalmente as questões hídricas e atmosféricas, já que, segundo os autores, esses fatores são os mais marcantes e preponderantes para a elevada pluviosidade acarretada. Já Andrade-Lima (1982), considerava, como fatores mais característicos dos brejos, as altitudes elevadas (acima de 500m) e sua posição em relação às correntes de ar, sobretudo aquelas vindas do sudeste. Esses pontos de vista mostram o quanto complexa é essa região.
entre 25 e 30 metros, havendo também um estrato arbustivo de 1 a 3 metros (Rizzini, 1997).
Segundo Bigarella & Andrade-Lima (1982) e Bigarella (1991), os Brejos de Altitude devem ter-se originado no Paleoceno, quando havia um manto contínuo de formação florestal na região onde atualmente se encontram os Brejos de Altitude. Com as modificações climáticas que ocorreram durante o Pleistoceno, como por exemplo, a glaciação de Würm, houve mudanças na vegetação com expansão daquela adaptada à seca, com a cobertura florestal inicial ficando restrita apenas ao topo das serras (Ab’ Saber, 1977).
As áreas consideradas Brejos de Altitude foram listadas por Vasconcelos Sobrinho (1971) apud Tabarelli & Santos (2004). Na proposta dele, haveria um total de 43 brejos de altitude pelo nordeste, espalhados pelos estados do Ceará, Paraíba, Pernambuco e Rio Grande do Norte. Entretanto, nos dias atuais, grande parte desses brejos encontra-se quase que totalmente devastado, principalmente devido à ocupação humana, à agricultura de subsistência praticada na região e a expansão da monoculttura de cana-de-açúcar (Luft, 1993; Barbosa et al., 2004; Tabarelli & Santos, 2004).
A degradação dessas regiões traz um sinal de alerta: uma região complexa e ainda pouco compreendida pelos pesquisadores no que toca os seus aspectos históricos, encontra-se a beira da extinção. Isso é alarmante, sobretudo porque os resultados de alguns estudos realizados na região mostram que há uma biota complexa na região, com algumas espécies típicas da Mata Atlântica e outras espécies mais relacionadas àquelas amazônicas (Vanzolini & Williams, 1970; Borges-Nojosa & Caramaschi, 2003). Ainda existem espécies endêmicas dos Brejos, como por exemplo, os escorpiões Hadrurochactas brejo (Lourenço, 1988) e Hadrurochactas araripe Lourenço, 2010, encontrados somente na Serra de Baturité (CE) e na Chapada do Araripe (CE), respectivamente. Também já foram documentados lagartos, aves e alguns grupos de insetos endêmicos dos Brejos de Altitude (Borges-Nojosa & Caramaschi, 2003), o que só ressalta a importância de mais pesquisas e da conservação dessa região.
das relações entre a Mata Atlântica ao norte do rio São Francisco, a Floresta Amazônica e os Brejos de Altitude. Os autores concluíram que os brejos mais a oeste estão mais relacionados à Floresta Amazônica e que esta, por sua vez, está mais relacionada à Mata Atlântica ao norte do rio São Franscisco. Entretanto os demais brejos mais ao leste tiveram pouca resolução nas análises. Assim, ficaram algumas perguntas em aberto: qual a relação entre os Brejos de Altitude e a Mata Atlântica ao norte do Rio São Francisco? Seriam os brejos uma unidade biogeográfica distinta (como defendido por Silva & Casteleti, 2005) ou constituem regiões da Mata Atlântica de isolamento recente? Essas questões até o momento permanecem sem resposta.
1.5. OS OPILIÕES DA MATA ATLÂNTICA.
O Brasil apresenta uma das maiores diversidade de opiliões do mundo, com cerca de 950 espécies. Dessas, cerca de 600 são exclusivas da Mata Atlântica (Bragagnolo & Pinto-da-Rocha, 2009a), a maioria delas ocorrendo na região Sudeste do Brasil.
As primeiras pesquisas com opiliões na Mata Atlântica brasileira datam do início do século XIX (Bragagnolo & Pinto-da-Rocha, 2009a). Grande parte dos estudos foi de cunho sistemático, tratando de descrever a opiliofauna até então desconhecida. Com esses esforços iniciais, foi sendo desenvolvido um entendimento sobre os padrões de distribuição das espécies. Uma das primeiras conclusões foi a constatação do alto grau de endemismo apresentado por esses aracnídeos (Pinto-da-Rocha et al., 2005).
Mais alguns fatores ainda prejudicavam as pesquisas biogeográficas com o grupo: o pouco entendimento sobre as relações de parentesco entre as diversas espécies e baixa quantidade de inventários com relação aos opiliões que ocupam as diferentes fisionomias da Mata Atlântica. Entretanto, essa situação vem mudando nos últimos anos, com a intensificação nas pesquisas com o grupo (Bragagnolo & Pinto-da-Rocha, 2009a).
2.
JUSTIFICATIVA
Tão importante quanto conhecer a fauna de uma área é reconhecer as suas relações históricas para se estabelecer uma unidade de conservação. Isto é, reconhecer quais organismos ocupam uma determinada e não outros e o porque de ocuparem essa área e não outra. Nesse aspecto é que entra a Biogeografia Histórica. O questionamento sobre a funcionalidade da Biogeografia Histórica no fornecimento de informações a fim de se definir uma área de conservação é amplamente discutido na literatura. Posadas et al. (2006) afirmam que a Biogeografia Histórica não apenas mostra as informações necessárias para conservação, mas também dispõe metodologias que podem ser diretamente aplicadas na determinação de prioridades para escolha dessas áreas.
3.
OBJETIVOS
• Comparar os resultados obtidos pelas diferentes metodologias utilizadas e avaliar a influência do tamanho das células sobre os resultados;
4.
MATERIAL E MÉTODOS
4.1. TÁXONS UTILIZADOS.
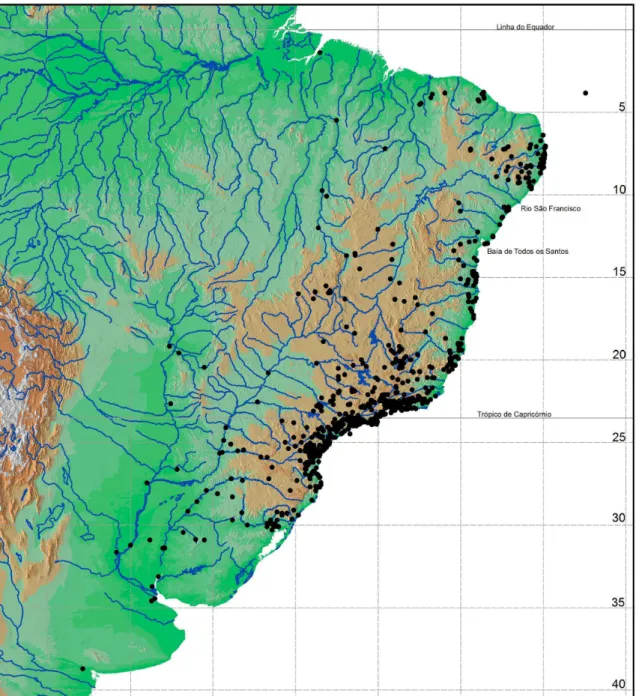
Foram incluídos nas análises táxons das seguintes famílias de opiliões da subordem Laniatores: Cosmetidae, Escadabiidae, Gonyleptidae, Minuidae, Kimulidae, Stygnidae e Zalmoxidae (Tabela 1). Um dos critérios utilizados para a inclusão ou exclusão dos táxons nas análises foi a situação taxonômica do táxon supra-específico. Foram incluídos os gêneros (ou subfamílias, no caso de Gonyleptidae) cuja revisão taxonômica tenha sido feita recentemente (a partir da década de 1990), ou aqueles cuja identificação não seja duvidosa, como por exemplo, gênero com espécies simpátricas de difícil reconhecimento das mesmas. Este critério é muito importante, dado o grau de confusão taxonômica que se encontra a maior parte dos taxa de opiliões neotropicais (p.ex., DaSilva & Gnaspini, 2009). Ao todo, foram utilizadas 1581 ocorrências de 224 espécies (Figura 1) que ocorrem por toda a Mata Atlântica, sendo que dessas, 18 ainda não foram descritas (Tabela 1).
Somente serão discutidos aqui os resultados que abrangem a Mata Atlântica nordestina, visto que as áreas encontradas ao sul e sudeste estão discutidas em DaSilva et al. (em preparação; manuscrito em anexo). Além do que a inclusão de todas as espécies visou avaliar o impacto do uso de um maior banco de dados sobre as análises. Os táxons a serem incluídos nas análises foram aqueles que possuíam distribuição em comum com ao menos outro táxon (simpatria parcial ou total). Logo, todos aqueles sem nenhuma sobreposição a outros táxons foram excluídos, pois não apresentam nenhum padrão de congruência biogeográfica, condição primária para a delimitação de áreas de endemismos e da Biogeografia Histórica em geral. Ainda foram também excluídos das análises os táxons com um único registro e/ou lote. Por terem registro e/ou lote único, esses taxa não apresentam informação suficiente para inferência de sua distribuição. Se algum taxon é registrado duas vezes, em momentos diferentes, ainda que na mesma localidade, já existe evidência de distribuição restrita.
4.2. COMPILAÇÃO DAS OCORRÊNCIAS.
vez numa mesma localidade, em momentos diferentes. Essas ocorrências foram obtidas através de três fontes: coletas, literatura disponível e visitas a museus.
4.2.1. Coletas.
De uma forma geral, a literatura é incipiente para os Opiliones no nordeste. Boa parte das espécies tem suas distribuições registradas apenas na localidade-tipo, sem novos registros (Kury, 2003). Assim sendo, tornou-se necessário a realização de coletas, a fim de se obter novos registros. As localidades amostradas foram escolhidas por serem apontadas como áreas importantes para a conservação da Mata Atlântica ao norte do rio São Francisco (Bencke et al., 2006). Foram realizadas coletas nos seguintes locais:
• Alagoas: Murici (Estação Ecológica de Murici – 9º 11’S 35º 53’W), São José da Laje e Ibateguara (Fragmentos da Usina Serra Grande – 8º 57’S 36º 06’W e 8º 59’S 35º 53’W, respectivamente);
• Ceará: Ubajara (Parque Nacional de Ubajara – 3º 42’S 40º 47’W), Baturité e Guaramiranga (Serra de Baturité – 4º 15’S 38º 58’W), Pacatuba e Maranguape (Serra da Aratanha – 3º 53’S 38º 44’W);
• Paraíba: Areia (Parque Estadual Mata do Pau-Ferro – 6º 57’S 35º 40’W), Cabedelo (Floresta Nacional de Cabedelo: Mata do Amém – 7º 03’S 34º 51’W), Mamanguape (Reserva Biológica Guaribas – 6º 45’S 35º 09’W) e João Pessoa (Jardim Botânico Benjamin Maranhão – Mata do Buraquinho – 07º 06’S 34º 52’W);
• Pernambuco: Bonito (Reserva Municipal Mata do Mucuri, 8º 59’S 36º 02’W), Caruaru (Parque Natural Municipal João Vasconcelos Sobrinho – Brejo dos Cavalos, 8º 16’ S 35º 58’ W), Igarassu (Refúgio Ecológico Charles Darwin e Mata da Usina São José – 7º 49’S 35º 00’W), Inajá (Reserva Biológica da Serra Negra – 8º 36’ S 38º 34’ W) e Jaqueira (Reserva Particular do Patrimônio Natural Frei Caneca – 8º 43’S 35º 54’W);
• Bahia: Maracás – 13º 40’S 40º 40’W.
coletam animais de hábitos e tamanhos diferentes. Os animais coletados foram fixados e conservados em álcool a 70% e depositados na coleção e aracnídeos da UFPB e no Museu de Zoologia da Universidade de São Paulo (MZSP).
4.2.2. Compilação dos dados da literatura.
Como parte do levantamento de ocorrências, também foi compilada a informação proveniente da literatura, principalmente de revisões taxonômicas e de listas de espécies (Revisões: Cosmetidae, Metavononoides – Ferreira, 2006; Gonyleptidae, Bourguyiinae – Yamaguti & Pinto-da-Rocha, 2009; Gonyleptidae, Caelopyginae – Pinto-da-Rocha, 2002; Gonyleptidae, Eusarcus – Hara & Pinto-da-Rocha, 2010; Gonyleptidae, Goniosomatinae – DaSilva & Gnaspini, 2009; Gonyleptidae, Hernandariinae – DaSilva & Pinto-da-Rocha, 2010; Gonyleptidae, Heteropachylinae – Mendes, 2011; Gonyleptidae, Longiperna – Pinto-da-Rocha & Bragagnolo, 2010a; Gonyleptidae, Progonyleptoidellinae – Kury, 2003; Gonyleptidae, Promitobates – Bragagnolo & Pinto-da-Rocha, 2012; Gonyleptidae, Roeweria – Bragagnolo & Pinto-da-Rocha, 2009b; Sodreaninae – Pinto-da-Rocha & Bragagnolo, 2010b; Stygnidae – Pinto-da-Rocha, 1997; Stygnidae, Pickeliana – Hara & Pinto-da-Rocha, 2008) . Essa parte do levantamento de ocorrências foi de suma importância por fornecer registros de lotes depositados em coleções que não foram visitadas no referido projeto.
4.2.3. Visita aos Museus.
A fim de aumentar a quantidade de ocorrências, também foi visitada a coleção do MNRJ (Museu Nacional da Universidade Federal do Rio de Janeiro) por dois motivos: por ser uma das coleções mais representativas em quantidade de lotes de opiliões e porque as demais coleções representativas para o táxon de estudo já foram visitadas recentemente e já haviam sido incluídas na planilha em trabalhos anteriores (DaSilva, comunicação pessoal; manuscrito anexo). Os lotes foram analisados e determinados até o nível de espécie, utilizando-se literatura especializada disponível (p. ex., Roewer, 1949; Pinto-da-Rocha, 1997; Kury, 2003; Ferreira, 2006; Pinto-da-Rocha et al., 2007; Mendes, 2011).
4.2.4. Georreferenciamento.
• Global Gazeteer: http://www.fallingrain.com/world/ • IBGE Cidades: http://www.ibge.gov.br/cidadesat/
Quando havia um detalhamento acerca da localidade, as coordenadas eram definidas com duas casas decimais. Entretanto, nem sempre isso foi possível, haja vista que algumas etiquetas não apresentavam tal detalhamento. Em tais casos, foi utilizada como coordenada o ponto médio do município, com somente uma casa decimal. Nessa etapa também foram excluídos os registros cuja localidade era duvidosa, imprecisa ou cuja etiqueta era pouco informativa e a localidade em questão era completamente fora da distribuição da espécie em questão.
4.3. ANÁLISES.
Para delimitar as áreas de endemismo, foram utilizados três métodos distintos: a Análise de Endemicidade (NDM), a Análise de Parcimônia de Endemismo (PAE) e Análise de Elementos Bióticos (Biotic Elements Analysis – BEA). Os três métodos tem por base de suas análises uma grade sobreposta a um mapa com as distribuições individuais de cada táxon. Para avaliar a influência do tamanho das células sobre os resultados foram usados três grades diferentes em cada método: 2° X 2°, 1° X 1° e 0,5° X 0,5°. Todas as grades sobrepostas ao mapa tinha origem em 6° a 52° S de latitude e 29° a 65° W de longitude. Além dos três métodos numéricos, também foi utilizada uma análise qualitativa utilizando uma série de critérios descritos na literatura e compilados por DaSilva (2008) e DaSilva & Pinto-da-Rocha (2011).
4.3.1. Análise de Endemicidade (NDM).
Onde:
• IE – Indice de Endemicidade do táxon “X”;
• p – número de células da área em que o táxon “X” está presente;
• i – número de células da área em que a presença do táxon “X” foi inferida, ou seja, células adjacentes as de presença real;
• t – número total de células da área;
• o – número de células adjacentes a área onde o táxon “X” está presente; • Fi – fator para presenças inferidas dentro da área (0,75 por padrão); • Fo – fator para presenças observadas fora da área (0,50 por padrão).
Ao final, cada área ganha um valor de endemicidade, que nada mais é que o somatório dos índices de endemicidade de cada táxon incluídos na área, ou seja, esse valor é influenciado por dois fatores: quantidade de espécies incluídas na área e pelo IE de cada espécie incluída. É considerada área de endemismo o grupo de células que tiverem dois pontos, ou seja, duas espécies perfeitamente congruentes com o grupo ou mais de duas espécies que somem dois pontos. A análise foi realizada utilizando os programas NDM/VNDM (Goloboff, 2005) utilizando a configuração padrão estabelecida pelo software.
4.3.2. Análise de Parcimônia de Endemicidade (PAE).
A Análise de Parcimônia de Endemicidade (PAE) foi proposta por Brian Rosen em 1988 e aperfeiçoado no mesmo ano por Rosen e Smith (Crisci et al., 2003). Este método possui algumas semelhanças com os cladísticos utilizados em análises filogenéticas, agrupando localidades (semelhante a táxons em cladística), pelos seus táxons compartilhados (semelhantes a caracteres em cladística). Essa metodologia aplica-se a táxons que compartilham diferentes localidades para inferir relacionamento entre as biotas amostradas nessas localidades, produzindo cladogramas de área diretamente das distribuições geográficas.
construção da matriz, foram usados os mapas gerados no NDM, sendo que somente as células com a ocorrência de pelo menos uma espécie foram contabilizadas. Para cada tamanho de célula foi confeccionada uma matriz para análise com a PAE, tendo cada matriz a seguinte quantidade de terminais: 301 (grade de 0,5º), 160 (grade de 1º) e 80 (grade de 2º). Como unidades operacionais geográficas (Operative Geografic Units - OGUs), foram utilizadas as células, que são reunidas pela presença compartilhada dos táxons (Morrone, 1994). Essa análise baseada em uma grade sobreposta ao mapa de distribuição das espécies é também chamada de Análise de Parcimônia de Endemismo baseada em células (Crisci et al., 2003). Essa metodologia difere um pouco do proposto inicialmente por Rosen (1988), pois, ao invés de assumir as localidades como OGUs, aqui as mesmas serão células com ocorrências das espécies (Morrone, 1994). A presença do táxon é tomada como o estado derivado (codificado como 1) e a ausência como primitivo (0). Analogamente à sistemática, o grupo externo é concebido hipoteticamente como sendo uma área primitiva, sem a presença de qualquer táxon (codificado como zero para todos).
Por esse método, serão considerados como área de endemismo os agrupamentos de células que apresentarem ao menos duas espécies endêmicas. A análise foi conduzida com os programas Nona 2.0 (Goloboff, 1999), em conjunto com o WinClada 1.00.08 (Nixon, 1999-2002). Foi utilizado o algoritmo heurístico TBR “tree bisection and reconection”. Além disso, também foi aplicado o comando “hold10000; mult*100; hold/10; mult*Max*” e, após a obtenção das árvores, foi aplicado o consenso estrito.
4.3.3. Análise de Elementos Bióticos (BEA).
A Análise de Elementos Bióticos (do inglês, Biotic Element Analysis – BEA) surgiu a partir do conceito de Hausdorf (2002), que discute as áreas de endemismo sob a perspectiva do modelo vicariante. Assim, segundo esse autor, as unidades básicas a serem usadas em estudos de biogeografia cladística deveriam ser “Elementos Bióticos”, que são grupos de táxons cujas distribuições são significativamente mais similares entre si que com qualquer outro grupo de táxons. Partindo desse princípio, Hausdorf & Hennig (2003) e Hennig & Hausdorf (2004) desenvolveram uma série de testes estatísticos.
como na PAE, a presença foi codificada como “1” e a ausência como “0”. O segundo arquivo de entrada consistia num arquivo com indicação de todas as células, com as respectivas células vizinhas. Igualmente a abordagem aqui utilizada na PAE, também foram construídas três matrizes com tamanhos de células diferentes para as análises na BEA.
Os testes foram aplicados utilizando o software R! (R Development Core Team, 2012) com o pacote Prabclus (Hennig, 2003). A primeira etapa da análise consiste num teste a fim de verificar se as distribuições são agrupadas ou não. Três especificações são necessárias: uma medida de distância, um teste estatístico e um modelo nulo (Hausdorf & Hennig, 2003). A medida de distância utilizada é a distância de Kulczynski, por ser mais adequada em casos de táxons de distribuição muito restrita. O teste estatístico é baseado nos dados de distribuições e o modelo nulo aplicado simula casos em que todas as diferenças encontradas nos dados possam ser atribuídas ao tamanho das distribuições dos táxons, ao número de táxons por célula e autocorrelação espacial das ocorrências dos táxons. Após essa etapa, é aplicado um Escalonamento Multidimensional Não-Métrico (NMDS) à matriz de distâncias de Kulczynski gerada pela etapa anterior. Por fim, um Agrupamento Baseado sobre um Modelo Gaussiano (MBGC) é aplicado sobre essa mesma matriz para identificar os elementos bióticos (mais detalhes em Hennig & Hausdorf, 2004).
4.3.4. Critérios combinados.
Além dos métodos numéricos, foram utilizados outros critérios para a delimitação das áreas de endemismo (DaSilva, 2008; DaSilva & Pinto-da-Rocha, 2011). Eles são usados de forma combinada e têm como fundamento tanto evidências bióticas quanto abióticas:
1 - Congruência da área de distribuição de pelo menos duas espécies: segundo esse critério, a delimitação de uma área de endemismo pode ser feita a partir da ocorrência de duas espécies em uma mesma área.
restritas a um NC, 2) endêmicas, mas com distribuições exclusivas, maiores que o NC, o que forma a região Máxima de Endemismo (RME) e 3) amplilocadas, ou seja, presentes em mais de um NC.
3 - Áreas de endemismo inclusivas: As espécies amplilocadas podem formar padrões de sobreposição mais amplos. Esses padrões não podem ser considerados como áreas de endemismo, pois podem incluir dois ou mais núcleos de congruência. Mas eles podem ser indicativos de relacionamento entre esses núcleos. Assim, servem como apoio para inferências históricas para explicar o surgimento dessas áreas menores, mas não são áreas de endemismo neste nível de análise.
4 - Exclusividade mútua: uma área de endemismo não deve apresentar qualquer sobreposição com outra área, pois sua biota é única. Assim, áreas com relacionamento duvidoso ou confuso podem ser resolvidas pelo acréscimo de novos dados ou pela fusão em uma única área.
5 - Padrões de endemismo por exclusão: podem surgir casos em que a distribuição de uma espécie pode apresentar pouca ou mesmo nenhuma simpatria com qualquer outra. Essa característica pode ser um indicativo da existência de uma área de endemismo à parte, que não se enquadrou em nenhum dos critérios anteriores, e que, portanto necessita de evidências para sua confirmação. Uma delas será a ocorrência de espécies-irmãs em áreas de endemismo próximas.
6 - Evidências geográficas independentes: além da distribuição das espécies (fator biótico), a delimitação de uma área de endemismo pode envolver outras evidências, como dados geológicos (fatores abióticos). Assim, espécies endêmicas sem sobreposição de distribuição, mas pertencentes à mesma unidade topográfica, podem ser indicativos de uma área de endemismo.
4.4. PROTOCOLO PARA DELIMITAÇÃO DAS ÁREAS DE ENDEMISMO. A seguir é apresentando o protocolo que foi utilizado aqui para a delimatação das áreas de endemismo após o uso dos métodos numéricos. Como a quantidade de áreas encontradas pelos métodos numéricos é muito grande, torna-se necessário o uso de um protocolo para a correta delimitação das áreas de endemismo, apresentando áreas que estejam em concordância com os Critétios Combinados. Esse protocolo encontra-se detalhado em DaSilva et al. (em preparação; texto em anexo).
(Morrone & Escalante, 2002; Deo & DeSalle, 2006). Isso foi muito importante devido ao banco de dados utilizado, que incluiu as ocorrências de opiliões de toda a Mata Atlântica, e já se sabe que as áreas de endemismo encontradas na Mata Atlântica do sul e sudeste apresentam tamanhos diferentes daquelas encontradas no nordeste (DaSilva & Pinto-da-Rocha, 2011). Além disso, diferentes tamanhos de células ajudam a reduzir vieses causados pela posição da grade, que pode considerar a presença de registros em uma célula e não em outra.
2) Nas análises do NDM, devido a grande quantidade de áreas encontradas pelo método, foi usada a opção de consenso estrito (Szumik et al., 2002) com o número mínimo de táxons compartilhados, ou seja, qualquer espécie encontrada em diferentes grupos de células foram unidas no consenso (foi usado 5% de similaridade em espécies). Isso é conceitualmente baseado sobre o Critério Combinado 4, que diz que as áreas de endemismo devem ser mutuamente exclusivas.
3) A delimitação das áreas inicia com os resultados da grade mais detalhada que representa o Núcleo de Congruência (NC) da área de endemismo, seguindo então para a grade menos detalhada. Grades menos detalhadas resultam em 1) áreas que não são encontradas quando se usa células menores, 2) a soma de área menores (que vai contra o critério combinado, 3) contradições com grupos de células menores, ou 4) artefatos causados pela cobertura da célula sobre diferentes áreas de endemismo. Diante disso, primeiramente devem ser considerados os resultados da grade mais detalhada, e posteriormente os resultados da grade menos detalhada.
4) Uma vez delimitado o NC das áreas de endemismo, o passo seguinte é classificar todas as espécies analisadas em três níveis de endemismo (critério combinado 2): 1) espécies restritas ao NC, indicada pelo métodos numéricos; 2) espécie ocorrendo somente em um NC, mas sem congruência distribucional suficiente (assim não detectada pelos métodos numéricos), formando uma Região Máxima de Endemismo (RME); e 3) espécies distribuídas em mais de um NC, ou amplilocadas.
5.
RESULTADOS
5.1. NDM.
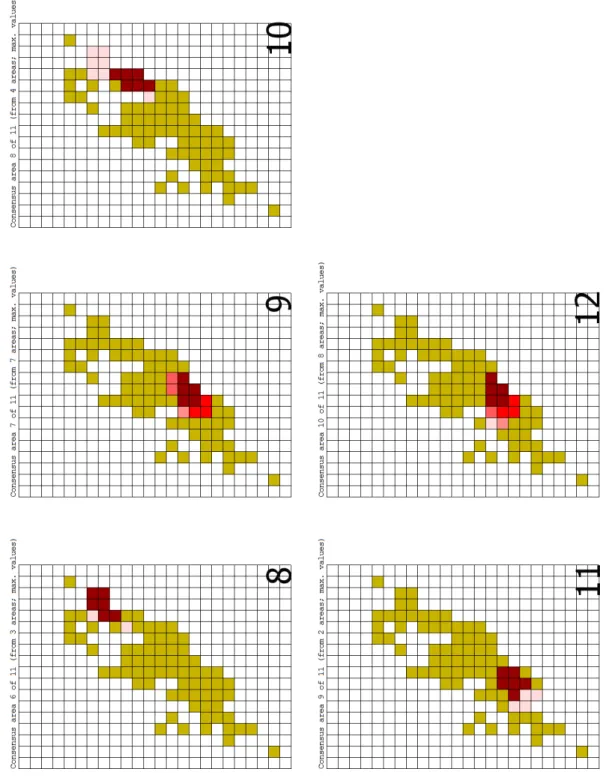
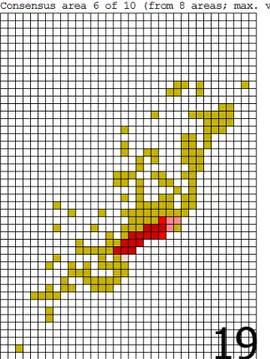
A análise com a grade de maior tamanho de células gerou um total de 42 áreas de endemismo. Várias dessas áreas não passam de redundâncias encontradas pelo programa. Ao ser gerado o consenso, a quantidade de áreas foi reduzida para 11. Dessas, somente quatro (Figuras 4, 7, 8 e 10) serão tratadas na discussão do trabalho, haja vista que as demais áreas estão relacionadas às florestas do sul de sudeste do Brasil. Entre essas quatro áreas, uma (que é resultado de um consenso de outras quatro áreas encontradas pela NDM) é composta por espécies que ocorrem na Mata Atlântica da Bahia (Figura 4). Já a segunda área (consenso de uma única área - Figura 7) localiza-se no interior do Ceará, abrangendo os Brejos de Altitude mais ao norte desse estado (Serra de Baturité e Serra da Ibiapaba). A terceira área, que é o consenso de três áreas encontradas pela NDM, corresponde à Mata Atlântica localizada no litoral dos estados de Alagoas, Pernambuco e Paraíba (Figura 8). Por fim, a quarta área (um consenso de quatro áreas) é composta por um misto de espécies que ocorrem na Mata Atlântica dos estados de Paraíba, Pernambuco, Alagoas, Sergipe e Bahia (Figura 10).
Com a grade intermédiária, de 1º, a quantidade de áreas encontradas pelo programa aumentou para 51. Após o consenso, a quantidade de áreas foi reduzida para 10, curiosamente menos do que na grade menos detalhada. Somente duas das áreas de consenso interessam ao trabalho (Figuras 13 e 21), as demais não serão discutidas pelas mesmas razões utilizadas na grade de 2°. A primeira área (que é um consenso de três áreas) é composta por espécies com distribuição na Mata Atlântica no litoral dos estados de Sergipe, Alagoas, Pernambuco e Paraíba (Figura 13). Já a segunda (resultante do consenso de 2 áreas), inclui espécies que ocorrem na Mata Atlântica da Bahia (Figura 21).
pelo programa, por ter a ocorrência de somente uma das espécies que compõem a área de endemismo. Já a segunda área apresenta somente três espécies que ocorrem na Bahia.
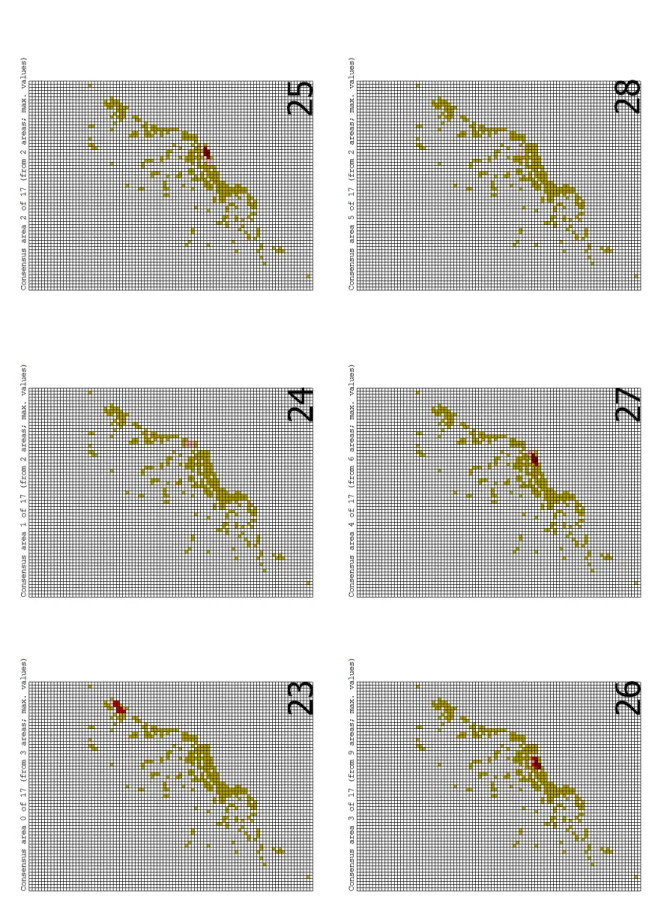
5.2. PAE.
O resultado da PAE com a matriz da maior grade gerou um total de 10.000 árvores. Essa quantidade poderia ter sido maior, já que a quantidade máxima imposta para buscar foi de 10.000 árvores. A grande quantidade de árvores encontrada já era esperada, haja vista existir uma grande quantidade de células sem resolução, o que ficou evidenciado quando foi pedido o consenso (L=397, Figura 40). Ao todo, sete conjuntos de células com, ao menos, duas espécies exclusivas foram encontrados pelo método (cada conjunto pode ser considerado uma área de endemismo). Dessas áreas, somente duas áreas interessam ao presente trabalho (Figuras 41 e 47). A primeira é composta por espécies que ocorrem na Mata Atlântica (Figura 41) da Bahia, enquanto que a segunda área é composta por espécies que ocorrem nas florestas dos estados de Alagoas, Paraíba e Pernambuco (Figura 47; mais detalhes ver Tabela 2).
Com a matriz de 1°, o resultado da busca por árvores foi semelhante, entretanto, ao ser gerado o consenso (L=503, Figura 48), assim como na análise da matriz de 2° foram encontradas sete áreas de endemismo (Figuras 49-55). Dessas, somente uma área (Figura 52) foi incluída na discussão do presente trabalho, já que inclui espécies que ocorrem na Mata Atlântica da Bahia (Tabela 2).
Para a matriz com as células mais detalhadas (0,5° X 0,5°), novamente o programa atingiu o máximo de 10.000 árvores. Ao ser gerado o consenso (L=648, Figura 56), constatou-se que o método identificou, assim como nas análises anteriores, sete áreas de endemismo (Figuras 57-63). Entretanto, as áreas encontradas com a matriz de 0,5° estão fora da discussão do presente trabalho, por serem áreas das regiões sudeste e sul da Mata Atlântica (Tabela 2).
5.3. BEA.
agrupamentos, três incluíram as espécies ocorrentes na Mata Atlântica nordestina (Tabela 2). Entretanto nenhum dos três incluiu somente as espécies dessa região.
O primeiro agrupamento (Figura 64) é composto por um misto de espécies que ocorrem na Mata Atlântica nordestina, em Minas Gerais e outras espécies com distribuição ao sul, um padrão geral sem qualquer sentido biogeográfico. O segundo agrupamento (Figura 68) é bem menor do ponto de vista espacial e sua composição apresenta espécies que ocorrem nas florestas entre o sul da Bahia e o norte do Espírito Santo. O terceiro agrupamento, um pouco maior que o anterior espacialmente, é composto por espécies que ocorrem ao sul da Bahia e norte do Espírito Santo e por algumas espécies da Mata Atlântica do sudeste (Figura 69), com distribuições disjuntas entre elas, ou seja, um agrupamento com espécies alopátricas.
Utilizando a matriz intermediária, o BEA conseguiu identificar nove agrupamentos de células significativos estatisticamente (T= 0,6485, p= 0,0297). Diferentemente do que ocorreu com a matriz de 2°, o valor de T encontrado ficou dentro do previsto pelo modelo nulo, que teve uma variação entre 0,6342 e 0,7530, com média de 0,6975. Dos nove agrupamentos identificados, quatro deles abarcam os dados do presente trabalho (Tabela 2).
O primeiro agrupamento (Figura 73) assemelha-se ao primeiro agrupamento encontrado na análise com a matriz de 2°. Assim como este, é composto por espécies que ocorrem no sudeste, em Minas Gerais e no nordeste, ou seja, sem o menor sentido biogeográfico. O segundo agrupamento (Figura 74) apresenta várias espécies que ocorrem ao norte do rio São Francisco, uma espécie com distribuição além dos limites da Mata Atlântica (Stygnus polyacanthus (Mello-Leitão, 1923)) e umas poucas espécies cuja distribuição vai até a Baía de Todos os Santos. O terceiro agrupamento (Figura 75) apresenta uma composição de espécies que ocorrem entre o sul da Bahia e o norte do Espirito Santo e uma espécie cujos pontos de ocorrência estão no Ceará e Ilha de Fernando de Noronha (Pseudopucrolia rugosa (Roewer, 1930)), ou seja, totalmente disjunta em relação ao seu próprio agrupamento. O quarto e último agrupamento apresenta uma grande quantidade de espécies que ocorrem na Mata Atlântica da Bahia (Figura 76), sendo que algumas poucas têm distribuição que chegam ao norte do Espirito Santo e uma espécie cuja distribuição se estende do norte do Espirito Santo até Alagoas (Pseudopucrolia mutica (Perty, 1833)).
significativos estatisticamente (T= 0,7868, p= 0,3168). O valor de T encontrado ficou dentro do esperado pelo modelo nulo cuja variação era de 0,7483 a 0,8410, com média de 0,7947. Isso significa que, embora o programa tenha encontrado alguns agrupamentos com uma composição de espécies coincidente com o encontrado por outros métodos, eles podem ser frutos do acaso (Tabela 2).
5.4. ASSOCIANDO OS MÉTODOS NUMÉRICOS AOS CRITÉRIOS
COMBINADOS.
Associando os resultados obtidos pelas análises numéricas com os critérios combinados e aplicando o protocolo proposto por DaSilva et al., (em preparação; manuscrito em anexo), fez-se então a delimitação das áreas de endemismo da Mata Atlântica nordestina. Foram encontrados agrupamentos semelhantes àqueles já descritos na literatura (DaSilva & Pinto-da-Rocha, 2011), sendo que três são referentes a Mata Atlântica nordestina (Figura 89). Duas destas áreas já haviam sido reconhecidas nos trabalhos prévios com opiliões (DaSilva, 2008; DaSilva & Pinto-da-Rocha, 2011).Segue abaixo uma descrição de cada uma das áreas, com os respectivos estudos que as mencionam.
• Área de Endemismo Pernambuco (PE):
Localizada entre os estados de Alagoas, Pernambuco, Paraíba, Rio Grande do Norte e Sergipe. O relevo caracteriza-se por ser formado por, desde uma planície costeira até os Brejos de Altitude, localizados no agreste pernambucano e paraibano, que estão inseridos no planalto da Borborema (Bonito, Caruaru e Bezerros, todos em Pernambuco). A pluviosidade varia entre 1000 a 1500 mm/ano (IBGE, 1985), graças aos ventos alísios vindos do sudeste. A cobertura vegetal inclui Floresta Ombrófila Densa, a Floresta Ombrófila Aberta e a Floresta Estacional Semidecidual (Tabarelli et al., 2005). O Núcleo de Congruência (NC) estende-se desde João Pessoa-PB até Ibateguara-AL no sentido norte-sul, e da costa até Caruaru-PE. A Região Máxima de Endemismo (RME) abrange a porção mais ao norte da Paraíba, estendendo-se a oeste para o Brejo de Altitude de Areia-PB, enquanto que seus limites ao sul chegam até o estado de Sergipe.
As espécies endêmicas a essa área são (mais detalhes na Tabela 3; NC = restrita ao Núcleo de Congruência):
Escadabius spinicoxa – NC Escadabius ventricalcaratus – NC Eusarcus sp – NC
Garanhunsa pectanalis – NC Iguarassua schubarti – NC Iguarassua sp – NC
Minuides rudicoxa – NC Pickeliana pickeli
Pirassunungoleptes analis Pseudopucrolia incerta Tegipiolus pachypus
• Área de Endemismo Bahia (BA):
Essa área caracteriza-se por ser uma pequena região formada pela planície costeira irregular ao sul da Bahia. Seus limites a oeste vão até os maciços cristalinos, que são mais altos e acidentados. A pluviosidade média anual oscila entre 1200 mm/ano até cerca de 1500 mm/ano (IBGE, 1985). A vegetação predominante é a Floresta Ombrófila Densa. O Núcleo de Congruência abrange as localidades de Camacan, Ilhéus, Itororó, Jussari e Una. A Região Máxima de Endemismo tem seus limites em Sergipe ao norte e, ao sul, próximo ao vale do Rio Doce.
As espécies endêmicas a essa área são (mais detalhes na Tabela 3; NC = restrita ao Núcleo de Congruência):
Baculigerus littoris Encheiridium sp – NC
Heteropachylus crassicalcanei Heteropachylus perachii
Heteropachylus ramphonotus – NC Jim benignus
Magnispina neptunus Metavononoides sp1
Mitogoniella unicornis – NC Pickeliana albimaculata Pickeliana capito
Protimesius junina
• Área de Endemismo Brejos Cearenses (BCE):
Trata-se de uma área de endemismo inédita para opiliões, caracterizada por incluir áreas de serras cristalinas e planaltos sedimentares florestados e isolados em meio à vegetação de Caatinga, com elevação variando entre 600 a 1000 metros de altitude (Borges-Nojosa & Caramaschi, 2003). A pluviosidade anual média varia em torno de 900 a 1400mm/ano, que é muito mais alta quando comparada a área de Caatinga circudante (cerca de 700mm/ano). A alta pluviosidade decorre das chuvas orográficas a barlavento, promovida pela ação dos ventos litorâneos (Mantovani, 2007). A vegetação predominante é a Floresta Estacional Semidecidual Montana (IBGE, 1985). O núcleo de congruência compreende as localidades de Baturité-CE, Guaramiranga-CE, Pacatuba-CE, Maranguape-CE e Ubajara-CE. A Região Máxima de Endemismo (RME) extende-se até o interior de Pernambuco e da Paraíba, abrangendo os Brejos de Altitude de Inajá-PE e Maturéia-PB.
As espécies endêmicas a essa área são (mais detalhes na Tabela 3): Escadabiidae sp
6.
DISCUSSÃO
6.1 AVALIAÇÃO DOS MÉTODOS: NDM, PAE e BEA. 6.1.1 O tamanho da grade.
Todos os métodos aqui utilizados (NDM, PAE e BEA) têm como base para análise a sobreposição de uma grade sobre um mapa com as distribuições, onde a unidade operacional é cada uma das células dessa grade. Mesmo sendo um fator tão importante, poucos autores mencionaram a influência causada pelo uso de diferentes tamanhos de células sobre os resultados (Posadas & Miranda-Esquivel, 1999; Morrone & Escalante, 2002; Hausdorf &Hennig, 2003; Mast & Nyffeler, 2003).
Posadas & Miranda-Esquivel (1999) mencionam que o tamanho da grade deve ser ajustado para o devido reconhecimento das áreas de endemismo pela PAE. Os autores sugerem a fusão de células ou o contrário, a depender da quantidade de dados disponíveis. Essa temática foi também discutida por Morrone & Escalante (2002). Os autores mencionaram que o uso de uma única grade pode trazer problemas, pois alguns registros podem ser divididos pelo tamanho da célula, ou que uma barreira geográfica (um rio, por exemplo) pode ser incluída numa única célula, o que poderia resultar em áreas resultantes de vieses do uso da grade.
pelo programa aumenta à medida que o detalhamento da grade diminui, ou seja, maior tamanho de células. Quanto ao aparecimento e desaparecimento de algumas áreas quando se altera o grau de detalhamento da grade, na análise atual, isso pode estar relacionado aos padrões de endemismo das espécies analisadas ou então ao banco de dados utilizado, que é bem detalhado.
Diferente dos métodos anteriores, o desempenho da BEA foi inversamente proporcional ao detalhamento da grade: quanto mais detalhada a grade, menos significativos os resultados. Com a grade menos detalhada, o método encontrou áreas extremamente significativas estatitiscamente, o que não foi visto quando se aumentou o detalhamento. Na grade mais detalhada, os resultados nem sequer foram significativos, e a quantidade de áreas encontradas caiu. Essa característica de perder a eficácia à medida que se aumenta o detalhamento também foi mencionada por Hausdorf & Hennig (2003). Para esses autores, essa queda de rendimento pode ter relação com o grau de refinamento dos dados. Grades muito detalhadas, oriundas de um banco de dados com um baixo esforço de coleta criariam lacunas nas distribuições das espécies que o programa interpretaria como ruído. Essa afirmativa, entretanto não funciona para o banco de dados aqui utilizado, que é amplo e bem detalhado.
6.1.2 Avaliando o desempenho dos métodos numéricos.
Os três métodos numéricos utilizados no presente trabalho (NDM, PAE e BEA) tiveram resultados, em alguns momentos, similares e, em outros, foram completamente diferentes. Embora com diferentes métodos de busca, os três métodos tem a mesma estratégia para encontrar as áreas de endemismo: procurar padrões de congruência entre as ocorrências das espécies. A partir desse ponto inicial, cada programa interpreta os dados de forma diferente.