Orie
Co-o
Júri: Pres Supe Voga Espí Dout da F Dout Agro Dout InstitPr
traç
Dis
ntadora: D
orientador:
: idente: Dou erior de Agr ais: Doutor rito Santo tora Maria T aculdade d tor José Ca onomia da U tora Elisabe tuto Superiorotecçã
ça-do-to
ssertação
Doutora Elis
Doutor Di
utor António ronomia da Dirceu Pr Teresa Fer e Ciências arlos Franc Universidad ete Tavare or de Agrono biológ
omateiro
Rosâ
o para obt
Engenha
sabete Tav
rceu Pratis
o Maria Ma Universida ratissoli, Pr reira Ramo da Universi co Santos S e Técnica d es Lacerda nomia da UnL
gica e m
o, Tuta a
ângela P
tenção do
aria Agro
vares Lace
ssoli
arques Mex de Técnica ofessor As os Nabais d idade de Lis Silva, Profe de Lisboa de Figuei niversidadeLisboa, 201
monitor
absolut
Payer
o Grau de
onómica
erda de Fig
xia, Profess de Lisboa ssociado da de Oliveira R sboa essor Auxili redo Olivei Técnica de10
rização
ta (Meyr
e Mestre
a
gueiredo O
sor Catedrá a Universid Rebelo, Pro ar do Instit ira, Profess e Lisboade
rick)
em
Oliveira
ático do Ins ade Feder ofessora Au tuto Superio sora Auxilia stituto ral do uxiliar or de ar doSão fúteis e cheias de erros as ciências que não nasceram da experimentação,
mãe de todo conhecimento.
ii À minha mãe, Carmem Peruch Payer, a minha grande impulsionadora, pelo amor, dedicação, carinho e incentivo para ultrapassar todos os obstáculos que até agora tive de
enfrentar. Obrigada por acreditar.
As minhas irmãs, Gissele e Ludmila, pelo apoio e amor com que me ajudaram mesmo distantes. Obrigada pelo vosso carinho.
À minha irmã Mônica e ao meu cunhado Rodolfo, pela ajuda e incentivo, sem eles eu não teria vindo para Portugal e chegado até aqui.
Aos meus sobrinhos, Juninho e Gugu, por trazerem momentos de muita alegria. Ao meu pai, ao meu cunhado e a minha avó (in memoriam), sempre lembrarei de vocês com
muito carinho e amor.
iii
AGRADECIMENTO
É com muita felicidade que chego ao fim desta etapa que marcou e muito estes últimos anos da minha vida.
Antes de mais quero agradecer à Cooperativa Nuestra Señora de Las Virtudes S.C.A., que através do estágio que realizei em 2009, permitiu o meu primeiro contacto com Tuta
absoluta.
À minha orientadora, Professora Elisabete Figueiredo, por quem tenho grande admiração e carinho, por permitir a realização desta tese, pelo valioso apoio, tempo, amizade e profissionalismo. Uma verdadeira mãe e um exemplo a seguir.
Ao meu co-orientador, Professor Dirceu Pratissoli, por quem tenho muita admiração, por aceitar este desafio, pela preciosa orientação e pelo trabalho desenvolvido na área de entomologia, no qual baseei grande parte desta tese.
Ao professor António Mexia, um verdadeiro motivador nato, pelo apoio logístico e pelas preciosas dicas.
Aos agricultores João Geada, Cidalina Silva, Ildefonso Fernandes e Fernanda Vieira Lopes, que disponibilizaram as parcelas para a realização do ensaio de campo.
Ao agricultor Sr. José Firmino, pela disponibilidade e simpatia.
À empresa AT&F, pelo fornecimento de cápsulas de feromona da marca Pheromon. À engenheira Sofia Rodrigues, pelo apoio e ajuda junto aos agricultores.
À Inês Silva, pela simpatia e ajuda na criação de Tuta absoluta.
À Sofia Melvill e à Rita Carvalho, pela amizade e pela hospitalidade em Valência, Espanha, no 1º Encontro Internacional sobre Tuta absoluta.
Ao engenheiro Juán A.Perez Rivero, que permitiu o meu contacto com a praga aqui estudada.
À engenheira Vera Zina, pelas dicas e pela foto da capa e a Doutora Elsa Borges da Silva pela ajuda nas fotos ao microscópio estereoscópico.
iv À técnica Luísa Valério, pela ajuda na rega dos tomateiros.
A todos os funcionários do Instituto Superior de Agronomia, pela amizade e simpatia. Ao Instituto Superior Técnico, pela aprendizagem inicial da minha carreira.
A todos os professores, os meus mestres, porque sem eles eu não teria a “bagagem” de suporte para este trabalho. Vou ter imensas saudades.
À Deus, o meu fundamento para continuar o caminho trilhado.
À engenheira e amiga para toda a vida, Nuria Borrego Pliego, pela amizade e apoio. Ao meu amigo espanhol, Manuel Nuñez Moreno, pela amizade e motivação.
À Deus, o meu fundamento e força para continuar o meu caminho.
À minha irmã portuguesa, Joana Martins, pelo apoio sempre presente, pela amizade, pelo carinho e pela dedicação. Obrigada pela ajuda preciosa na concretização desta tese, sem a qual não teria sido possível.
Ao meu grande amigo Diogo Zibaia, pelo apoio incondicional principalmente nas horas mais difíceis.
Ao Fernando Gaspar, pelo apoio e amizade ao longo dos últimos anos. À família Martins, pelo apoio incondicional e pela amizade.
Ao Frederico Cunha, pela amizade e ajuda preciosa nos pormenores deste trabalho.
À Emedina Silva, Ana Cadete, Carla Vieira, Carla Vinagre, Cármen Ribeiro, Sérgio Vitorino e Rita Messias, pelo carinho, apoio e amizade.
Ao Maxi Christl, pelo apoio, amizade, carinho e força.
Aos meus recentes amigos Júlia, Dário, Carol, Arlindo, Eduardo, Ricardo, Chico, Pépa, Pipa, Fonseca, Susana, Fred, Guerreiro, Rocheta, Helena Sanchéz, Maria, Catarina Antunes, Sara, Togi, Piriquito, Vitinho, David, Gui, André, Rute Rute e tantos outros, pelos momentos de distracção e de descontracção.
À minha família pela compreensão da minha ausência e pelo apoio sempre presente.
v
RESUMO
Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) para além dos estragos que provoca em
solanáceas, principalmente em tomate, tem demonstrado ter uma grande adaptabilidade, desde que se estabeleceu na Europa.
Com objectivo de utilizar espécies autóctones, como o parasitóide oófago Trichogramma
evanescens Westwood (Hymenoptera: Trichogrammatidae) e o ectoparasitóide Diglyphus isaea Walker (Hymenoptera: Eulophidae) contra T. absoluta, avaliaram-se as
potencialidades de ambos. Para o primeiro, estudou-se a taxa de parasitismo e a proporção de ovos em que o parasitóide atingiu o estado adulto e a taxa de viabilidade a diferentes temperaturas (22ºC, 24ºC e 25,5ºC). A taxa de parasitismo e de ovos com orifício de emergência maior a 22ºC e a taxa de viabilidade do tricograma maior a 24ºC. Para D. isaea, estudou-se a viabilidade desta praga como hospedeiro e a capacidade de predação por fêmeas adultas, utilizando-se dois tipos de ensaio, em placa de Petri e em gaiolas sobre plantas. D. isaea agiu como predador larvar, preferencialmente, larvas L1 e L2 (taxas de
predação: 17% em placas de Petri; 38% sobre planta).
Quatro marcas comerciais de feromona sexual de T. absoluta foram testadas em searas de tomate, no Ribatejo. O número de capturas de machos adultos não diferiu significativamente.
Palavras-chave: Tuta absoluta, Trichogramma evanescens, Diglyphus isaea, predação, parasitismo, feromonas
vi
ABSTRACT
Since its establishment in Europe, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) has shown to be very adaptable, and also to cause damage in the Solanaceae crops, mainly in tomatoes.
The egg parasitoid Trichogramma evanescens Westwood (Hymenoptera: Trichogrammatidae) and the ectoparasitoid Diglyphus isaea Walker (Hymenoptera: Eulophidae) were evaluated as potential biological control agents in order to use native species for T. absoluta control. For the egg parasitoid, the parasitism rate, the proportion of eggs with parasitoid emergence and the parasitoid viability (number of egg in the emergence/ number of parasitized egg) were evaluated at three different temperatures (22ºC, 24°C and 25.5ºC). The parasitism rate and the proportion of eggs with emergence were higher at 22ºC but viability was higher at 24ºC. For D. isaea, the parasitism rate and the larvae predation were evaluated in two scenarios: in Petri dishes and using tomato plants in a cage, registering larval parasitism and larval predation. The larval parasitoid acted as a predator, preferably on L1 and L2 T. absoluta larvae and predation rates were 17% and 38%,
respectively, in Petri dishes and tomato cages assays.
Four T. absoluta sexual pheromone trademarks were tested under field conditions. Male captures were not found to be statistically different.
Key words: Tuta absoluta, Trichogramma evanescens, Diglyphus isaea, predation, parasitism, pheromone
vii
EXTENDED ABSTRACT
The South American tomato moth, Tuta absoluta (Meyrick) affects many Solanaceae plants, and specially tomato. In Portugal and many other countries, it is considered now as a key pest.
In integrated pest management (IPM) of solanaceous crops, biological control of T. absoluta is very important. Monitoring of this pest population and monitoring the damage on plants is also very important to decide what are the most appropriate protection methods. In addition, it is essential to promote biological control by the natural enemies of this pest through the study of native species able to prey/parasitize it, such as the egg parasitoid Trichogramma
evanescens Westwood (Hymenoptera: Trichogrammatidae) and the ectoparasitoid Diglyphus isaea Walker (Hymenoptera: Eulophidae).
In this study, the potential of Trichogramma evanescens as parasitoid of T. absoluta eggs was evaluated at three different temperatures. Initially, the acceptance of T. absoluta eggs for T. evanescens oviposition was studied and it was found this parasitoid can complete its development in these eggs. In consequence, two assays were performed: one putting two female and another one putting only one T. evanescens female per tube and each containing 10 eggs of the host to assess the parasitism rate. The best results were obtained in the second assay, probably because in the latter the technique for introducing the female into the test tube was improved. Analyzing together the collected data obtained in these two assays, the highest parasitism rate and the highest proportion of eggs with adult emergence, respectively 50.8% and 43.2%, were obtained at 22 ° C. Furthermore, the viability (number of eggs with adult emergence/number of parasitized eggs) was higher at 24 °C (84%). Time to the first parasitoid adult emergences was also evaluated: 10, 11 and 12 days respectively, for 25, 5 ºC, 24 °C and 22 °C.
The autochthonous larval parasitoid Diglyphus isaea Walker was the other natural enemy studied. The possibility of the parasitoid parasitize/prey the T. absoluta larvae was evaluated in a preliminary assay. Only death by predation was observed. So, predation by D. isaea females was assessed in the other assays. The parasitism rate and the larvae predation were evaluated in two scenarios: in Petri dish and using tomato plants in a cage. In the first assay, leaflets with mines containing alive larvae and two D. isaea adults were placed in Petri dishes. The larvae were allowed to develop during seven days under controlled conditions. Proportion of predated larvae in the dishes was statistically different in the presence of D. isaea females but differences in predation rate between dishes with one
viii female (17%) and two females (16%) were not found. By removing the larvae present in the leaflets that were already too old for being predated, the rate increased a lot (30% and 26%, respectively). In the second assay, three plants infested with L1 and L2, T. absoluta larvae
and four adults of D. isaea were placed in each plastic cage. The predation rate was 38% and the percentage of larvae killed by unknown causes was 55%, being the total dead rate 93%. It is important to note that larvae presenting feeding bites showed an abnormal mine of irregular shape. Bites occurred mostly on L1 and L2 larvae and some few L3 recently molt. At
least in this case, bites probably occurred at the ecdisis.
The effectiveness of four sexual pheromone trademarks (dispenser +pheromone blend) for the South American tomato moth (Koppert, Suterra, Biobest and Pheromon) were also evaluated. For this, delta traps distant at least 150 m from each other were placed in four selected plots. The traps’ positions were changed every week, rotating them in the plot, and adult catches were weekly counted. No significant differences were found between the four pheromone trademarks regarding the adult catches in 25 week x plot. The Pheromon was better correlated with all the others trademarks.
ix
ÍNDICE
AGRADECIMENTO ... iii
RESUMO ... v
ABSTRACT ... vi
EXTENDED ABSTRACT ... vii
ÍNDICE DE FIGURAS ... xii
ÍNDICE DE QUADROS ... xiv
1 INTRODUÇÃO ... 1
2 REVISÃO BIBLIOGRÁFICA ... 3
2.1 CARACTERIZAÇÃO SUMÁRIA DE Tuta absoluta ... 3
2.1.1. Origem, sinonímia e taxonomia ... 3
2.1.2. Nomes comuns ... 3
2.1.3. Distribuição geográfica actual ... 3
2.1.4. Hospedeiros ... 5
2.1.5. Descrição, bioecologia e comportamento ... 5
2.1.6. Sintomas, estragos e prejuízos... 8
2.1.7. Estimativa do risco e meios de protecção ...12
2.2 CARACTERIZAÇÃO SUMÁRIA DE Trichogramma spp. ...19
2.2.1. Bioecologia ...19
2.3 CARACTERIZAÇÃO SUMÁRIA DE Diglyphus isaea ...22
2.3.1. Bioecologia ...22
2.3.2. Morfologia ...24
3 AVALIAÇÃO DE PARASITISMO POR Trichogramma evanescens ... 25
3.1 MATERIAL E MÉTODOS ...25
3.1.1. Criação e manutenção de Tuta absoluta (Meyrick, 1917) ...25
3.1.2. Aceitação por Trichogramma evanescens de Tuta absoluta como hospedeiro ....26
3.1.3. Avaliação da taxa de parasitismo ...26
x
3.2 RESULTADOS E DISCUSSÃO ...27
3.2.1 Aceitação de Tuta absoluta como hospedeiro ...27
3.2.2 Avaliação da taxa de parasitismo ...28
3.2.3. Determinação do tempo de desenvolvimento ...31
4 AVALIAÇÃO DE PARASITISMO E PREDAÇÃO POR Diglyphus isaea ... 32
4.1 MATERIAL E MÉTODOS ...32
4.1.1. Aceitação por Diglyphus isaea de Tuta absoluta como hospedeiro ...32
4.1.2. Avaliação do parasitismo/predação em placas de Petri ...32
4.1.3. Avaliação de parasitismo/predação em planta de tomateiro ...33
4.2 RESULTADOS E DISCUSSÃO ...34
4.2.1. Aceitação por Diglyphus isaea de Tuta absoluta como hospedeiro ...34
4.2.2. Avaliação do parasitismo/ predação em placas de Petri ...34
4.2.3. Avaliação do Parasitismo/ Predação em plantas de tomateiro...37
5 COMPARAÇÃO DA EFICÁCIA DE MARCAS COMERCIAIS DE FEROMONA SEXUAL DE TRAÇA-DO-TOMATEIRO ... 39 5.1 MATERIAL E MÉTODOS ...39 5.2 RESULTADOS E DISCUSSÃO ...40 6 CONCLUSÕES ... 45 7 REFERÊNCIAS BIBLIOGRÁFICAS ... 47 ANEXOS ... 56
Anexo 1 - Dados laboratoriais relativos a Trichogramma evanescens. ...56
Anexo 2 – Análise estatística a normalidade, rank do Teste de Kruskal-Wallis e estatística do teste de Mann-Whitney para a taxa de parasitismo, proporção de ovos com orifício de emergência e viabilidade de Trichogramma evanescens nos ensaios I, II e I + II com a transformação angular da percentagem...59
Anexo 3 – Resultados observados no Ensaio I – Avaliação do parasitismo/predação de Diglyphus isaea em placas de Petri. ...64
Anexo 4 – Resultados observados no Ensaio II – Avaliação do parasitismo/ predação de Diglyphus isaea em gaiolas. ...66
Anexo 5 – Análise estatística a normalidade, posição média do Teste de Kruskal-Wallis e do Teste de Mann-Whitney para a predação, larvas mortas por causa desconhecida e
xi larvas potencialmente atacáveis referentes ao Ensaio I - Avaliação da predação/parasitismo de Diglyphus isaea em placas de Petri com a transformação angular da proporção. ...67 Anexo 6 – Análise estatística a normalidade e ao teste t-student para a predação referentes ao Ensaio II - Avaliação do parasitismo/ predação de Diglyphus isaea em plantas de tomateiro com a transformação angular da proporção de larvas predadas. ...70 Anexo 7 – Representação esquemática de cada uma das parcelas, A, B, C e D, com as respectivas posições das armadilhas do tipo delta – ensaio de campo de comparação de feromonas sexuais de Tuta absoluta ...71 Anexo 8 – Dados e observações recolhidos no ensaio de campo de comparação de marcas comerciais de feromonas ...72 Anexo 9 – Análise estatística do ensaio de campo realizado entre os dias 26 de Julho e 28 de Setembro de 2010 para comparaçºao de feromonas de Tuta absoluta: testes de normalidade, teste de Friedman e teste t-student. ...75 Anexo 10 – Genitália: Identificação de Tuta absoluta em folhas de Solanum tuberosum e
xii
ÍNDICE DE FIGURAS
Figura 1. Mapa de distribuição geográfica de Tuta absoluta na Europa, Médio Oriente e Norte de África (a presença num país/região é indicada no mapa) (Anónimo, 2009b). ... 5 Figura 2. Ciclo de vida de Tuta absoluta (originais da autora excepto ovo de Anónimo (2009a)). ... 7 Figura 3. Adultos de Tuta absoluta realizando a cópula (original da autora). ... 8 Figura 4. Minas de Liriomyza sp. (galerias, setas azuis) e de Tuta absoluta (setas a vermelho) em folhas de tomate. Observam-se também estragos no caule (Autoria: Elisabete Figueiredo, 2010)... 10 Figura 5. Folhas de Solanum nigrum com sintomas de Tuta absoluta no campo de tomate (originais da autora). ... 10 Figura 6. Larva e estragos de Tuta absoluta em folhas de batateira, no distrito de Setúbal (originais de Eng. Paula Santos, AT& F.) ... 11 Figura 7. Estragos causados por Tuta absoluta em plântula de pimento (Capsicum annuum) à esquerda e em beringela (Solanum melongena) à direita (originais de EPPO Gallery 2010). ... 11 Figura 8. Observação de larvas, galerias em folhas e adulto de Tuta absoluta em tomate de estufa na região Oeste (originais da autora). ... 11 Figura 9. Tomateiros ao ar livre atacados pela traça-do-tomateiro (originais da autora). ... 11 Figura 10. Larva de Tuta absoluta e estragos em frutos verdes e maduros no campo de tomate. Observam-se os orifícios de entrada e saída, principalmente junto ao pedúnculo (originais da autora). ... 12 Figura 11. Fruto verde com duas pupas de Tuta absoluta no pedúnculo. Em ambos os frutos são visíveis os estragos (originais da autora). ... 12 Figura 12. Estragos em frutos provocados pela alimentação larvar da traça-do-tomateiro (originais da autora). ... 12 Figura 13. Armadilha de água comercial (à esquerda) e artesanal (à direita) com feromona sexual para a captura da traça-do-tomateiro (originais da autora). ... 15 Figura 14. Adulto fêmea de Trichogramma evanescens (SESIL, s/data). ... 19 Figura 15. Desenvolvimento da lagarta do hospedeiro versus desenvolvimento de Trichogramma spp. (Knutson, 1993). ... 21 Figura 16. Diferenças entre as antenas de Trichogramma spp. macho, com muitos pelos longos e a antena da fêmea com poucos pelos curtos (Knutson, 1993). ... 21 Figura 17. Ciclo de vida de Diglyphus isaea: 1. Adulto, 2- Ovo, 3- Larva e 4- Pupa (Malais & Ravensberg, 1996) 23 Figura 18. Duas pupas de Diglyphus isaea dentro da mina da larva mineira (Bio-Bee, 2007)... 23 Figura 19. Adultos de Diglyphus isaea (foto à direita da autoria de A. Mil Homens; à esquerda original da autora). ... 24 Figura 20. Visão geral do suporte com gaiolas de acrílico com larvas, pupas e adultos de Tuta absoluta (originais da autora). ... 25 Figura 21. Dois tubos com cartão com 9 e 7 ovos parasitados por Trichogramma evanescens (Ensaio II), dos 10 inicialmente colocados (originais da autora). ... 30
xiii Figura 22. Ovos com orifício de emergência e fêmea de Trichogramma evanescens (originais da autora). ... 31 Figura 23. Seis placas de Petri com folíolos de tomateiro e adultos de Diglyphus isaea do ensaio de avaliação de parasitismo/predação (originais da autora). ... 33 Figura 24. Larvas predadas por Diglyphus isaea. As setas a vermelho indicam o local da picada de alimentação (original da autora). ... 36 Figura 25. Diferenças observadas no ensaio de predação em tomateiro entre os estragos causados por: A - larva predada e B - larva saudável (original da autora). ... 37 Figura 26. Armadilha do tipo delta na parcela A (à esquerda) e cartão impregnado com cola e difusor (à direita) (originais da autora). ... 39 Figura 27. Evolução de capturas da traça-do-tomateiro na parcela A, com as quatro marcas de feromonas. .... 41 Figura 28. Evolução de capturas da traça-do-tomateiro na parcela B, com as quatro marcas de feromonas. .... 42 Figura 29. Evolução de capturas da traça-do-tomateiro na parcela C, com as quatro marcas de feromonas. .... 42 Figura 30. Evolução de capturas da traça-do-tomateiro na parcela C, com as quatro marcas de feromonas. .... 42 Figura 31: Genitália do macho de Tuta absoluta (original de EPPO Gallery, 2010). ... 77
xiv
ÍNDICE DE QUADROS
Quadro 1. Posição taxonómica (Karsholt et al., 2009). ... 3
Quadro 2. Zero de desenvolvimento estimado para cada um dos estados de desenvolvimento de Tuta absoluta (Barrientos et al., 1998). ... 8
Quadro 3. Inimigos naturais mais importantes de Tuta absoluta na América Latina. ... 17
Quadro 4. Taxa de parasitismo, proporção de ovos com orifício de emergência e viabilidade de Trichogramma evanescens, referentes ao Ensaio I, em diferentes temperaturas, 70 ± 10 % de HR e fotoperíodo 14h L: 10h D. 29 Quadro 5. Taxa de parasitismo, proporção de ovos com orifício de emergência e viabilidade de Trichogramma evanescens, referente ao Ensaio II, em diferentes temperaturas, 70 ± 10 % de HR e fotoperíodo 14h L: 10h D. . 29
Quadro 6. Taxa de parasitismo, proporção de ovos com orifício de emergência e viabilidade de Trichogramma evanescens, referente ao Ensaio I e II em conjunto, em diferentes temperaturas, 70 ± 10 % de HR e fotoperíodo 14h L: 10 h D. ... 30
Quadro 7. Resultados da análise de variância não paramétrica (Teste de Kruskal-Wallis) para o Ensaio I. ... 30
Quadro 8. Resultados da análise de variância não paramétrica (Teste de Kruskal-Wallis) para o Ensaio II. ... 31
Quadro 9. Resultados da análise de variância não paramétrica (Teste de Kruskal-Wallis) para o Ensaio I+ II. ... 31
Quadro 10. Resultados do ensaio preliminar de aceitação de Tuta absoluta como hospedeiro por Diglyphus isaea. ... 34
Quadro 11. Percentagem de predação, larvas mortas por causa desconhecida e larvas potencialmente atacáveis por Diglyphus isaea, referente ao ensaio em placa de Petri. ... 35
Quadro 12. Análise estatística dos resultados de predação do ensaio de avaliação em placa de Petri utilizando a ANOVA não paramétrica, teste de Kruskal-Wallis a, b. ... 36
Quadro 13: Comparação do efeito de largada, em placa de Petri, de uma e de duas fêmeas, na predação por Diglyphus isaea, utilizando o teste de Mann-Whitney. ... 36
Quadro 14. Taxa de predação, de larvas saudáveis, de larvas mortas por causa desconhecida e de mortalidade atribuível à predação por Diglyphus isaea, em plantas de tomateiro. ... 37
Quadro 15. Caracterização de cada um das parcelas utilizadas na monitorização de Tuta absoluta. ... 40
Quadro 16. Feromonas utilizadas no ensaio de campo e respectivos lotes. ... 40
Quadro 17. Captura média para cada uma das feromonas. ... 43
Quadro 18. Correlação para as diferentes marcas de feromonas. ... 43
Quadro 19. Contagem dos ovos de Ephestia kuehniella parasitados e não parasitados por Trichogramma evanescens nos cartões fornecidos pela empresa Andermatt. ... 56
Quadro 20. Procedimentos e observações realizadas do ensaio preliminar, para avaliar a capacidade de aceitação por Trichogramma evanescens de ovos de Tuta absoluta. ... 56
Quadro 21. Resultado das observações efectuadas no ensaio I. ... 57
Quadro 22. Resultado das observações efectuadas no ensaio II. ... 58
Quadro 23. Teste a normalidade para o Ensaio I. ... 59
xv
Quadro 25. Teste a normalidade para o Ensaio I + II. ... 60
Quadro 26. Posição média do Ensaio I. ... 60
Quadro 27. Posição média do Ensaio II. ... 61
Quadro 28. Posição média do Ensaio I + II. ... 61
Quadro 29. Estatística do teste de Mann-Whitney para a taxa de ovos com orifício de emergência as temperaturas de 22 ºC e 24 ºC. ... 62
Quadro 30. Estatística do teste de Mann-Whitney para a taxa de ovos com orifício de emergência as temperaturas de 22 ºC e 25,5 ºC. ... 62
Quadro 31. Estatística do teste de Mann-Whitney para a taxa de ovos com orifício de emergência as temperaturas de 24 ºC e 25,5 ºC. ... 62
Quadro 32. Estatística do teste de Mann-Whitney para a taxa de viabilidade as temperaturas de 22 ºC e 24 ºC. ... 62
Quadro 33. Estatística do teste de Mann-Whitney para a taxa de viabilidade as temperaturas de 22 ºC e 25,5 ºC. ... 63
Quadro 34. Estatística do teste de Mann-Whitney para a taxa de viabilidade as temperaturas de 24 ºC e 25,5 ºC. ... 63
Quadro 35. Contabilização do número de larvas e pupas e número de fêmeas em cada uma das placas de Petri ... 64
Quadro 36. Teste a normalidade para larvas predadas, mortas por causa desconhecida e potencialmente atacáveis (após transformação angular) ... 67
Quadro 37. Posição média do Teste de Kruskal-Wallis de larvas predadas, mortas por causa desconhecida e potencialmente atacáveis. ... 68
Quadro 38. Posição média do Teste de Mann-Whitney de larvas predadas, mortas por causa desconhecida e potencialmente atacáveis. ... 68
Quadro 39: Estatística do Teste de Mann-Whitney de larvas predadas, mortas por causa desconhecida e potencialmente atacáveis. ... 69
Quadro 40. Teste a normalidade b,c para as larvas predadas após transformação angular. ... 70
Quadro 41. Estatística descritiva a após transformação angular. ... 70
Quadro 42. Teste t-student após transformação angular... 70
Quadro 43. Dados e observações recolhidas em quatro parcelas, durante o ensaio de campo de comparação de feromonas. ... 72
Quadro 44. Teste a normalidade para cada uma das feromonas estudadas no ensaio de campo. ... 75
Quadro 45. Posição média e estatística do teste de Friedman para as feromonas estudadas. ... 75
Quadro 46. Estatística descritiva relativa ao teste t-student. ... 76
A cultura do tomate em Portugal tem grande importância, sendo o tomate de indústria a principal produção horto-industrial (MADRP/GPP, 2010). De acordo com o Instituto Nacional de Estatística (INE, 2010), a produção de tomate atingiu no ano de 2009 as 1346.702 toneladas. Todavia, a cultura do tomate é atacada por inúmeras pragas da qual se destaca Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) (Pratissoli & Parra, 2000). Desde que foi detectada, em 2006, na província de Castellón, em Espanha, esta praga, com o nome vulgar de traça-do-tomateiro, expandiu-se rapidamente, num curto espaço de tempo, por toda a Europa, Norte de África e Médio Oriente (EPPO, 2005; Hernández et al., 2008; Anónimo, 2009b, Arnó & Gabarra, 2010; Cabello, 2010; Llopis et
al., 2010). Actualmente, é considerada uma praga-chave para a cultura do tomate, em
Espanha, apesar de também atacar outras solanáceas (Camacho et al., 2009). As larvas da traça-do-tomateiro podem causar estragos em todas as fases de crescimento da planta, produzindo minas nas folhas, perfurando caules, gomo apical e frutos verdes e maduros (Lietti et al., 2005).
Para o combate de T. absoluta é necessária uma estratégia de protecção integrada baseada na combinação de diferentes estratégias de protecção, nomeadamente culturais, biotécnicas, químicas e biológicas (Monserrat, 2009). No entanto, muitos agricultores optam apenas pela aplicação de pesticidas, causando problemas de resistência (Pratissoli & Parra, 2000). Além disso, é de frisar que, em Portugal, apenas uma substância activa, o indoxacarbe, está autorizada para a traça-do-tomateiro (DGADR, 2010). Considerando estes aspectos, torna-se importante a possibilidade de utilização de parasitóides oófagos do género Trichogramma, pois trata-se de “vespas” que parasitam um grande número de hospedeiros e são altamente eficientes (Pratissoli & Parra, 2001). Por exemplo, Trichogramma evanescens é uma espécie que existe no Ribatejo e que limita naturalmente noctuídeos em searas de tomate (Silva et al., 1999; Amaro & Mexia, 2006), para além de ser usado como agente de protecção biológica em tratamentos biológicos contra várias espécies de lepidópteros (Amaro, 2003). Cabello, (2010) cita a importância de se trabalhar com os possíveis inimigos naturais autóctones que se podem adaptar a esta nova presa/hospedeiro. Mantendo esta linha de pensamento, a avaliação da possibilidade de Diglyphus isaea Walker (Hymenoptera: Eulophidae) funcionar como provável auxiliar no combate a T. absoluta é uma outra forma de obter um valor acrescentado, já que este ectoparasitóide é utilizado em Portugal em conservação e para o tratamento biológico contra Liriomyza spp. (Amaro, 2003). Por outro lado, a utilização de armadilhas com feromona sexual é uma forma de detectar atempadamente a presença de T. absoluta e avaliar a densidade das
2 populações (Anónimo, 2009b), servindo para monitorizar e realizar capturas em massa desta traça.
Desta forma, o objectivo deste trabalho foi o desenvolvimento de estudos biológicos utilizando as espécies autóctones, Trichogramma evanescens e Diglyphus isaea, visando uma possível aplicação dos mesmos em tratamento biológico contra a traça-do-tomateiro. A outra vertente do presente trabalho foi o estudo da eficácia de feromonas de Tuta absoluta.
Em síntese, consideram-se os seguintes objectivos específicos: - em ensaios laboratoriais:
(1) determinar a aptidão de Tuta absoluta como hospedeiro para o parasitóide oófago Trichogramma evanescens através da avaliação da taxa de parasitismo e da proporção de ovos com emergência de adulto (ovos com orifício de emergência) e viabilidade do adulto e avaliar o tempo de desenvolvimento a diferentes temperaturas;
(2) avaliar a possibilidade de Diglyphus isaea parasitar/predar larvas de Tuta
absoluta;
- em ensaio de campo:
(1) Avaliar a eficácia de quatro marcas comerciais de feromona sexual de Tuta
absoluta em campos de tomate de indústria, para monitorização da população
com vista à sua utilização na estimativa do risco.
Tendo em consideração o que foi dito anteriormente, este trabalho foi estruturado em seis capítulos. Após a introdução, é apresentada a revisão bibliográfica subdividida em três subcapítulos. No primeiro destes foi feita uma descrição sumária de T. absoluta, no segundo e terceiro subcapítulos referiram-se alguns aspectos relativos a T. evanescens e D. isaea, respectivamente. No terceiro capítulo fez-se a avaliação do parasitismo por
T. evanescens, descrevendo o material e métodos, resultados obtidos e sua discussão.
No quarto capítulo, avaliou-se o parasitismo e predação de larvas de T. absoluta por D.
isaea, descrevendo também a metodologia, os resultados e a sua discussão. No quinto
capítulo estudou-se a eficácia de feromonas, em condições de campo. Neste, descreve-se o material e método e faz-descreve-se uma análidescreve-se dos resultados. Optou-descreve-se por esta divisão, para uma melhor organização do trabalho e, consequentemente, mais fácil leitura. No último capítulo são apresentadas as conclusões deste trabalho.
3
2.1 CARACTERIZAÇÃO SUMÁRIA DE Tuta absoluta
2.1.1. Origem, sinonímia e taxonomia
Tuta absoluta (Meyrick, 1917) (Lepidoptera: Gelechiidae) é um microlepidóptero
originário da América do Sul. Esta espécie foi descrita pela primeira vez em Huancayo, no Peru, em 1917, por Meyrick, como Phthorimaea absoluta (Hernández et al., 2008). Para além desta sinonímia, também foi denominada Gnorimoschema absoluta Clarke, 1962, Scrobipalpula absoluta Povolny, 1964, Scrobipalpuloides absoluta Povolny, 1987 até ser incluída no género Tuta, em 1994 (EPPO, 2005; Hernández et al., 2008).
A posição sistemática da espécie T. absoluta encontra-se descrita no Quadro 1.
Quadro 1. Posição taxonómica (Karsholt et al., 2009).
Taxa Nome Filo Arthropoda Classe Insecta Ordem Lepidoptera Superfamília Gelechioidae Família Gelechiidae Subfamília Gelechiinae Tribo Gnorimoschemini
Género Tuta Kieffer & Jörgensen, 1910
Espécie T. absoluta (Meyrick, 1917)
2.1.2. Nomes comuns
As denominações vulgarmente atribuídas a T. absoluta estão relacionadas com a língua de cada país. Em português é conhecida como a traça-do-tomateiro (EPPO, 2005); em francês, “mineuse de la tomate” (Ramel & Oudard, 2008); em inglês, “tomato borer”, “South American tomato moth”, “tomato leaf miner” or “South American tomato pinworm” (EPPO, 2005); e, em espanhol como “polilla del tomate” (Monserrat, 2009), “polilla perforadora”, “cogollero del tomate”, “gusano minador del tomate”, “minador de hojas” y “tallos de la papa” (EPPO, 2005).
2.1.3. Distribuição geográfica actual
Desde 1960 T. absoluta é considerada uma praga-chave da cultura de tomate em vários países da América do Sul (Lietti et al., 2005). Esta praga encontra-se na Argentina, Bolívia, Brasil, Chile, Colômbia, Equador, Paraguai, Peru, Uruguai e Venezuela. No
4 entanto, não aparece acima dos 1000 m de altitude (Korycinska & Moran, 2009), uma vez que neste caso, a temperatura é um factor limitante para a sua sobrevivência (Hernández et al., 2008). Também é curioso notar que T. absoluta ainda não foi observada na América Central nem na América do Norte (Garcia-Marí & Vercher, 2010).
Na Argentina, esta praga foi, pela primeira vez, citada no ano de 1969, na província de Mendoza, tendo como provável foco de entrada tomates importados do Chile no ano de 1964. Em 1968, foi detectada no Uruguai (Caffarini et al., 1999). No Brasil, este insecto foi referenciado pela primeira vez, em 1980, no Estado de São Paulo (Gonçalves-Gervásio et al., 1999).
No continente asiático existe um registo antigo de T. absoluta atacando Solanum
lyratum Thunberg no Japão, mas não existem registos recentes (EPPO, 2005).
T. absoluta entrou na Europa via Espanha, onde foi detectada pela primeira vez no final
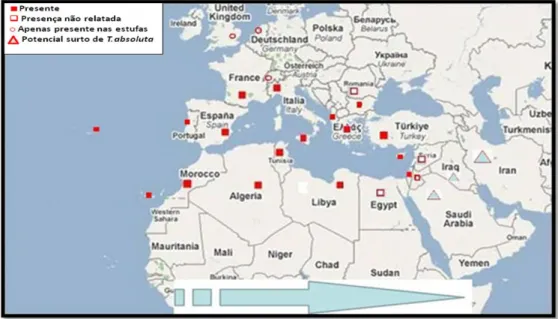
de 2006, na Comunidade Valenciana em Castellón (Hernández et al., 2008; Ramel & Oudard, 2008). Desde então, e num curto espaço de tempo e devido a incrível adaptabilidade, esta praga tem-se espalhado por diversos países (Figura 1). Durante os últimos dois anos, T. absoluta foi detectada em Itália, França, Malta, Marrocos, Argélia, Tunísia, Líbia, Albânia, Reino Unido, Grécia, Suíça e Portugal (EPPO, 2005; Hernández
et al., 2008; Anónimo, 2009b). De acordo com National Plant Protection Organization
(NPPO), esta praga foi detectada no Reino Unido, em Março de 2009, numa central de embalamento, vinda através de tomate importado de Espanha e Itália (EPPO, 2009). Em Portugal, a traça-do-tomateiro foi assinalada no Algarve, em Maio de 2009, em estufas na zona de Faro e, em Julho, de 2009, na região Oeste, em estufas na zona da Silveira e A-dos-Cunhados (Anónimo, 2009b; Figueiredo et al., 2010). No Verão de 2009 foi detectada em armadilhas iscadas com feromona sexual na Lezíria do Ribatejo e, no Outono de 2009, verificou-se a presença em estufas do Entre-Douro e Minho (Figueiredo, com. pes.). Em Setembro deste mesmo ano, a praga foi identificada na Alemanha, em Novembro em Chipre, em Dezembro na Bulgária (Anónimo, 2009b) e Rússia (Garcia-Marí & Vercher, 2010).
Recentemente, T. absoluta foi detectada na Síria, Turquia, Israel, Iraque e Arábia Saudita (Anónimo, 2009b).
5
Figura 1. Mapa de distribuição geográfica de Tuta absoluta na Europa, Médio Oriente e Norte de África (a presença num país/região é indicada no mapa) (Anónimo, 2009b).
Actualmente, T. absoluta encontra-se na Lista A1 da “European and Mediterranean
Plant Protection Organization” (EPPO), sendo esta espécie considerada um inimigo que
pode causar avultados prejuízos, devendo as plantas e frutos procedentes de regiões onde este foi detectado serem isentos deste insecto (EPPO, 2005).
2.1.4. Hospedeiros
O tomate é o principal hospedeiro de T. absoluta, podendo atacar outras solanáceas, nomeadamente beringela, tabaco e batata, bem como espécies de solanáceas adventícias como Solanum nigrum, Solanum elaeagnifolium, Solanum puberulum,
Datura stramonium, Nicotiana glauca e Datura ferox (Camacho, et al., 2009; Payer &
Figueiredo, não public. - Anexo 10). A espécie Chenopodium murale também pode servir de alimento para as larvas (Camacho, et al., 2009).
Em Itália, foi recentemente identificado o feijão-verde (família Fabaceae) como hospedeiro (EPPO, 2005; Monserrat, 2009; Anónimo, 2009b).
2.1.5. Descrição, bioecologia e comportamento
A traça-do-tomateiro é um pequeno lepidóptero que apresenta um alto potencial reprodutivo, desde que tenha alimento disponível (Anónimo, 2009b). O seu ciclo biológico varia entre 29 a 38 dias, dependendo das condições ambientais (Biurrun, 2008; Vieira, 2008). Estudos realizados no Chile têm mostrado que o desenvolvimento leva 76,3 dias a 14ºC, 39,8 dias a 19,7ºC e 23,8 dias a 27,1º C (EPPO, 2005). De acordo com Monserrat (2009), em condições óptimas, um ovo pode alcançar o estado de adulto em pouco mais de 20 dias. No Inverno, este período pode prolongar-se até 80
6 a 90 dias (Monserrat, 2009; Serra et al., 2009). Devido à curta duração do ciclo de vida e à sobreposição de gerações, na Europa, pode chegar a apresentar 10 a 12 gerações por ano, se as condições forem favoráveis, embora apenas cinco gerações por ano tenham sido observadas na Argentina (Monserrat, 2009). De acordo com os estudos realizados na região espanhola de Múrcia, com clima muito suave, estima-se que T.
absoluta poderia completar 9-10 ciclos num um ano (Monserrat, 2009). É de realçar que
devido à recente introdução na Europa, a biologia de T. absoluta neste continente não está tão bem estudada como no continente de origem (Korycinska & Moran, 2009). A capacidade de voo dos adultos é desconhecida (Korycinska & Moran, 2009).
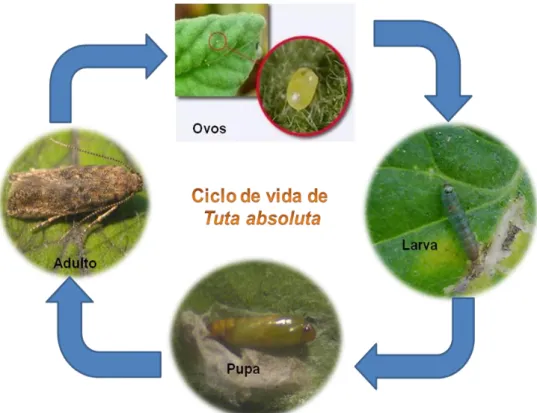
O ciclo de vida da traça-do-tomateiro (Figura 2) inclui os estados de ovo, larva, pupa e adulto (Serra et al., 2009). Cada fêmea fecundada chega a pôr entre 180 a 260 ovos ao longo de vários dias, normalmente isolados ou em pequenos grupos de não mais de 2-3 (Monserrat, 2009), de preferência nas folhas, na página inferior, mas também no caule, pedúnculo e/ou fruto (EPPO, 2005; Serra et al., 2009).
A lagarta, ao eclodir, penetra nos tecidos vegetais e alimenta-se no interior dos folíolos do tomateiro, formando uma mancha branca larga de mesófilo consumido, de contorno irregular (Serra et al., 2009), deixando a epiderme intacta (Caffarini et al., 1999). Pode alimentar-se do caule e frutos, penetrando nestes, geralmente, próximo do pedúnculo (Serra et al., 2009). As larvas podem passar de folhas ou frutos afectados para outros que estão isentos de estragos (Camacho et al., 2009). Depois de passarem por quatro instares larvares (EPPO, 2005; Biurrun, 2008), por vezes cinco (Monserrat, 2009), pupam no solo ou na superfície foliar ou ainda dentro das galerias, dependendo das condições ambientais (EPPO, 2005; Serra et al., 2009). Quando não pupa no solo, normalmente é construído um casulo (EPPO, 2005) de seda (Anónimo, 2009c). No final do desenvolvimento larvar, a lagarta transforma-se em pupa, da qual emergirá uma pequena borboleta, que tem hábitos de voo crepusculares/nocturnos, permanecendo escondida entre a folhagem durante o dia (EPPO, 2005; Anónimo, 2009; Anónimo, 2009c; Serra et al., 2009). Ao agitar a folhagem é possível observar o voo destes pequenos insectos (Serra et al., 2009). Esta praga pode passar o Inverno nos estados de ovo, pupa ou adulto (EPPO 2005; Korycinska & Moran, 2009).
Ovo
O ovo apresenta uma forma elíptica, de cor creme ou amarelada (Anónimo, 2009c), com 0,36 mm de comprimento e 0,22 mm de largura (EPPO, 2005). Os ovos são postos preferencialmente na página inferior das folhas, no pedúnculo floral ou nos frutos (Vieira, 2008). A eclosão ocorre, de acordo com EPPO (2005), 4-5 dias após a postura. No entanto, alguns autores consideram que o desenvolvimento embrionário pode
7 prolongar até 10 dias após a postura, dependendo da temperatura ambiente (Vieira, 2008).
Figura 2. Ciclo de vida de Tuta absoluta (originais da autora excepto ovo de Anónimo (2009a)).
Larva
A larva é inicialmente de cor creme com a cabeça escura, tornando-se esverdeada ou rosada do 2º ao 4º instar larvar (Anónimo, 2009c). A larva L1 mede, no início, 0,9 mm e a
L4 atinge 7,5 mm. O estádio larvar é de 12 a 15 dias, dependendo da temperatura
(EPPO 2005; Vieira, 2008). Alguns autores referem, por exemplo, que o desenvolvimento da larva pode demorar entre 25 e 35 dias (Biurrun, 2008). A duração do estado larvar varia uma vez que o desenvolvimento da larva depende tanto da temperatura como da quantidade de substrato alimentar disponível. As larvas apresentam uma armadura bucal trituradora responsável pelos estragos. Não entram em diapausa enquanto o alimento estiver disponível (EPPO, 2005; Anónimo, 2009b; Korycinska & Moran, 2009).
Pupa
A pupa é de cor acastanhada, de forma cilíndrica e mede cerca de 4 mm de largura e 10 mm de comprimento. Estas podem encontrar-se num casulo rudimentar, branco sedoso na folha ou no solo (Serra et al., 2009). A duração deste estado é de 9-11 dias (EPPO 2005; Vieira, 2008).
8
Adulto
Os adultos são pequenas borboletas (Figura 3), com cerca de 7 mm a 10 mm de comprimento (Biurrun, 2008; Anónimo, 2009c). Morfologicamente caracterizam-se por apresentarem a cabeça e o tórax cinzentos (Vieira, 2008), palpos labiais longos e recurvados para cima (Garcia-Marí, 2010), asas revestidas por escamas com pequenas manchas cinzentas e pretas (Camacho et al., 2009). As antenas são filiformes (Vieira, 2008) longas e recurvadas para trás, com tegumento alternadamente claro e escuro.
Figura 3. Adultos de Tuta absoluta realizando a cópula (original da autora).
Korycinska & Moran (2009) e Barrientos et al. (1998) demonstraram que o zero de desenvolvimento de ovo a adulto de T. absoluta é, em média, de 8,14°C, sendo os zeros de desenvolvimento para cada um dos estados indicados no Quadro 2.
Quadro 2. Zero de desenvolvimento estimado para cada um dos estados de desenvolvimento de Tuta absoluta (Barrientos et al., 1998).
Estado de desenvolvimento Zero de desenvolvimento estimado (ºC)
Ovo 6,9
Larva 7,6
Pupa 9,2
2.1.6. Sintomas, estragos e prejuízos
Tuta absoluta (Meyrick) é uma das pragas que mais ataca o tomate e é considerada
uma praga limitante para a produção de tomate na maioria dos países da América Latina, originando prejuízos até 70% (Oliveira et al., 2009). Alguns autores citam mesmo 100% de prejuízos (Anónimo, 2009b; Desneux et al., 2010). Tais estragos são provocados pela actividade alimentar das larvas.
Esta é uma praga muito prolífera produzindo estragos sobretudo na parte aérea da planta. As larvas da traça-do-tomateiro podem causar estragos em todas os estados de desenvolvimento da planta, produzindo minas nas folhas, perfurando caules, gomo
9 apical e frutos verdes e maduros (Lietti et al., 2005). Também se podem alimentar de tubérculos e sementes (Garcia-Marí & Vercher, 2010). Podem atacar qualquer fase do ciclo vegetativo da cultura (Cunha et al., 2008; Vieira, 2008) e causar prejuízos importantes na produção de diferentes regiões e em diversificados sistemas de produção (Lietti et al., 2005). No entanto, verifica-se que os principais prejuízos derivam dos ataques aos frutos, em especial devido à dificuldade da sua detecção (Serra et al., 2009).
Nas folhas, as larvas de T. absoluta fazem minas que lembram as larvas mineiras (Figura 4), nomeadamente, Liriomyza huidobrensis (Blanchard) e Liriomyza trifolii (Burgess), alimentando-se de todo o mesófilo da folha deixando apenas a epiderme intacta (Caffarini et al., 1999; Biurrun, 2008). A galeria produzida pelas larvas de
T.absoluta vão se ampliando à medida que esta se alimenta (Arnó & Gabarra, 2010).
Estas minas, posteriormente, acabam por necrosar (Vieira, 2008; Silva, 2009). Pode haver várias minas na mesma folha (Korycinska & Moran, 2009) com restos de excrementos escuros acumulados numa zona (Camacho et al., 2009). As minas apresentam áreas irregulares mais ou menos amplas; colocando a folha em contra-luz, tal como se faz para as larvas mineiras, pode-se observar a lagarta que em geral são mais escuras do que aquelas (Biurrun, 2008). Com frequência, as lagartas saem para o exterior da folha para iniciar novas galerias ou se deslocar para outras folhas ou frutos e nestes podem penetrar em toda a superfície mas, mais frequentemente, próximo da zona peduncular (Serra et al., 2009) e em qualquer estado de maturação (Monserrat, 2009).
Fernandez & Montagne (1990) e Garcia-Marí et al. (2010) alertam para a dificuldade em distinguir no campo Tuta absoluta das espécies Keiferia lycopersicella Walsingham e
Phthorimaea operculella Zeller
(ambas também Gelechiidae), uma vez que os estragos
provocados por ambas são parecidos aos da traça-do-tomateiro. A espécie Keiferialycopersicella está referida na Europa, em Itália, onde foi introduzida acidentalmente,
em 2008 (Savino & Espinosa, 2008).
Nos frutos, as larvas formam galerias que rapidamente são susceptíveis a infecções secundárias de patogénios (Vieira, 2008; Arnó & Gabarra, 2010), conduzindo a podridão dos frutos (EPPO, 2005) e, consequentemente, inviabilizando a sua comercialização (Cunha et al. 2008), chegando a afectar até 81% dos frutos (Caffarini et al. 1999). Os orifícios de entrada são pequenos (Serra et al., 2009).
10
Figura 4. Minas de Liriomyza sp. (galerias, setas azuis) e de Tuta absoluta (setas a vermelho) em folhas de tomate. Observam-se também estragos no caule (Autoria: Elisabete Figueiredo, 2010).
As galerias quando efectuadas no caule afectam o crescimento das plantas (Caffarini et
al. 1999). No entanto, verifica-se que os estragos em gomos apicais e nos caules são
menos frequentes que a presença de minas nas folhas (Biurrun, 2008).
Na batata cultivada em zonas quentes e de baixas atitudes (abaixo dos 1000 m), T.
absoluta é considerada uma das pragas mais importantes das folhas (EPPO, 2005;
Anónimo, 2009b). Ataca a parte aérea desta solanácea (Figura 6), não se desenvolvendo nos tubérculos (EPPO, 2005; Vieira, 2008) Contudo, recentemente, foram referidos estragos nos tubérculos (Anónimo, 2009b).
Nas Figuras 5 a 12, podem se observar os estragos directos causados pelas larvas da traça-do-tomateiro em diferentes hospedeiros.
Figura 5. Folhas de Solanum nigrum com sintomas de Tuta absoluta no campo de tomate (originais da autora).
11
Figura 6. Larva e estragos de Tuta absoluta em folhas de batateira, no distrito de Setúbal (originais de Eng. Paula Santos, AT& F.)
Figura 7. Estragos causados por Tuta absoluta em plântula de pimento (Capsicum annuum) à esquerda e em beringela (Solanum melongena) à direita (originais de EPPO Gallery 2010).
Figura 8. Observação de larvas, galerias em folhas e adulto de Tuta absoluta em tomate de estufa na região Oeste (originais da autora).
12
Figura 10. Larva de Tuta absoluta e estragos em frutos verdes e maduros no campo de tomate. Observam-se os orifícios de entrada e saída, principalmente junto ao pedúnculo (originais da
autora).
Figura 11. Fruto verde com duas pupas de Tuta absoluta no pedúnculo. Em ambos os frutos são visíveis os estragos (originais da autora).
Figura 12. Estragos em frutos provocados pela alimentação larvar da traça-do-tomateiro (originais da autora).
2.1.7. Estimativa do risco e meios de protecção
A base de uma protecção eficaz contra Tuta absoluta é o conhecimento da sua biologia e etologia, sendo necessário integrar, de uma forma sustentável, todos os meios de protecção disponíveis, uma vez que os estragos podem assumir uma importância assinalável, sobretudo se a detecção não for precoce (Serra et al., 2009).
Alguns aspectos são importantes na hora de estabelecer as estratégias de protecção. Um destes aspectos é o facto de as larvas que penetram directamente nos frutos, de postura realizada sobre o pedúnculo, não saírem para o exterior durante as primeiros
13 instares larvares, dificultando o seu combate. Também assume importância o facto de, na ausência de plantas hospedeiras, as fêmeas fecundadas poderem sobreviver na parcela durante várias semanas, ao contrário dos machos, uma vez que estes apresentam uma menor viabilidade. O solo também pode estar contaminado com a praga no estado de pupa (Monserrat, 2009), durante muito tempo, sobretudo durante o Inverno.
Monitorização e estimativa do risco
O uso de armadilhas com feromona é um método confiável para detectar a presença de
Tuta absoluta, bem como para avaliar a densidade das populações (Anónimo, 2009b).
Outro método importante é a observação visual de folhas, flores, frutos e caules atacados (Serra et al., 2009).
A monitorização pode ser realizada através da instalação de armadilhas sexuais do tipo delta, brancas, com feromona específica para Tuta absoluta, de forma a capturar os machos e detectar precocemente a praga (Serra et al., 2009). Em culturas protegidas, deve-se colocar 2-4 armadilhas/ha, próximo de aberturas, a 1,5-2 m de altura, para detectar a presença da praga e a 40 cm de altura para captura em massa no caso de esta praga já estar instalada. Deve-se efectuar contagens semanais, retirar os insectos capturados e trocar a feromona a cada 6-7 semanas (Anónimo, 2009c), de acordo com as instruções do fabricante.
Protecção cultural
Foi demonstrado por Monserrat (2009) que as parcelas em que se verificou ataques de
T. absoluta podem ficar contaminadas durante várias semanas através da presença de
pupas no solo, como já foi referido atrás. A presença desta praga sobre o terreno pode variar entre 4 e 6 semanas, podendo este período prolongar-se caso as temperaturas sejam baixas. Assim, com base nesta informação, uma das primeiras medidas de protecção para a redução da incidência da praga em novas plantações é deixar um período de 4-6 semanas desde a limpeza dos restos da cultura até à nova transplantação para permitir a eliminação de adultos antes da colocação da nova cultura (Monserrat, 2009; Serra et al., 2009).
Outras medidas culturais a adoptar são:
- fazer rotação com não solanáceas (Serra et al., 2009);
- eliminar e destruir os materiais infestados, colocando-os em sacos bem fechados (Anónimo, 2009c);
14 - eliminar infestantes solanáceas na proximidade das áreas infestadas para evitar a acumulação de um reservatório populacional potencial (Korycinska & Moran, 2009); - instalar e fazer a manutenção de redes de exclusão nas aberturas das estufas, de modo a impedir a entrada dos insectos adultos, sempre que tal não impossibilitar a protecção contra doenças criptogâmicas por diminuição de ventilação (Serra et al., 2009); por exemplo, utilização de malha de 10x20 fios/cm2 (Camacho et al., 2009); - fazer rega por aspersão (Vieira 2008); no entanto, ter em atenção doenças como o míldio;
- utilizar plântulas isentas da praga (Serra et al., 2009);
- destruir pupas dos restos de culturas e do solo, quando existem densidades elevadas (Anónimo, 2009c);
- fazer solarização para a eliminação de pupas que se refugiam no solo (Camacho et al., 2009);
- utilizar genótipos resistentes a Tuta absoluta (Giustolin et al., 2001).
Em relação a este último aspecto, alguns estudos estão sendo direccionados para a procura de variedades de tomate resistentes a Tuta absoluta (EPPO, 2005; Oliveira et
al., 2009).
Protecção biotécnica
A captura em massa é uma técnica que envolve a colocação de um maior número de armadilhas na área de cultura em várias posições estratégicas para capturar os machos antes de fecundarem as fêmeas. É amplamente utilizada em conjugação com outras medidas de protecção para reduzir até níveis aceitáveis os estragos e simultaneamente reduzir os tratamentos com insecticidas (Anónimo, 2009b).
A captura em massa da traça-do-tomateiro pode ser realizada através da colocação de armadilhas de água com feromona sexual (Figura 13) a 40 cm de altura do solo (Serra
et al., 2009), utilizando algumas gotas de detergente ou óleo para baixar a tensão
superficial da água e, assim, evitar que os adultos escapem da armadilha (Camacho et al., 2009). Para além destas medidas, a água deve ser renovada frequentemente (Serra
et al., 2009). O recipiente deverá ter uma grande superfície, com aberturas amplas e
com a feromona próxima da água. É aconselhável a colocação nas estufas de 15-20 armadilhas/ha e ao ar livre de 20-40 armadilhas/ha (Anónimo, 2009c). No entanto, por razões práticas, verifica-se que a captura em massa tem maior hipótese de sucesso em culturas protegidas (Anónimo, 2009b).
15 Camacho et al. (2009) aconselham a utilização de armadilhas amarelas adesivas (50/ha) e com feromona (20-25/ha) para capturar os adultos. Estas últimas devem ser, segundo estes autores, distribuídas regularmente, junto ao solo, colocando-as duas semanas antes da plantação, uma vez que a longevidade das fêmeas pode atingir 15 dias.
Figura 13. Armadilha de água comercial (à esquerda) e artesanal (à direita) com feromona sexual para a captura da traça-do-tomateiro (originais da autora).
Outra técnica que pode ser utilizada é a colocação de armadilhas luminosas. Tem a vantagem de capturar tanto machos como fêmeas. Deve-se colocar as armadilhas apenas em estufas bem fechadas para evitar atrair mais adultos do exterior e colocar um recipiente com óleo debaixo da armadilha para recolher as traças que caiem depois de chocarem contra esta. Também se podem complementar as armadilhas luminosas com placas cromotrópicas adesivas para capturar as traças que voam à volta, mas não chocam, com as armadilhas luminosas (Camacho et al.,2009).
Protecção química
Características biológicas da espécie, como a sobreposição de gerações e o hábito das larvas se alimentarem em galerias, diminuem a eficácia da protecção química (Caffarini
et al., 1999). No entanto, o principal método de protecção contra a traça-do-tomateiro
tem sido a utilização de insecticidas (Oliveira et al., 2009; Desneux et al., 2010). Apesar de se efectuarem tratamentos em número elevado, em alguns casos, até 36 por cultura (Cunha et al., 2008; Anónimo, 2009b), a protecção química não se tem revelado satisfatória (Medeiros et al., 2009) devido ao desenvolvimento de populações resistentes às substâncias activas empregadas e à eliminação de populações dos inimigos naturais da traça-do-tomateiro (Cunha et al., 2008). Há relatos de resistência de T. absoluta a abamectina, cartap, metamidofos e permetrina (Anónimo, 2009b).
No Brasil, uma das formas encontradas para diminuir o desenvolvimento de resistência por este insecto foi a alternância de grupos químicos, como por exemplo, organofosforados, piretróides, carbamatos, reguladores de crescimento, etc. Cada um
16 deles deve ser utilizado por um período de 28 dias para atingir aproximadamente uma geração da praga. Os insecticidas piretróides e organofosforados devem ser utilizados, preferencialmente, no período da manhã, quando ocorre a menor actividade dos adultos com o objectivo da selecção de populações resistentes ocorrer apenas num estado de desenvolvimento, isto é, no estado larvar (Bôas et al., 2005). Entre as substâncias activas recomendadas no Brasil para combate a esta praga estão flufenoxurão, lufenurão, teflubenzurão e tebufenozida (RCI- reguladores de crescimento de insectos), abamectina (avermectina), clorpirifos e fosalona (organofosforados), carbaril (carbamato) e delta-endotoxinas de Bacillus thuringiensis Berl. (Vieira, 2008).
Em Espanha, têm sido utilizadas substâncias activas que têm demonstrado eficácia satisfatória contra a larva de T. absoluta: delta-endotoxinas de Bacillus thuringiensis var.
kurstaki, indoxacarbe e spinosade e a deltametrina revelou ter um efeito "knock-down"
napopulação adulta (Anónimo, 2009b; Korycinska & Moran, 2009).
Em Portugal, apenas o indoxacarbe está homologado pela DGADR (Direcção-Geral de Agricultura e Desenvolvimento Rural) (DGADR, 2010). No entanto, o COTHN (Centro Operativo e Tecnológico Hortofrutícola Nacional) divulgou que a AIHO (Associação Interprofissional de Horticultura do Oeste) obteve uma autorização excepcional na utilização de benzoato de emamectina para o combate a Tuta absoluta na cultura do tomateiro, de 14 de Abril a 14 de Agosto de 2010 (DGADR, 2010).
Protecção biológica
Na protecção biológica contra T. absoluta há que optimizar e integrar estratégias de protecção que fomentem a biodiversidade dos inimigos naturais deste fitófago. Para tal, é importante o conhecimento dos inimigos naturais da zona de que é originário ou em que já está há muitos anos estabelecido (Quadro 3).
No Brasil, Bacci et al., (2008) verificaram uma maior mortalidade no estado de ovo de T.
absoluta devido a predação por percevejos e ao parasitismo por Trichogramma pretiosum (Riley). Entre os percevejos destacam-se: Orius sp., Lasiochilus sp. e Xylocoris sp. (Hemiptera: Anthocoridae), Annona bimaculata (Distant) e Hyaliodocoris insignis (Stal) (Hemiptera: Miridae). Estes são excelentes predadores de ovos e larva L1
(Bacci et al., 2008). Os mesmos autores apontaram a vespa predadora Protonectarina
sylveirae (Saussure) (Hymenoptera: Vespidae) como potencial agente de protecção
biológica, uma vez que estas apresentaram uma maior actividade de predação. A mortalidade da traça-do-tomateiro por P. sylveirae ocorreu nos instares L2, L3 e L4, com
17
Quadro 3. Inimigos naturais mais importantes de Tuta absoluta na América Latina.
Tipo de inimigo natural
Ordem Família Espécie Referência
P
re
dador
Hemiptera Anthocoridæ
Xylocoris sp. Bacci et al., 2008; Miranda et al., 1998 Orius sp.
Bacci et al., 2008
Lasiochilus sp.
Miridæ Annona bimaculata (Distant)
Hyaliodocoris insignis (Stal)
Pentatomidæ Podisus nigrispinus (Dallas) Vivan et al., 2002
Hymenoptera Vespidæ Protonectarina sylveirae (Saussure) Bacci et al., 2008;
Coleoptera Coccinellidæ Cycloneda sanguinea (L.)
Miranda et al., 1998
Thysanoptera Phlaeothripidæ sp. Não identificada
Neuroptera Chrysopidæ Chrysoperla externa (Hagen) Alvino et al., 2009
P a ra s it ói de Hymenoptera
Bethilidæ Goniozus nigrifemur (Ashmead) Miranda et al., 2005;
Miranda et al., 1998
Ichneumonidæ Campoplex haywardi (Blanchard)
Polack, 2007
Diadegma sp. Temelucha sp.
Braconidæ Apanteles gelechiidivoris (Marsh)
Bajonero et al., 2008
Pseudapanteles dignus (Muesebeck) Polack & Brambilla, 2000
Earinus sp. Marchiori et al., 2004; Bacci et al., 2008
Bracon sp.
Miranda et al., 2005; Marchiori et al., 2004; Bacci
et al., 2008 Bracon lucileae (Marsh) Berta & Colomo, 2000
Bracon tutus (Berta e Colomo) Berta & Colomo, 2000
Bracon lulensis (Berta e Colomo) Berta & Colomo, 2000
Chelonus sp. Hernández et al., 2008;
Miranda et al., 2005
Orgilus sp Hernández et al., 2008
Eulophidæ Dineulophus phthorimaeae (De Santis) Larraín 1986
Neochrysocharis formosa (Westwood) Polack 2007
Trichogrammatidæ
Trichogramma pretiosum (Riley) Bacci et al., 2008; Miranda et al., 2005 Trichogramma exiguum Pinto &
Platner Noyes, 2003
Trichogramma nerudai Pintureau &
Gerding Noyes, 2003
Trichogrammatoidea bactrae Nagaraja Noyes, 2003
Chalcididæ Conura sp. Marchiori 2004
Diptera Tachinidæ Exoristini sp. Colomo & Berta, 2006
P a ra s it a
Acari Pyemotidæ Pyemotes sp. Oliveira et al., 2007
Na Argentina, Colomo & Berta (2006) referem que os parasitóides pertencentes à ordem Hymenoptera são os principais inimigos naturais de T. absoluta. Estes autores descreveram pela primeira vez um díptero a atacar a traça-do-tomateiro. Trata-se de uma espécie de endoparasitóide solitário pertencente à família Tachinidae, subfamilia Goniinae, tribo Exoristini.
18 Foram ainda referidos os parasitóides larvares Campoplex haywardi (Blanchard) e Neochrysocharis formosa (Westwood), e outros parasitóides dos géneros Diadegma,
Orgilus e Temelucha que podem também parasitar a traça-do-tomateiro (Polack, 2007).
A mortalidade causada por predadores pode chegar até 79,8%, devido maioritariamente à acção de predadores generalistas (Hernández et al., 2008). Segundo Vivan et al. (2002) a espécie neotropical Podisus nigrispinus (Dallas) apresenta um elevado potencial de predação de larvas da traça-do-tomateiro.
Marchiori et al. (2003) descrevem pela primeira vez 13 espécies de Conura sp. (Chalcididae) parasitando pupas de Tuta absoluta, em Minas Gerais (Brasil) com taxa de parasitismo de 2,6%. Posteriormente, em 2004, estes autores verificaram uma taxa de parasitismo de 4,2 % e 0,2% para um complexo de braconídeos composto por 21 espécies de Bracon sp. e uma espécie de braconídeo Earinus sp., respectivamente (Marchiori et al., 2004). Estas espécies têm preferência por larvas L3 e L4 (Bacci et al.,
2008). Também no Brasil, Miranda et al. (2005) referiram parasitismo larvar de T.
absoluta por Bracon sp., Chelonus sp. Goniozus nigrifemur (Ashmead) (Bethylidae), e
parasitóides oófagos principalmente Trichogrammatidae, tendo a capacidade de se estabelecer mesmo com densidades muito baixas de ovos de T. absoluta.
No que se refere a observação na região Paleártica, em Espanha, com o percevejo
Nabis pseudoferus (Remane), foram realizados dois ensaios biológicos de semi-campo,
com a libertação de 8 ou 12 ninfas do primeiro instar de N. pseudoferus por planta, demonstrando uma redução importante no número de ovos de T. absoluta, entre 92 e 96 %, respectivamente (Cabello et al., 2009a). Também em Espanha, Mollá et al. (2010) observaram os ácaros fitoseídeos Amblyseius swirskii Athias-Henriot e Amblyseius
cucumeris Oudemans predando ovos de T. absoluta, podendo também se alimentar de
larvas do primeiro instar. Recentemente, foram assinaladas espécies de parasitóides autóctones que adaptaram a traça-do-tomateiro como novo hospedeiro (Arnó & Gabarra, 2010; Desneux et al., 2010)
O parasitóide idiobionte Necremnus artynes (Walker) (Hymenoptera: Eulophidae) foi observado na região de Valência parasitando, principalmente, larvas de segundo e terceiro instar de T. absoluta (Mollá et al., 2010).
Trichogramma achaeae Nagaraja & Nagarkatti é um parasitóide oófago introduzido em
Espanha e que demonstrou ter uma eficácia elevada. Foi testado em largadas de 30 adultos/planta a cada 3-4 dias, nos meses de Agosto e Setembro, proporcionando reduções nos estragos de 91,74% (Cabello et al., 2009b). Neste mesmo país, aconselha-se a introdução de mirídeos desde as primeiras semanas da cultura, através