a
U
NIVERSIDADE DE
T
RÁS
-
OS
-M
ONTES E
A
LTO
D
OURO
D
EPARTAMENTO DE
G
ENÉTICA E
B
IOTECNOLOGIA
Development of models to determine dynamic GNE expression
in vivo to aid resolution of sialic acid dysfunction associated with
Hereditary Inclusion Body Myopathy
Sofia Melo Pereira
Dissertação orientada por:
Doutor Professor Moisés Mallo Perez
Doutora Jennifer Rowland
M
ESTRADO EM
B
IOTECNOLOGIA PARA AS
C
IÊNCIAS DA
S
AÚDE
I
A
CKNOWLEDGMENTSApós a realização desta dissertação de mestrado, resta-me agradecer a todas as pessoas e entidades que contribuíram para a sua realização:
À Coordenação de Curso e Comissão de Estágios, pela aceitação do projecto de estágio. Ao Instituto Gulbenkian de Ciência, a instituição científica em que este trabalho foi realizado, por todas as facilidades concedidas para a realização deste estágio.
Ao Moisés Mallo, por ter aceitado ser meu orientador de estágio pela possibilidade de integração no seu grupo de investigação, assim como pelos ensinamentos prestados com profissionalismo e competência que sempre demonstrou. Agradeço também a tua disponibilidade, interesse e correcção cuidada desta dissertação de mestrado.
À Jenniffer Rowland, por ter aceitado ser minha co-orientadora de estágio. Jen, in the first place, thank you for “stealing” me; otherwise, none of this would have ever happened. Thank you for looking at a lost girl and seeing a great future. Thank you for always believing in me, even in the most difficult times, you have become a great friend that I hope never to lose. And, at last, thank you Rafa for coming into this world.
À Sílvia Cardoso e Sofia Rebelo, por toda amabilidade e carinho sempre disponíveis.
A todas as meninas e menino do laboratório, pelo espírito de equipa e bons momentos passados ao longo deste ano. Decididamente, com vocês, o trabalho tornou-se bem mais divertido. Ficarão para sempre no meu coração.
Ao Miguel, um amigo incondicional, obrigada pela infinita paciência e carinho.
Mais uma vez, aos meus pais, pois tudo o que sou hoje devo-o a vocês. Obrigada por “stressarem” mais do que eu.
II
A
BSTRACTSialic acids mainly occur as terminal components of cell surface glycoproteins and glycolipids, playing a major role in the biological diversity of glycoconjugates and in the variety of cellular signaling functions (Varki, 1997). In sialic acid biosynthesis, the bifunctional rate-limiting enzyme UDP-N-acetylglucosamine-2-epimerase/N-acetylmannosamine kinase (GNE) catalyzes the two initial steps (Keppler et al., 1999) and, therefore, is responsible for the regulation of cell surface sialylation. It is also known to be essential during early embryonic development, since the complete inactivation of the GNE gene by gene targeting leads to early embryonic lethality in mice (Schwarzkopf, et al., 2002). This statement is further supported by the absence of homozygous null mutations in humans (Huizing and Krasnewich, 2009). However, mutations in the GNE enzyme are known to cause two human disorders, sialuria and hereditary inclusion body myopathy (HIBM).
Sialuria is an autosomal dominant disorder and one missense mutation in the allosteric site of GNE is sufficient to cause loss of feedback inhibition of epimerase GNE activity, resulting in cytoplasmic accumulation and excretion of large quantities of free sialic acid in the urine (Seppala et
al., 1999). On the other hand, HIBM is an autosomal recessive disorder and characterized by
adult-onset of muscle weakness in upper and lower limbs, with an interesting sparing of the quadriceps muscles. Patients with two GNE mutations throughout the GNE enzyme (except in the allosteric site), usually have decreased GNE enzyme activity which, in some cases, can lead to decreased sialylation of glycoproteins and to the appearance of HIBM phenotype (Noguchi et al., 2004, Huizing et al., 2004). The mechanisms and cellular pathways affected by GNE mutations and causing the muscle weakness were not identified so far.
A number of studies have been pursued over the past decade to investigate the role of GNE both in vitro and in vivo. Herein, these studies are reviewed to analyze where the field has been leading and its current body of knowledge which aims to shed light in particular on the mechanism of the HIBM, the essential role that GNE plays in the human body and the type of therapies that may be able to be utilized in the future to treat patients suffering from sialic acid related disorders.
The aim of this thesis was to conduct an investigation to evaluate the exact tissue specific expression of GNE in vivo, during embryogenesis and adulthood. For this, two main approaches were pursued in tandem: one was to develop a transgenic mouse line, where β-galactosidase expression would be under control of the native GNE promoter and thus permitting the tracking of GNE expression in vivo by β-galactosidase staining; and the other one was to search for key points during development where sialylation might be most important. For this last approach, whole-mount in situ hybridization was first conducted on wild-type mouse embryos, in which the results would be later confirmed by β-galactosidase staining of transgenic mouse embryos. Unfortunately, these transgenic mice were not able to be produced by the time of this thesis conclusion and so, other techniques were followed for a better understanding of the role of GNE during embryogenesis, like Northern
III blot hybridization and real-time PCR analysis. Relying only in the mouse development stages investigated in this thesis, from E7.5 until E11.5, we found some evidence to support the hypothesis that sialic acid might be crucial at certain time points during embryogenesis, such as at age E8.0, where the placental tissues must act like a vital source of sialic acid, reassuring the needs of the developing embryo. At age E11.5, we also found that requirements of sialic acid are very high and that GNE transcripts are mainly localized in middle portion of the embryo’s body, in particular in the right atria of the developing heart.
IV
R
ESUMOOs ácidos siálicos ocorrem maioritariamente como componentes terminais da superfície celular de glicoproteínas e glicolípidos, desempenhando um papel essencial na diversidade biológica dos glicoconjugados e na variedade de funções de sinalização celular (Varki, 1997). Na biossíntese do ácido siálico, a enzima bifuncional UDP-N-acetilglucosamina-2-epimerase/N-acetil Manosamina cinase (GNE) actua de uma forma limitada e catalisa as duas etapas inicias (Keppler et al., 1999), sendo por isso responsável pela regulação pelo processo de sialilação da superfície celular. A enzima é também reconhecida por essencial durante o desenvolvimento embrionário, pois a completa inactivação deste gene em murganho resulta numa precoce letalidade embrionária (Schwarzkopf, et
al., 2002). Esta afirmação é confirmada pela ausência de indivíduos homozigóticos com mutações
nulas em ambos os alelos do gene GNE (Huizing e Krasnewich, 2009). Contudo, mutações neste gene são a causa de duas doenças na população humana, designadamente sialuria e “hereditary inclusion body myopathy” (HIBM).
A sialuria é uma doença dominante autossómica, em que uma única mutação missense no local alostérico do gene GNE é suficiente para causar a perda de inibição por “feedback” da actividade enzimática da epimerase do GNE, resultando na acumulação citoplasmática de ácido siálico livre e excreção de grandes quantidades deste componente na urina (Seppala et al., 1999). Por outro lado, HIBM é doença de carácter autossómico recessivo, iniciando-se habitualmente na idade adulta, e caracterizada por uma fraqueza muscular dos membros superiores e inferiores, com expecção dos músculos quadríceps. Indivíduos com duas mutações no gene GNE (excluindo o sítio alostérico) por norma têm uma actividade enzimática do GNE diminuída a qual, por vezes, pode levar a um decréscimo de sialilação de glicoproteínas e ao aparecimento do fenótipo típico de HIBM (Noguchi et al., 2004, Huizing et al., 2004). Até hoje não foram identificados mecanismos ou vias celulares afectados por mutações neste gene.
Na última década, um grande número de estudos têm sido efectuados de forma a investigar a função essencial do GNE, tanto in vivo como in vitro. Na presente tese, diversos estudos são analisados de forma abordar as áreas de investigação deste tema, as quais têm por finalidade a pesquisa de possíveis mecanismos que levam a HIBM, o papel essencial que o GNE desempenha no corpo humano e o tipo de terapias que poderão ser utilizadas futuramente no tratamento de pessoas com doenças relacionadas a deficiências de ácido siálico.
O principal objectivo desta tese foi efectuar um trabalho de investigação para avaliar a expressão específica do GNE in vivo, durante a idade embrionária e adulta. Assim, duas abordagens foram essencialmente seguidas em conjunto. A primeira consistiu no desenvolvimento numa linha transgénica de murganhos, onde a expressão do gene de β-galactosidase estaria sobre o controlo do promotor nativo do gene GNE, permitindo assim o acompanhamento da expressão do GNE in vivo, através da coloração de β-galactosidase. A segunda abordagem consistiu na investigação de pontos
V fulcrais durante o desenvolvimento embrionário, no qual o processo de sialilação poderá ser mais relevante. Para esta última abordagem, foram efectuadas técnicas como a hibridação in situ em embriões “wild-type” de murganhos, nos quais os resultados seriam mais tarde confirmados pela coloração de β-galactosidase nos embriões transgénicos de murganho. Infelizmente, a produção dos animais transgénicos não foi possível de ser realizada até a altura de conclusão desta tese e, portanto, outras técnicas foram seguidas para uma melhor compreensão da função do GNE durante a embriogénese, tais como hibridação por Northern blot e análise de PCR em tempo real. Com base nos estádios de desenvolvimento analisados neste projecto, entre E7.5 e E11.5, encontramos algumas evidências que apoiam a hipótese de que o ácido siálico poderá ser essencial em algumas alturas durante a embriogénese, tal como na idade embrionária E8.0, onde os tecidos placentários poderão actuar como uma fonte vital deste componente, assegurando as necessidades do embrião em desenvolvimento. No dia embrionário E11.5, de forma similar, averiguamos que as necessidades de ácido siálico são muito elevadas e que os transcritos de GNE estão maioritariamente localizados na porção média do embrião, mais propriamente na artéria direita do coração em desenvolvimento.
T
ABLE OF CONTENTSAcknowledgments ... I Abstract ... I Resumo ... IV List of Figures ... VIII List of Tables ... IX List of Abbreviations ... X Chapter I Introduction
Overview ... 1
I.1 Biology of Sialic Acid ... 1
I.1.1 Cellular and Molecular interactions ... 2
I.1.2 Sialic acid Distribution ... 2
I.1.3 Sialic Acid Biosynthesis ... 4
I.2 UDP-N-acetylglucosamine 2-epimerase/ N-acetylmannosamine kinase Enzyme ... 6
I.2.1 GNE gene, transcripts and proteins ... 6
I.2.2 GNE Biochemistry and Post-transcriptional modifications ... 8
I.2.3 GNE expression Analysis ... 9
I.3 Sialic acid disorders and Mutation Spectrum of GNE...11
I.3.1 Sialuria ... 11
I.3.2 Hereditary Inclusion Body Myopathy ... 13
I.4 Transgenic animal models ...17
I.4.1 Complete Knockout of GNE ... 17
I.4.2 GNE Knockin Mouse – M712T mutation ... 19
I.4.3 GNE Knockin Mouse – D176V mutation ... 19
Overall findings About GNE ...20
I.5 Objectives ...21
Chapter II Materials and Methods II.1 Production of a transgenic GNE construct ...22
II.1.1 Molecular cloning of genomic homology arms and plasmid fragments ... 22
II.1.2 Molecular cloning of targeting construct ... 28
II.1.3 Sequencing ... 29
II.1.4 Construct DNA preparation ... 30
II.2 Cell Culture ...31
II.2.1 Production and selection of homologous recombinants ... 31
II.3 Southern Blot hybridization ...33
II.3.1 Preparation of Random-Prime labeled probes... 34
II.3.2 Southern Blot Probes ... 34
II.3.3 Restriction digest on ES cell genomic DNA ... 34
II.3.4 Gel electrophoresis and Southern Blotting ... 35
II.3.5 Southern Blot hybridization ... 35
II.4 Mouse strain and housing conditions ...36
II.4.1 Mice dissection and sample collection ... 36
II.4.2 Time mating and embryo collection ... 36
II.5 Whole-mount in situ hybridization ...36
II.5.1 RNA probe preparation for whole-mount in situ hybridization ... 37
II.5.2 Whole-mount in situ hybridization of mouse embryos ... 39
II.5.3 Histological Analysis ... 40
II.6 Extraction of total RNA ...41
II.7 Northern Blot Hybridization ...41
II.7.1 Northern Blot Probes ... 42
II.7.2 Sample preparation ... 42
II.7.3 Gel electrophoresis and Northern Blotting ... 42
II.7.4 Northern Blot hybridization... 43
II.8 Synthesis of cDNA by RT-PCR ...43
II.9 Transcript Quantification by SYBR Green Real-Time PCR ...43
II.9.1 Transcript Quantification analysis ... 44
Chapter III Results Results I Creation of a ES cell line carrying a GNE allele with -galactosidase replacing GNE ...45
III.1 The gene targeting strategy ... 45
III.2 Design and Creation of a Transgenic GNE construct ... 47
III.3 Homologous recombination of ES cells ... 53
Results II Analysis of GNE expression during embryonic development ...58
III.4 Whole-mount in situ hybridization Analysis ... 58
III.5 Extraction of total RNA ... 61
III.6 Northern Blot Analysis ... 62
III.7 Transcript Quantification Analysis ... 63
Chapter IVDiscussion and Conclusion IV.1 Creation of a ES cell line carrying a GNE allele with -galactosidase replacing GNE ...68
IV.2 Analysis of GNE expression during embryonic development ...70
IV.3 Conclusion and Future Prospects ...73
References ... 74 Appendixes
Appendix I – Buffers, Solutions and Media ... A-1 Appendix II – Plasmid Maps ... A-8 Appendix III – Detailed Sequences ... A-10 Appendix IV – Collection of Biological Samples ... A-15 Appendix V – Transcript Quantification Output ... A-18
VIII
L
IST OFF
IGURESFigure 1 – Conformation of N-acetylneuraminic acid and N-glycoloylneuraminic acid ... 1
Figure 2 – Sialic acid biosynthesis ... 5
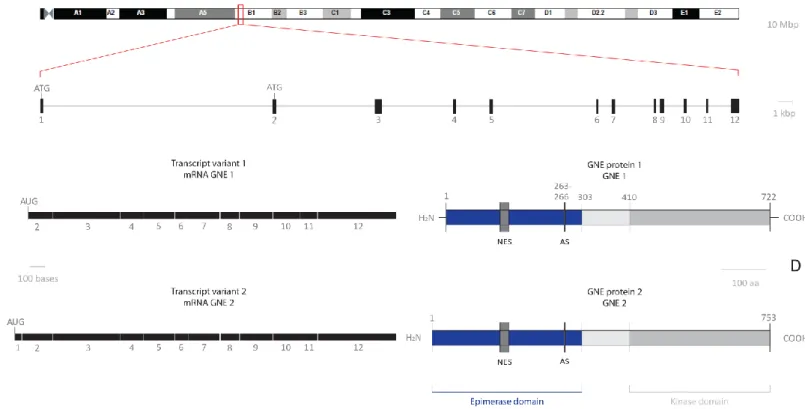
Figure 3 – GNE genomic locus, mRNA variants and protein isoforms in mouse. A – Chromosome 4 of Mus musculus. B – The mouse GNE gene structure. C – Two full length GNE mRNA transcript variants naturally occurring in mice (transcript variant 1 and transcript variant 2). D – Schematic representation of GNE protein structure ... 7
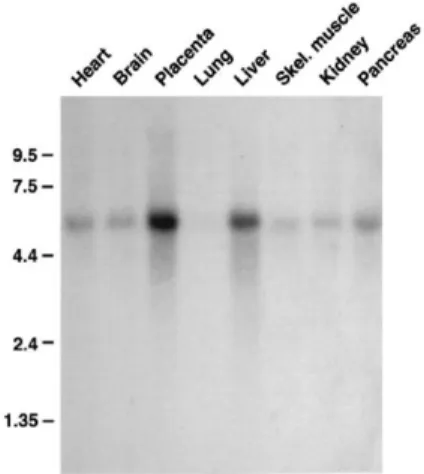
Figure 4 – Northern blot analysis of mouse RNA for expression of GNE ... 9
Figure 5 – Northern blot analysis of human RNA for expression of GNE ... 9
Figure 6 – Gene expression levels of GNE in the central nervous system, liver and small and large intestine throughout development and aging in the mouse. ...10
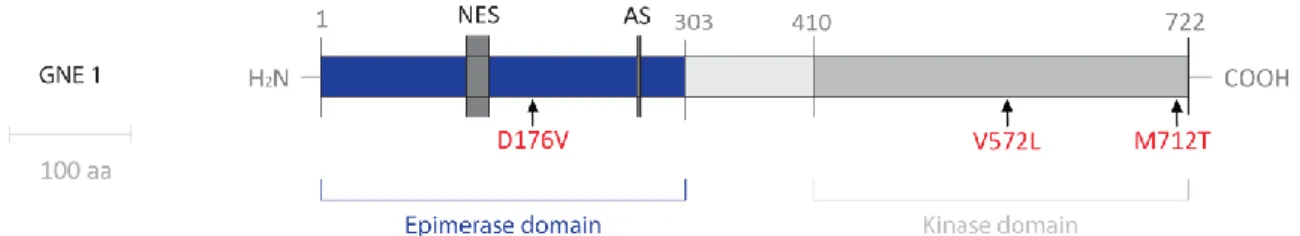
Figure 7 – Schematic drawing representing GNE enzyme with major HIBM mutations ...14
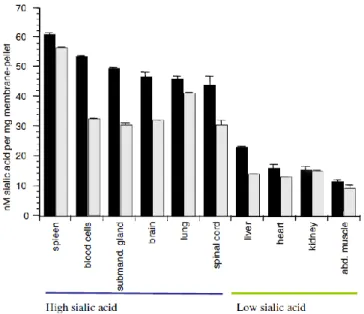
Figure 8 – Quantification of membrane-bound sialic acid in several organs ...18
Figure 9 – Genomic DNA locus of GNE. A – Chromosome 4 of Mus musculus. B – The mouse GNE gene structure. C – Genomic fragments obtained by PCR to create the GNE transgenic construct. D – Schematic representation of the genomic homology arms necessary to create the GNE construct...22
Figure 10 – Plasmid fragments obtained by PCR to create a transgenic GNE construct ...23
Figure 11 – Schematic representation of 5’ β-galactosidase reporter in frame incorporation to the start codon (ATG) of exon 2 ...24
Figure 12 – GNE gene targeting strategy. A - The mouse GNE gene structure, with focus on exon 2 and surrounding introns. B – Transgenic GNE construct, with a short (5’) and long (3’) genomic homology arms, a β-galactosidase reporter with a poly-A sequence and neomycin cassetteflanked by FRT sites ...47
Figure 13 – Cloning strategy to assemble 5’ homology arm with β-galactosidase ...48
Figure 14 – Cloning strategy to assemble 3’ homology arm with a neomycin cassette ...50
Figure 15 – Cloning strategy to assemble the final construct ...52
Figure 16 – Homologous recombination of transgenic GNE construct and overview of the Southern blot strategy ...53
Figure 17 – Genomic DNA from ES cells homologous recombinants digested with XbaI enzyme ...55
Figure 18 – Southern blot results using XbaI/Probe Ex3 of four different membranes, namely Plate I -Top gel, Plate I – Bottom gel, Plate II – Top Gel, Plate II – Bottom Gel ...56
Figure 19 – Southern blot results using XbaI/Probe β-gal for only one membrane (Backup Plate I – Top gel) ...57
Figure 20 – Representative scheme of probes used in whole-mount in situ hybridization analysis ...58
Figure 21 – Whole-mount in situ hybridization results on embryos E9.5 (A), E10.5 (B) and E11.5 (C) .60 Figure 22 – Whole RNA extracts on several tissues, after total RNA extraction with Trizol ...61
Figure 23 – Representative scheme of probes used in Northern blot hybridization analysis ...62
Figure 24 – Northern blot analysis of mouse RNA from different tissues for expression of GNE ...63
IX
Figure 26 – cDNA amplification products with GNE and GAPDH primers ...65
Figure 27 – GNE transcript quantification in four adult mouse tissues obtained from Real-Time PCR, using a SYBR Green approach ...65
Figure 28 – GNE transcript quantification in several mouse embryonic tissues obtained from Real-Time PCR, using a SYBR Green approach ...66
L
IST OFT
ABLES Table 1 – Quantification of membrane bound sialic acid in several organs of a WT mouse ... 3Table 2 – Conditions of PCR reaction and cycling for fragment amplification ...23
Table 3 – Genomic and plasmid fragments obtained by PCR ...25
Table 4 – Conditions used for ligation reactions. ...29
Table 5 – Primers used for sequence analysis when creating the transgenic GNE construct ...30
Table 6 – Sequence of primers used for generation of cDNA templates for Southern probes, annealing temperature used in each reaction and resultant amplification product. ...34
Table 7 – Information about anti-sense RNA probes used in whole-mount in situ hybridization of mouse embryos. ...37
Table 8 – Essential parameters necessary for generation of cDNA templates for Northern blot probes, either by restriction digestion or PCR amplification and correspondent annealing temperature, as well as the resultant amplification product ...42
Table 9 – Sequence of primers used for amplification of cDNA and resultant amplification product. .43 Table 10 – Real-Time PCR conditions used for transcript quantification. ...44
Table 11 – PCR amplification and cloning of fragments and confirmation of positive clones by restriction digest and sequence analysis. A – Genomic fragments obtained from 129/SVJ ES cell genomic DNA. B – Fragments obtained from plasmid DNA ...46
Table 12 – Cloning steps to assemble 5’ homology arm with β-galactosidase ...48
Table 13 – Cloning steps to assemble 3’ homology arm with a neomycin cassette ...51
Table 14 – Cloning steps to assemble the final construct ...52
Table 15 – Sample description and relevant information before cDNA synthesis ...64
Table 16 – Tukey-Kramer Multiple Comparisons Test on adult mouse tissue, from Real-Time PCR data (n=3; p=0.0001) ...65
Table 17 – Tukey-Kramer Multiple Comparisons Test on embryonic mouse tissues, from Real-Time PCR data (n=3; p=0.0001) ...67
X
L
IST OFA
BBREVIATIONSaa amino acids PCR Polymerase Chain Reaction
AS Allosteric site PEP Phosphoenolpyruvate
ATP Adenosine triphosphate PFA Paraformaldehyde
BM - RBC Bone Marrow - Red Blood Cells PL Placenta
BM + RBC Bone Marrow + Red Blood Cells QSM Quadriceps-sparing myopathy
bp, kbp, Mbp base pairs, kilo bp, Mega bp RN Normalized reporter
cDNA Complementary DNA RNA Ribonucleic acid
CIP Calf Intestine Phosphatase RT Reverse Transcriptase
CMP Cytidine monophosphate SD Standard deviation
CNS Central nervous system SPF Specific Pathogen Free
CT Cycle threshold T Tail
CTP Cytidine triphosphate Ta Temperature of annealing
Dec Decidua Tm Temperature of melting
DEPC Diethyl pyrocarbonate U Units
DIG Digoxigenin UTP Uridine triphosphate
DMRV Distal myopathy with rimmed vacuoles Vol Volume
DMSO Dimethylsulfoxide vs. versus
DNA Deoxyribonucleic acid WE Whole embryo
DTA Diptheria Toxin A WT Wild-type
E Embryonic day β-gal β-galactosidase
E11.5 M E11.5 Middle body portion
EC Enzyme commission codes
EDTA Ethylenediamine tetraacetic acid
EU European Union
Ex3 Exon 3
FCS Fetal calf serum
FHM Fetal human myoblast
GAPDH Glyceraldehyde-3-phophate dehydrogenase
gDNA Genomic DNA
GNE Uridine diphosphate-N-acetylglucosamine-2-epimerase/ N-acetylmannosamine kinase or UDP-GlcNAc 2-epimerase/ ManNAc kinase
H Head
HIBM Hereditary inclusion body myopathy
IBM Inclusion body myopathies
M Molecular weight marker
MCS Multiple cloning site
MEFC Mouse embryonic fibroblast cell
NCAM Neural cell adhesion molecules
Neo Neomycin
NES Nuclear Export Site
Neu5Ac 5-N-acetylneuraminic acid or sialic acid
Neu5Gc 5-N-glycolylneuraminic acid
OGS Oligosaccharides
OMIM Online Mendelian Inheritance in Man
p p-value
P Phosphate
p. Point mutation
PBS Dulbecco’s phosphate buffered saline
0
C
HAPTER
I
1
O
VERVIEWSialic acid is a key post-translational modifier in vivo, shown to be important in brain function, myogenesis and immune function. Production of sialic acid is regulated by UDP-N acetylglucosamine 2-epimerase/ N-acetylmannosamine kinase (GNE), a rate limiting dual function enzyme that metabolizes a critical step in the pathway to synthesize sialic acid in vivo. Over the last ten years, the role of GNE in many biological processes has become an important area of scientific investigation due to the finding that mutations leading to impaired GNE activity give rise to a debilitating hereditary myopathy. However, our understanding of GNE function, expression and its critical role in development and postnatal life requires further investigation. The main focus of this study is to address these issues; thus, the present review covers what has been discovered so far about sialic acid function and the biological importance of GNE.
I.1
B
IOLOGY OFS
IALICA
CIDSialic acid is a common term used in the description of N- and O-substituted derivatives of neuraminic acid (Wang, 2009), representing a unique monosaccharide determinant of numerous glycan-receptor connections involved in cell interaction and communication processes pertinent to health and disease (von Gunten and Bochner, 2008, Varki, 2007).
Structurally, sialic acids consist of a family of 9-carbon sugar acids with over 50 naturally occurring derivatives of neuraminic acid (Münster-Kühnel et al., 2004). This wide family can be divided in two branches, one of which forms N-acetylneuraminic acids (Neu5Ac), which are the most widespread form of sialic acid and almost the only form found in human glycoconjugates, including glycoproteins, glycosaminoglycans, gangliosides and mucins (Rosenberg, 1995, Schachter et al., 1997). The other branch is based on N-glycolylneuraminic acids (Neu5Gc) which is the type of sialic acid that most mammalian species use, including the mouse (Figure 1). However, humans do not have the ability to synthesize Neu5Gc (Chou et al., 1998) and this type of sialic acid is only present in low amounts in tumors such as colon and mammary carcinoma, gastric and liver cancer, malignant lymphoma and teratoma (Schauer et al., 1995).
Figure 1 – Conformation of N-acetylneuraminic acid (left) and N-glycoloylneuraminic acid (right) (Schauer et al.,
1995).
In mammalian cells, sialic acid monomers can be dimerised (disialic acid), oligomerised (oligosialic acid), or polymerized (polysialic acid) into linear, anionic chains consisting of α2-8-linked
C
HAPTERI
–
I
NTRODUCTION2 sialyl residues repeats. Polysialic acid is the major regulator of cell-cell adhesions and an unusual biological form of sialic acid that covalently modifies surface glycoconjugates on cells in a wide range of organisms (Wang, 2009).
I.1.1
C
ELLULAR ANDM
OLECULAR INTERACTIONSIn eukaryotic cells, modification of cell surface molecules with sialic acid is a frequent physiological event, where sialic acids are essential monosaccharide units of glycoconjugates. Sialic acids are usually encountered as terminal residues on cell surface glycoconjugates and due to their location and negative carboxylate function, sialic acids are very well suited as molecular determinants of specific biological processes (Varki, 1997). As a result, glycoproteins and glycolipids expressed at the cell surface can be modified to varying degrees by the addition of sialic acids, leading to specific sialylation patterns (Shauer et al., 1995). According to this, several remarkable differences are found within sialylation patterns of cells during mammalian development (Malicdan
et al., 2007), aging and oncogenesis (Shauer et al., 1995).
Sialic acid is an essential nutrient for several functions throughout the body, but it is particularly important for brain development, cognition and memory (Wang et al., 2009). It is significantly present in human diet and is an important component of human milk (Wang and Brad-Miller, 2003). Sialic acids are crucial in several cellular and molecular interactions, like regulating glycoprotein stability (Rens-Domiano and Reisine, 1991), controlling cellular interactions, including cell adhesion, migration and signal transduction (Keppler et al., 1999), modulation of surface recognition determinants (Varki, 1997) and hematopoietic cell differentiation (Keppler et al., 1999). In addition, sialic acids have been implicated in pathogenic processes such as inflammation (T and B cell activation) (Vestweber, 2003), adhesion of viruses or bacteria to target cells (Matrosovich and Klenk, 2003), metastasis formation and progression of a variety of tumors (Sawada et al., 1994).
I.1.2
S
IALIC ACIDD
ISTRIBUTIONSialic acids are found ubiquitously in all vertebrate cell types, tissues and body fluids, including saliva, gastric juice, serum, urine, tears and human milk. Free sialic acid is found in urine in very limited amounts (Montreuil et al., 1968). Sialic acids are found also present in the blood and their levels are used as a marker of the acute-phase response proteins. For example, fibrinogen and haptoglobin are two glycoproteins that significantly increase when exposed to cell injury (Taniuchi et
al., 1981). Sialic acids are found less frequently in prokaryotes, plants and invertebrates (Schauer,
2004, Troy, 1995).
In a study performed by Gagiannis and collaborators, in 2007, whilst trying to understand the changes in sialylation in heterozygous GNE-deficient mice, sialylation levels were also analyzed in detail for several organs in wild-type (WT) mice. By quantification of the membrane-bound sialic acid
3 concentration in each organ they found that although sialic acid was detected widely throughout the body, the levels seen in WT organs could be divided in two groups (Table 1): organs with high levels of membrane-bound sialic acid (40-60 nmol sialic acid per mg membrane), which included the brain, spinal cord, lung, spleen, blood cells and submandibulary gland; and organs with low levels of membrane-bound sialic acid (10-25 nmol sialic acid per mg membrane), represented by the heart, liver, kidney and abdominal muscle (Gagiannis et al., 2007).
Table 1 – Quantification of membrane bound sialic acid in several organs of a WT mouse. The values
represented correspond to nmol per mg of membrane (Adapted from Gagiannis et al., 2007).
The extensive distribution of sialic acids in addition to their structural versatility and peripheral position on oligosaccharide chains of glycoconjugates, allows them to be among the first molecules encountered by other cells or compounds coming into contact with the cell (Keppler et al., 1999). The characteristic distribution of sialic acids give rise to some outstanding examples, including the necessity for recognition of sialic acids by lectins of the inflammatory and immune response pathways (Vestweber and Blanks, 1999) or the particular sialylation of neural cell adhesion molecules (NCAM) that are responsible for the proper establishment of the vertebrate embryonic nervous system (Cunningham et al., 1983). In fact, NCAM is first expressed at mouse embryonic day 8 (E8), reaching maximal expression during the prenatal period, reflecting expression levels of polysialyltransferases and polysialic acid (Mühlenhoff et al., 2009). In addition, NCAM is heavily sialylated (Schwarzkopf, 2002), and plays a role in neuroblast migration, neurite outgrowth, axon pathfinding, fasciculation, synaptogenesis and synaptic plasticity (Mühlenhoff et al., 2009). Therefore, sialic acids are very important in neural function.
The highest levels of sialic acid are present within the mammalian central nervous system (CNS), mainly present in gangliosides and glycoproteins; only a small amount (around 3%) is found as free sialic acid within the CNS (Brunngraber et al., 1972). In fact, sialic acids are almost never seen in its free form in nature (Wang and Brand-Miller, 2003). During a study performed by Duncan and collaborators, in 2009, they speculated that due to the high concentrations of sialic acids necessary in the mammalian brain, an exogenous supply of this nutrient might be required, especially during
C
HAPTERI
–
I
NTRODUCTION4 growth and development (Duncan et al., 2009). This means that the sialic acid synthesized by the newborn, during the neonatal period, may not sufficient to all of the sialic acid requirements in the fast growing tissues; as a result, an exogenous source of sialic acid, for example in human milk, may contribute significantly to greater sialylation of gangliosides and glycoproteins in body fluids, tissues and brain glycoconjugates. However, the nutritional and biological roles of sialic acids in human milk and other dietary sources are still not fully understood (Wang and Brand-Miller, 2003).
I.1.3
S
IALICA
CIDB
IOSYNTHESISIn complement to dietary sources, all mammals have the capacity to synthesize sialic acid in every tissue from simple sugar precursors (Wang, 2009). Sialic acid biosynthesis is only possible due to a bifunctional enzyme, UDP-N-acetylglucosamine-2-epimerase/ N-acetylmannosamine kinase (hereafter designated as GNE enzyme), which catalyzes the two initial steps of sialic acid biosynthesis and is regulated by a downstream product, thus determining the extent of cell surface sialylation in cells.
In the mammalian system, this pathway begins with cytosolic glucose being converted in several steps into UDP-N-acetylglucosamine (UDP-GlcNAc), followed by five consecutive enzymatic steps until its final product is formed, 5-N-acetylneuraminic acid (Neu5Ac), the most common type of sialic acid. Figure 2 represents the biochemical pathway of sialic acid biosynthesis, which takes place in cellular cytoplasm. In the first committed step of the synthetic pathway, UDP-GlcNAc is converted to N-acetylmannosamine (ManNAc) and UDP by the action of UDP-GlcNAc-2-epimerase. This enzyme, the first portion of GNE enzyme, catalyzes an inversion of stereochemistry at C-2 of UDP-GlcNAc and hydrolysis of the glycosidic phosphate bond of the same sugar to give free ManNAc and UDP. As this first reaction is proven to be an irreversible, the enzyme is technically not an epimerase (Tanner, 2005), and this step is considered to be rate-limiting for sialic acid biosynthesis (Keppler et
al., 1999). The second step involves phosphorylation of ManNAc at position C-6 by
N-acetylmannosamine kinase (the second portion of GNE enzyme) to give ManNAc-6-phosphate (ManNAc-6-P), which in turn is converted to Neu5Ac-9-phosphate (NeuAc-9-P) by condensation with phosphoenolpyruvate (PEP) and catalyzed by Neu5Ac-9-phosphate synthase (Lawrence et al., 2000). The NeuNAc-9P is then dephosphorylated to generate Neu5NAc (sialic acid) (Hinderlich et al., 1997). The complexity of this pathway is amplified by the addition of ancillary epimerases and kinases, besides GNE epimerase and kinase activities (Huizing, 2004).
Afterwards, sialic acid is imported into the nucleus where it is converted to its activated form, cytidine monophosphate-N-acetylneuraminic acid (CMP-sialic acid), by the enzyme CMP-sialic acid synthetase and with the use of cytosine triphosphate (CTP) (Münster et al., 1998, Tanner, 2005). Although three decades have passed since the biosynthetic pathway of sialic acid was first described (Kean and Roseman, 1966), the precise subcellular localization of some key enzymes of this pathway is yet to be discovered. So far, only two enzymes have been characterized at the molecular level:
5 CMP-sialic acid synthetase is known to be almost exclusive to the cell nucleus (Münster et al., 2002), and GNE was found to be localized mainly in the cytoplasm, but also in the cell nucleus and Golgi complex (Krause et al., 2005).
Figure 2 – Sialic acid biosynthesis. The bifunctional enzyme UDP-GlcNAc 2-epimerase/ ManNAc kinase catalyzes
the first two steps in the biosynthesis of sialic acid. The epimerase part of the enzyme is feedback inhibited by the downstream product, CMP-sialic acid (dotted line). ATP - Adenosine triphosphate, PEP – Phosphoenolpyruvate, CTP – Cytidine triphosphate, OGS – Oligosaccharides (Galeano et al., 2007).
Once the CMP-sialic acid exits the nucleus into the cytosol it has two main functions. One of the roles of cytosolic CMP-sialic acid is to control GNE epimerase catalytic activity through a strong feedback inhibition by binding to its allosteric site (Seppala et al., 1999, Leroy et al., 2001), thereby contributing to the tight regulation of intracellular sialic acid production (Hinderlich et al., 1997). As CMP-sialic acid is proven to be an allosteric inhibitor of the GNE epimerase domain, previous sequestration of CMP-sialic acid in the nucleus may be necessary to prevent early inactivation of cytoplasmic GNE (Hinderlich et al., 1997, Stäsche et al., 1997).
On the other hand, the Golgi CMP-sialic acid transporter allows cytosolic CMP-sialic acid to enter the Golgi compartment, where CMP-sialic acid accomplishes its second role and is utilized as the substrate for Golgi-resident sialyltransferases in glycoconjugate biosynthesis (Harduin-Lepers et
C
HAPTERI
–
I
NTRODUCTION6
al., 2005). In this biosynthesis, sialyltransferases add CMP–activated sialic acid residues to specific
terminal non-reducing positions on oligosaccharide chains of proteins and lipids, leading to a differential expression in sialylation (Keppler et al., 1999). Finally, when sialylated glycoconjugates are no longer needed they are degraded within lysosomes, in which one of the first steps is the removal of sialic acid residues by the acidic enzyme neuraminidase (Sommar and Ellis, 1972).
In a similar fashion, exogenous sialic acid, from dietary intake, begins with endocytosis through the plasma membrane, where the endocytic vesicle is then delivered to the lysosomes in which bound sialic acid is removed by the same resident neuraminidase enzyme. Afterwards, sialic acid is transported out of the lysosomes and into the cell nucleus to be activated through conjugation to CMP; shortly after, sialic acid is activated to once again fulfill its main functions (Duncan et al., 2009).
I.2
UDP-N-
ACETYLGLUCOSAMINE2-
EPIMERASE/
N-
ACETYLMANNOSAMINE KINASEE
NZYMEI.2.1
GNE
GENE,
TRANSCRIPTS AND PROTEINSThe human GNE gene spans around 69.6 kbp (base pairs) of genomic DNA in chromosome 9p13.3 and contains 13 exons. The original human GNE gene is complemented by an additional 90 bp exon, named A1, located about 20 kbp upstream of exon 1. The splice variants result from alternative splicing of the exons A1, 1 and 2 producing different protein isoforms. So far, five different splice variants of human GNE mRNA have been described (Watts et al., 2003, Reinke and Hinderlich, 2007, Reinke et al., 2009, Tong et al., 2009). The mRNA transcript variant 1 encodes the longer protein (isoform 1) and is recognized in literature as GNE2. The mRNA transcript variant 2, on the other hand, is identified as GNE1. The other 3 mRNA transcript variants, specifically transcript variant 3, transcript variant 4 and transcript variant 5, encode proteins designated as GNE3, GNE4 and GNE5, respectively (NCBI Database).
Mouse GNE gene spans around 47.3 kbp of genomic DNA in chromosome 4qB1 and contains 12 exons (Figure 3, A and B). Alternative splicing of this gene results in two transcript variants encoding different isoforms. In mRNA transcript variant 1 the resulting protein (isoform 1, described as GNE1) has a shorter N-terminus compared to isoform 2. In contrast, the mRNA transcript variant 2 encodes the longer protein (isoform 2) and is identified as GNE2 in the scientific literature (Figure 3, C and D) (NCBI Database).
One of the splice variants in both species encode for the well-known GNE protein (GNE1) consisting of 722 amino acids (aa) (Lucka et al., 1999, Stäsche et al., 1997) and homology as high as 98.5% is seen between humans and mice. In other mammals, the amino acid sequences of GNE1 have similarities higher than 95%. Chicken protein GNE1 also reveals a high similarity of 93% compared to human GNE1, however, the similarity between human and zebrafish and pufferfishes GNE1 is only about 80% (Reinke and Hinderlich, 2007). In 2001, Eisenberg and collaborators
7 compared this human protein (GNE1) with its orthologs and came to the conclusion that no identity was found in Drosophila melanogaster, Caenorhabditis elegans or yeast. On the other hand, they found that GNE activity was present in bacteria and it was carried out by two independent enzymatic activities (Eisenberg et al., 2001). To this point, GNE is considered to be one of 40 genes that are exclusively shared between vertebrates and bacteria and it is also an example of horizontal transfer from bacteria to vertebrates (Salzberg et al., 2001).
Other splice variants contain open reading frames encoding for GNE proteins with modified N-termini, namely GNE2 (for mouse and human) producing 753aa long protein and GNE3 (only for human), containing 681aa. Sequence similarities were found between GNE2 proteins of human, ape, mouse (human-mouse similarities are up to 96.8%), rat and chicken, but no evidence of GNE2 was found in fish so far. Therefore, the presence of GNE2 seems to be evolutionarily conserved amongst mammals (Lucka et al., 1999, Reinke and Hinderlich, 2007) and the existence of GNE2 in chicken but not in fish leads to the conclusion of the emergence of a common ancestor after separation of the mammals and birds from fishes. The protein isoforms GNE3, GNE4 and GNE5 seem to be restricted to humans and so far they were not found in other species (Reinke and Hinderlich, 2007).
Figure 3 – GNE genomic locus, mRNA variants and protein isoforms in mouse. A – Chromosome 4 of Mus musculus (Ensembl: ENSMUSG00000028479) Scale bar: 10 Mbp. B – The mouse GNE gene structure. The
horizontal line represents intron sequence and the black boxes represent exons numbered below each box. Note there are two start codons (ATG) in the first and second exon. The total size of GNE locus is 47.3 kbp; Scale bar: 1 kbp. C – Two full length GNE mRNA transcript variants naturally occurring in mice (transcript variant 1 and transcript variant 2). The start codon of transcript variant 1 is present on exon 2, thus resulting in a shorter N-terminal portion in the corresponding protein GNE1. Scale bar: 100 bases (NCBI database). D –
C
HAPTERI
–
I
NTRODUCTION8
Schematic representation of GNE protein structure. The two different mouse protein isoforms (GNE1 and GNE2) are represented. The most important protein of GNE is a 722 amino acid bifunctional enzyme. The N-terminal GNE epimerase catalytic domain (aa 1-303) contains an allosteric site (AS, aa 263–266) and a putative nuclear export signal (NES, aa 121–140). The C-terminal portion of the enzyme harbors the GNE kinase catalytic domain (aa 410–722). Scale bar: 100 aa (Reinke and Hinderlich, 2007, Reinke et al., 2009, Huizing, 2004).
I.2.2
GNE
B
IOCHEMISTRY ANDP
OST-
TRANSCRIPTIONAL MODIFICATIONSGNE is a soluble protein and, as previously said, is mainly localized in the cytoplasm, but also in the Golgi complex and cell nucleus (Krause et al., 2005), although the role of this protein in the nucleus remains elusive. GNE is not predicted to undergo glycosylation, but it has several potential phosphorylation sites (Horstkorte et al., 2000). For this reason and because it is regulated at the gene expression level, GNE is controlled post-translationally by phosphorylation, which seems to depend on protein kinase C (Harms et al., 1973, Gagiannis et al., 2007), dimerization and hexamer formation to elicit both epimerase and kinase activities (Horstkorte et al., 1999).
Late in the 1990s, Hinderlich and colleagues noted that GNE in a dimer state was only able to phosphorylate ManNAc to ManNAc-6-P, whereas GNE in a tetrameric state was able to catalyze the epimerization of UDP-GlcNAc to ManNAc with release of UDP and subquent phosphorylation of ManNAc to ManNAc-6-P (Figure 2). They concluded that GNE enzyme could exist in two major oligomeric states, tetramers and dimers (Hinderlich et al., 1997). Around the same time, a mutational study of the rat enzyme showed that the N-terminal domain of the GNE enzyme bears the epimerase activity, whereas the C-terminal domain possesses kinase activity (Effertz et al., 1999). A decade later, in the study of Reinke et al., 2009, it was proven by different biophysical methods that protein GNE1 definitely exists either as a dimer or a tetramer, where the dimer state possesses only kinase activity and the tetrameric state has full enzyme activity (Reinke et al., 2009). Therefore, GNE consists of two functional domains, an UDP-GlcNAc 2-epimerase domain and a ManNAc kinase domain (Effertz et al., 1999). Despite the fact that these two domains could be expressed in separate, a strong intramolecular connection exists between the two domains (Blume et al., 2004), creating a unique bifunctional enzyme.
The UDP-GlcNAc 2-epimerase catalytic activity (EC 5.1.3.14) is located in the N-terminal portion of the protein (amino acids 1–303) and ManNAc kinase catalytic activity (EC 2.7.1.60) is in the
C-terminal portion (amino acids 410–722) (Effertz et al., 1999, Huizing, 2004). As it is possible to see
in figure 3 (D), the locations of all active sites within epimerase and kinase domains remain to be precisely determined (Harduin-Lepers et al., 2005). Up till now, the epimerase domain is known to have an allosteric site, within amino acids 263–266, where the downstream product of sialic acid biosynthetic pathway (CMP-sialic acid) can bind, leading to feedback inhibition of the epimerase activity (Seppala et al., 1991). This type of cellular regulation allows the stability and balance between the fully functional tetrameric state of the enzyme and the amount of sialic acid produced
9 (Ghaderi et al., 2007). Although both epimerase and kinase activities have been attributed to GNE enzyme, still little is known about the regulation, differential expression and subcellular localization of GNE in vivo.
I.2.3
GNE
EXPRESSIONA
NALYSISAlthough sialic acid distribution is widespread throughout the body and well studied, the production of sialic acid is restricted to cells possessing an active GNE enzyme.
In order to determine the expression of GNE enzyme at tissue level, several scientific groups have performed analysis on GNE mRNA transcripts and GNE protein through Northern blot and Western blot analysis, respectively. In 1999, Horstkorte et al. analyzed different adult mouse tissues and several embryonic stages during mouse development (figure 4). In the same year, Seppala et al. also analyzed the expression of GNE enzyme in some human tissues (figure 5). Both researchers came to the conclusion that GNE mRNA transcripts appeared to be in all tissues investigated, but with a large variety of expression levels. In the study of Horstkorte, the major transcript of the UDP-GlcNAc 2-epimerase detected was the size of 3.2 kb, with high expression detected in lung and kidney; however, the maximal expression was observed in the liver (Figure 4). Furthermore, during mouse embryonic development they also identified UDP-GlcNAc 2-epimerase transcript in the stages investigated (E7, E11, E15 and E17), but with no detectable differences between them. The authors did not investigate this point any further (Horstkorte et al., 1999). As for the study of Seppala et al. (1999), although the Northern blot analysis revealed a single band of >5.5 kb, GNE expression levels were also present in a variety of tissues, with highest level of expression of the human UDP-GlcNAc 2-epimerase in liver and placenta (Figure 5) (Seppala et al., 1999).
The published data has clearly shown that GNE is ubiquitously expressed, but is mainly present in the liver of both mice (Figure 4) and humans (Figure 5), with barely detectable transcript
Figure 4 – Northern blot analysis of mouse RNA Figure 5 – Northern blot analysis of human
for expression of GNE. Tissue analysis using 2 µg RNA for expression of GNE (Seppala et al., mouse mRNA per lane (Horstkorte et al., 1999). 1999).
C
HAPTERI
–
I
NTRODUCTION10 levels in the skeletal muscle compartment (Horstkorte et al., 1999, Seppala et al., 1999). In addition, analysis of the epimerase and kinase activities of GNE in mouse tissues showed that liver has the highest levels from the tissues examined, whilst skeletal muscle activities were negligible, which is also consistent with gene expression data(Stäsche et al., 1997).
A focused study by Duncan et al. (2009), using microarray gene expression data available on-line, established a developmental profile of GNE in several tissues; within the mouse liver they observed an increase of GNE expression right after birth and the highest expression in the weaning period, i.e., usually at 21 days post-partum (Figure 6) (Duncan et al., 2009). While studying the GNE expression in the central nervous system, the same authors considered it was worth noting that while membrane bound sialic acid was higher in brain tissue in comparison to the liver, however according to Gagiannis and collaborators (2007), the expression of GNE enzyme in the brain was relatively low in contrast to the liver; therefore, they suggested that the brain must depend on sialic acid synthesized in a different place in the mouse body (Duncan et al., 2009). However, the authors did not consider the fact that the brain has specialized regions that will certainly have different individual characteristics regarding sialic acid requirements and its biosynthesis. As a result, more studies are needed in this field in order to fully understand the detailed physiological impact, if any, of sialic acid biosynthesis produced in particular regions of the brain.
Figure 6 – Gene expression levels of GNE (sialic acid synthesis) in the central nervous system (CNS), liver and
small and large intestine throughout development and aging in the mouse. (E, embryonic; d, day; w, week; dashed line, not determined) (Adapted from Duncan et al., 2009).
GNE expression in the liver of rats and guinea pigs was also found to be very low right after birth and in the following days; after this first phase, an increase in GNE expression is detected corresponding to the weaning period, just as reported for mice (Gal et al., 1997). An interesting finding, once again in the study of Duncan et al. (2009), was that GNE epimerase activity increased at the same rate as the levels of 3’sialyllactose, an oligosaccharide present in milk. The authors suggested that a metabolic signal might exist between dietary sialic acid and liver GNE epimerase activity. Possibly, when milk sialic acid levels are very high, the endogenous sialic acid synthetic machinery is activated and therefore, by the time of weaning GNE expression is at its highest (Duncan et al., 2009). Similarly, Wang et al., in a study performed in 2007, demonstrated that GNE
11 expression increased in the liver and hippocampus of piglets, when these animals were provided with a diet supplemented with sialic acid (Wang et al., 2007).
Another interesting characteristic about GNE enzyme is that, apart from its main role in the sialic acid biosynthetic pathway, it may also control completely different signaling functions, such as sialyltransferase expression, ganglioside production, modulation of proliferation and apoptosis (Wang et al., 2006). As previously mentioned, GNE has a dynamic cellular localization, both in the cytoplasm and the cell nucleus (Krause et al., 2005). The nuclear localization, which is not required for ManNAc-6P production, suggests alternative roles of GNE, other than the sole pathway of sialic acid biosynthesis. Recently, several articles underlined this hypothesis and pursued novel functions for GNE. For example, in 2006, using a yeast two-hybrid matrix, Weidemann and colleagues proved that GNE was involved in protein-protein interactions with collapsin response mediator protein 1 and promyelocytic leukemia zinc finger protein (Weidemann et al., 2006).
I.3
S
IALIC ACID DISORDERS ANDM
UTATIONS
PECTRUM OFGNE
Nowadays, many disturbances affecting the metabolism of normal sialic acid biosynthesis are known; as a consequence, several rare disorders have been described, either caused by genetic defects, inborn errors or even environmental factors, that may be responsible for disease and, in some cases, lead to a premature death. For example, deficiencies of glycoproteins and gangliosides in the brain due to malnutrition in the first few months of life, where the brain is more actively accumulating gangliosides, are related to irreversible impaired learning behavior (McVeagh and Miller, 1997). More severe cases, like the accumulation of free sialic acid within the lysosomes, lead to diseases usually designated as free sialic acid storage disorders. Some examples of this group of disorders include Salla’s disease (Renlund et al., 1986) and infantile free sialic acid storage disease; both disorders are characterized by the same type of defects, however, in the latter deficiency is more pronounced leading to early death (Mancini et al., 1992).
For many years, scientists have known that different point mutations in the GNE gene can lead to two distinct human disorders, sialuria and hereditary inclusion body myopathy (HIBM). In a recent study by Kurochkina and collaborators, all human GNE missense mutations were mapped on a predicted three-dimensional model as well as active site residues of the UDP-GlcNAc 2-epimerase and ManNAc kinase domains of GNE enzyme, revealing that several mutations found in HIBM and sialuria patients are located proximal to proposed active sites of GNE (Kurochkina et al., 2010).
I.3.1
S
IALURIASialuria (MIM 269921) is a rare human metabolic disorder, also identified as a sialic acid storage disease, caused by failed feedback inhibition of GNE epimerase normal catalytic activity (Weiss et al.1989; Seppala et al.1991), where free sialic acid accumulates in the cell cytoplasm and
C
HAPTERI
–
I
NTRODUCTION12 large quantities of this compound are excreted in the urine (Thomas et al., 1985, Wilcken et al., 1987).
GNE mutation analysis
Until now, only seven people were described to have this disorder and just six were studied, all presenting identifiable mutations in GNE gene epimerase domain, most particularly in a short stretch of consecutive nucleotides that encode the amino acids 263 to 266 (Ferreira et al., 1999, Seppala et al., 1999, Aula and Gahl, 2001, Enns et al., 2001, Leroy et al., 2001). This disorder is caused by a binding defect of CMP-sialic acid, the downstream product of sialic acid biosynthesis, to UDP-GlcNAc 2-epimerase (the first portion of GNE enzyme) (Weiss et al., 1989). While the mutant epimerase continually produces free sialic acid, CMP-sialic acid can only inhibit the normal GNE epimerase, but not the mutant one, thus leading to a massive production and accumulation of free sialic acid (Seppala et al., 1999).
The localization of mutations, within GNE gene, in sialuria patients revealed that this genetic disorder is only caused by a missense mutation in one of two amino acids: either arginine at position 263 (R263L) or arginine at position 266 (R266Q; R266W), leading to speculation that the region within these codons is part of the allosteric site for CMP-sialic acid binding (see figure 3, D). In addition, the single mutation was found in only one allele of the GNE gene (Seppala et al., 1999). As a result, sialuria is an autosomal dominant disorder, where a single missense mutation is enough to cause the phenotype. However, in 2006, Penne and collaborators unintentionally discovered that a C303V mutation, in the GNE epimerase domain, produced a hyperactive enzyme with reduced feedback inhibition by CMP-sialic acid (Penner et al., 2006), similar to the ones found in sialuria patients. In another report, amino acids D255, E260, K268, and N275 were mutated, resulting in high levels of free sialic acid (Yarema et al., 2001). These studies led to the hypothesis that other possible mutations within the epimerase domain can result in the “sialuria phenotype” and that the exact dimensions of the allosteric site in GNE remain to be precisely determined (Kurochkina et al., 2010).
Clinical course and therapy
Sialuria is clinically characterized by variable and transient signs and symptoms, especially in infancy. Patients suffering from sialuria have variable degrees of developmental delay, coarse facial features, cytoplasmic accumulation and urinary excretion of large quantities (>1 g/day) of free sialic acid (Seppala et al., 1991, Leroy et al., 2001, Ferreira et al., 1999). Some sporadic clinical features include hepatosplenomegaly, delayed skeletal development, microcytic anemia and mild intellectual impairment. These clinical findings encountered in sialuria patients overlap the range of normal and so, despite very few sialuria cases have been reported, the prevalence of this disorder is probably underestimated. The long-term outcome of the disorder is unknown to date (Leroy et al., 2001).
13 A typical feature of free sialic acid storage disorders is the accumulation of sialic acid within the lysosomes, a characteristic that is not verified in sialuria, despite being considered a sialic acid storage disorder. The sialuria diagnosis is confirmed by the cytoplasmic, instead of lysosomal, distribution of free sialic acid (Aula and Gahl, 2001). Sialuria should be considered in the differential diagnosis of children with mild developmental delay and a close monitoring of liver and pulmonary function of these patients is recommended (Leroy et al., 2001).
At this time, only symptomatic therapy is available for sialuria patients but, perhaps the dietary ingestion of selenium, copper and zinc would help reduce the amount of free sialic acid accumulated in the cytoplasm, since these compounds are known to be inhibitors of GNE enzyme activity (Darvish et al., 2003). Two therapeutic approaches are currently being developed: one aims to decrease sialic acid levels, by providing CMP-sialic acid to cells to perform its function of allosteric inhibitor (Seppala et al., 1991), and the other one aims to decrease GNE activity at the protein level, by enzyme inhibitors (Blume et al., 2002), or at the gene level, by silencing of the dominant allele (Miller et al., 2003).
I.3.2
H
EREDITARYI
NCLUSIONB
ODYM
YOPATHYHereditary inclusion body myopathy (HIBM) also termed h-IBM, Inclusion Body Myopathy 2 (IBM2) or hereditary IBM quadriceps sparing type (OMIM 600737) is an autosomal recessive neuromuscular disorder caused by mutations in the GNE gene (Eisenberg et al., 2001). It is the same entity as Distal myopathy with rimmed vacuoles (DMRV) which was originally reported by Nonaka et
al. (1981) in Japanese patients, and thus it is also known as Nonaka Myopathy (OMIM 603824).
Initially described and delineated as separate myopathies, HIBM and DMRV, genetic analysis and the study of the muscle pathology revealed that these entities represent various stages in the natural course of the same disorder (Tomimitsu et al., 2004, Huizing, 2005, Huizing and Krasnewich, 2009).
GNE mutation analysis
HIBM was initially reported by Argov and Yarom (1984), in Jews of Persian descent who first described the disorder as a myopathy that primarily affects leg muscles, but with an unusual distribution that spares the quadriceps (Argov and Yarom, 1984). This particular pattern of weakness distribution is termed quadriceps-sparing myopathy (QSM). This major thigh muscle is markedly affected in many other forms of inclusion body myopathies and other degenerative myopathies. Almost a decade later, Sadeh and colleagues found that HIBM was not limited to Persian-Jews, but it was also found in individuals of Afghani, Iraqi and Egyptian descent (Sadeh et al., 1993).
Haplotype analysis around the GNE gene region of 104 affected people from 47 Middle Eastern families indicated one unique ancestral founder mutation in this community. The p.M712T mutation was the first mutation that incriminated GNE as the gene responsible for HIBM (Eisenberg
C
HAPTERI
–
I
NTRODUCTION14 mutations have been described. The second most common GNE mutation is p.V572L, predominantly identified in Japanese patients and some of Asian descent (Kayashima et al., 2002, Kim et al., 2006) and the third GNE founder mutation is p.D176V, mainly occurring in the Japanese population (Nishino et al., 2002).
Figure 7 – Schematic drawing representing GNE enzyme with major HIBM mutations (in red). Scale bar: 100 aa;
NES – Nuclear Export Site; AS – Allosteric site (Adapted from Sparks et al., 2005).
Over the last decade, the number of patients reported with HIBM from various backgrounds has significantly increased and, currently, 62 mutations have been described worldwide associated with HIBM (Huizing and Krasnewich, 2009). These GNE gene mutations were reported to occur in the regions encoding the epimerase domain (with the exception of the enzyme’s allosteric site), the kinase domain, or both, resulting in the same HIBM phenotype with sparing of the quadriceps (Eisenberg et al., 2002). A study by Sparks and colleagues revealed that the reduction in epimerase and kinase enzymatic activity is mutation-dependent and that mutations in one enzymatic domain also affect the activity of the other domain. This fact can explain the clinical similarities between patients with mutations on the epimerase domain and patients with mutations in the kinase domain (Sparks et al., 2005).
As this is an autosomal recessive disorder, two mutations are necessary to produce the HIBM phenotype. In addition, GNE mutations are spread throughout the GNE gene in one or in both alleles, producing a compound heterozygous or a homozygous genotype respectively. Most mutations found are missense, with a percentage of around 82%; however, 11% of all mutations reported so far are null, nonsense or frame shift mutations. These last types of mutations, which most probably result in RNA with limited or no protein coding capacity, where never encountered in both alleles of GNE (Huizing and Krasnewich, 2009). Therefore, the fact that none of the patients of any of the various ethnic origins carries two truncating mutations, together with the lethality associated with a knockout mouse of GNE (Schwarzkopf et al., 2002), raises the possibility that the complete absence of GNE enzyme is probably lethal. The study of GNE mutations also lead to another interesting discovery. According to Huizing and Krasnewich, GNE appears to have some “genetic hotspots” for mutation, i.e., a certain mutation that for some reason occurs independently in different families (Huizing and Krasnewich, 2009). One example of this occurrence is mutation R246Q that was found present in patients from Italy, the Bahamas and Taiwan (Eisenberg et al., 2001, Broccolini, et al., 2004).
15 Although most individuals with two GNE mutations will most likely display a HIBM phenotype, the penetrance of this disease is probably not 100%, as three individuals were identified in a populational study, who carried two disease-causing mutations and were asymptomatic at an advanced age (Argov and Soffer, 2003). Nevertheless, based on the increasing number of mutations reported just in the last decade, genetic analysis of GNE gene should be performed on patients presenting distal vacuolar myopathy (Grandis et al., 2010).
Molecular and cellular features
A unique feature of this disease is the progressive muscle weakness with sparing of the quadriceps muscles, partially or completely, even in the advanced stages of the disease (Argov and Yarom, 1984). Histologically, muscle fibers degenerate and develop cytoplasmic rimmed vacuoles and cytoplasmic or nuclear filamentous inclusions, measuring about 18–20 nm in diameter (Griggs et
al., 1995, Sivakumar and Dalakas, 1996).
The pathomechanism behind HIBM muscle degeneration is still a mystery. One possible hypothesis is the concept of hyposialylation, i.e., the decrease of sialic acid biosynthesis due to the reduced GNE enzyme activity, would lead to the muscle degeneration in HIBM patients, involving several glycans crucial for muscle normal functioning. Evidence of impaired sialylation of glycoproteins in the muscles of patients with HIBM was only demonstrated in 2004, by Huizing and colleagues. The group was able to prove that HIBM muscle displays decreased α-dystroglycan reactivity on its surface, probably due to impaired glycosylation. α-dystroglycan is a central protein of the skeletal muscle dystrophin–glycoprotein complex and when hypoglycosylated, is unable to stabilize this complex which in turn cannot maintain the structural stability of the sarcolemma during cycles of contraction and relaxation (Huizing et al., 2004). This protein is and it is also known to be involved in the pathomechanism of several congenital muscular dystrophies (Hayashi et al., 2001, Kobayashi et al., 1998). In contrast, Hinderlich et al. showed normal concentrations of membrane bound sialic acid in lymphoblastoid lines with the p.M712T mutation, the most predominant mutation found in HIBM patients (Hinderlich et al., 2004).
These results appear to be contradictory and also several inaccuracies can be detected; for example, in vitro studies of these primary cell lines derived from patients may not be representative because the culture medium used contains a high concentration of sialic acids. A transgenic mouse approach would prove to be a more adequate model to analyze the tissues in the appropriate endocrine environment.
Clinical course and therapy
Hereditary inclusion body myopathy (OMIM 600737) is a neuromuscular disorder that starts to affect individuals from 15 years of age up to 40 years, with an average onset of 26 years (Galeano