SCIENTIFIC NOTE Identification, Geographical Distribution and Host Plants of Bemisia tabaci (Genn.) Biotypes (Homoptera: Aleyrodidae) in the State of Paraná, Brazil
Texto
Imagem
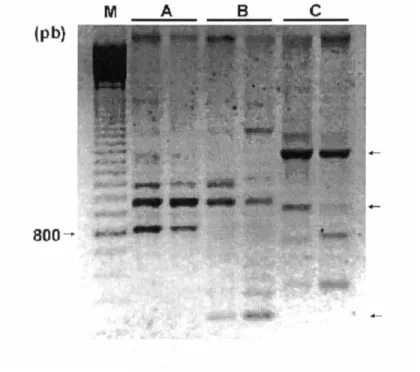
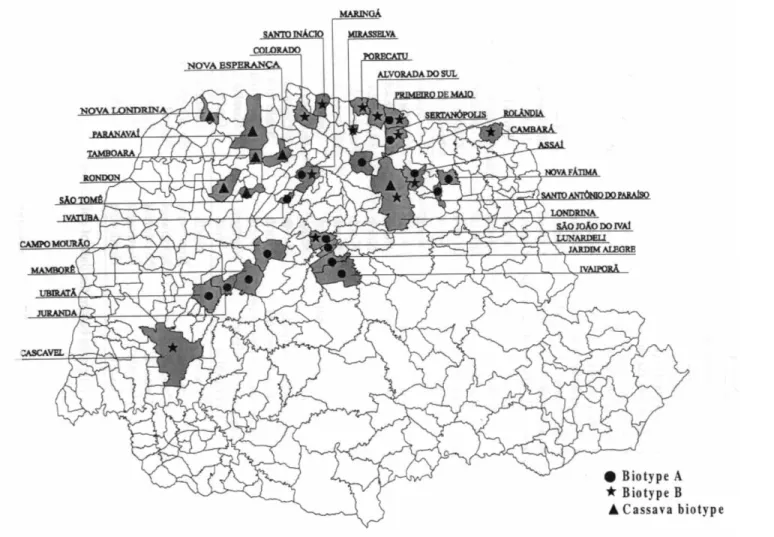
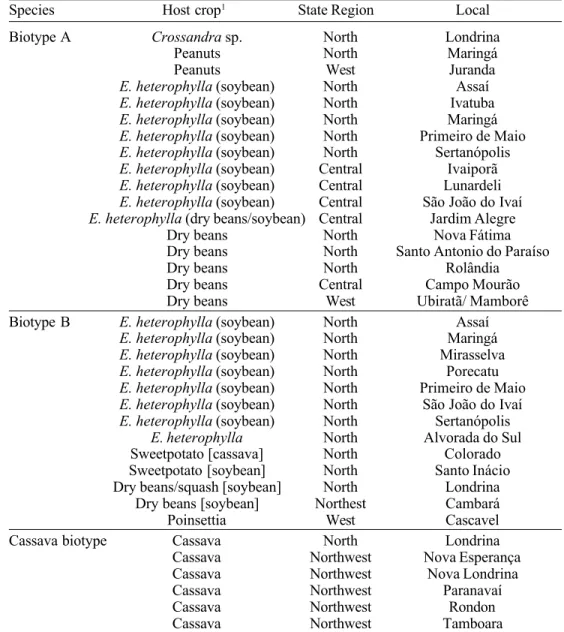
Documentos relacionados
Membranas obtidas por sinterização de partículas de PEUAPM foram desenvolvidas visando sua aplicação no tratamento de efluentes. As membranas foram sinterizadas em
Uma das explicações para a não utilização dos recursos do Fundo foi devido ao processo de reconstrução dos países europeus, e devido ao grande fluxo de capitais no
Neste trabalho o objetivo central foi a ampliação e adequação do procedimento e programa computacional baseado no programa comercial MSC.PATRAN, para a geração automática de modelos
FIGURA 5 - FLUTUAÇÃO POPULACIONAL DE Hypothenemus eruditus, Sampsonius dampfi e Xyleborus affinis (SCOLYTINAE, CURCULIONIDAE) COLETADOS COM ARMADILHAS ETANÓLICAS (25%)
Os modelos desenvolvidos por Kable & Jeffcry (19RO), Skilakakis (1981) c Milgroom & Fry (19RR), ('onfirmam o resultado obtido, visto que, quanto maior a cfiráda do
17 — Como fórmula de equilíbrio entre as várias soluções, pensamos que, para manter a igualdade real no processo, po- demos adotar a ampliação dos deveres do juiz nos termos do
b) um corpo-de-prova pertencente ao grupo SFC, isento de exposição à fumaça do cigarro. Assim, cada parte coronal configurou-se como unidade controle de si mesma. Após a
Neste contexto, este trabalho teve como objetivo caracterizar o ecossistema de inovação em bionegócios no qual está inserida a Rede Nordeste de Biotecnologia e
