Universidade de Trás-os-Montes e Alto Douro
Desenvolvimento de equações de volume em florestas
tropicais:
Ensaio de metodologia de agrupamento de dados para as espécies
de Timor-Leste
Dissertação de mestrado em Engenharia Florestal
Leandro Pereira Dias
Teresa de Jesus Fidalgo Fonseca
i Dissertação para obtenção do grau de Mestre em Engenharia Florestal pela Universidade de Trás-os-Montes e Alto Douro.
iii
Resumo
A gestão florestal sustentável está dependente das ferramentas de que os gestores florestais disponham para tomar decisões. As equações de volume individual das árvores surgem como uma das ferramentas mais importantes para a silvicultura, permitindo que as decisões que envolvem a quantificação do material lenhoso e da componente mercantil do tronco sejam tomadas com maior certeza e eficácia.
A disponibilidade de equações volumétricas por espécie é a situação ideal. No entanto, no caso de florestas tropicais, o elevado número de espécies diferentes e o escasso número de observações por espécie pode tornar impraticável desenvolver equações individualizadas para todas as espécies, havendo necessidade de as agrupar de modo a poderem estabelecer-se equações gerais com o menor erro possível.
A técnica de agrupamento pode fazer-se por género, família, critérios ecofisiológicos ou por análise estatística multivariada, que já mostrou ser bastante satisfatória noutros estudos. O estudo que se apresenta respeita ao desenvolvimento de equações de volume para espécies da floresta timorense, tendo como suporte os dados colhidos aquando do I Inventário Florestal de Timor-Leste. O inventário teve lugar em dois distritos timorenses, o distrito de Bobonaro e o distrito de Covalima, resultando na listagem de mais de 150 espécies florestais diferentes em cerca de 5000 observações feitas.
O desenvolvimento deste trabalho permitiu a criação de dois modelos de regressão para estimar o volume das espécies timorenses isoladamente ou em grupos. A tarefa de encontrar uma relação entre o crescimento e o local de origem foi também bem sucedida, obtendo-se para algumas das espécies uma equação que pode ser aplicada sem que se faça a distinção entre local de proveniência. Apesar de haver pouca informação e não haver uma forma mais correta de fazer o agrupamento de espécies, a metodologia aplicada mostrou resultados encorajadores e repartiu aquelas espécies incapazes de fundamentar uma equação em dez grupos homogéneos para os quais se estipulou um modelo geral e com bom ajuste estatístico.
Palavras chave: Inventário florestal, floresta tropical, agrupamento de espécies, análise
v
Abstract
Sustainable forest management depends on the available tools that forest engineers have in order to make decisions. The volume equations appear on the top of the tools that are most important to the silvicultural practices, allowing that the decisions may be taken with more efficiency and certain.
The development of volume equations by specie is possible, in fact, it’s the ideal situation. However, considering tropical rain forests, the huge number of different species and the insufficient number of observations for each one of the species can make impractical build equations for each one. The solution for this is the species grouping.
Taxonomic features, physiologic characteristics and statistical analysis are examples of methods used by different authors to aggregate species into homogeny groups. Statistical analysis showed encouraging results in other studies.
The work presented goes towards the development of volume equations for the tropical forest of Timor. It´s supported by the data collected in the first forestry inventory made in the country. It took place in two districts of Timor-Leste, Bobonaro and Covalima, resulting in a list around 5000 observations and 150 different species.
The development of this academic report was able to create two regression models that can predict tree volume by species or groups. The task to find a close relation between the yield and the location was also successful. It was possible to obtain, for some of the species that were comun between districts, one equation that can be used without destintion of local of origin.
Although theres not many information and a correct way of grouping species the methodology applied in this report as showed remarkable results and was able to divide those species with fewer observations into ten homogeneous groups for wich was estipulated a general equation with a good performance.
Keywords: Forest management, tropical rain forest, species grouping, Statistical
vii
Índice geral
Resumo ... iii Abstract ... v Índice de figuras ... ix Índice de quadros ... xi 1. Introdução ... 152. Desenvolvimento de equações de volume para espécies florestais ... 19
2.1. Modelos comuns de regressão ... 20
2.2. Aplicação de uma variável binária para agrupar dados de uma espécie provenientes de locais distintos ... 22
2.3. Metodologia para o agrupamento de espécies ... 23
2.3.1 Agrupamento pela taxonomia da espécie ... 24
2.3.2 Agrupamento pela ecofisiologia da espécie ... 24
2.3.3 Agrupamento através do fim comercial... 24
2.3.4 Agrupamento por análises estatísticas ... 25
3. Material e métodos ... 29
3.1. Descrição do material ... 29
3.2. Tratamento da base de dados ... 36
3.3. Agrupamento de dados de ambos os distritos ... 37
3.3.1 Espécies com pelo menos 30 observações em cada distrito ... 37
3.3.2 Espécies com pelo menos 60 observações em ambos os distritos ... 37
3.3.3 Restantes casos ... 38
3.4. Aplicação de uma variável binária para estudo da influência do local ... 39
3.5. Estimação de modelos para as espécies menos representadas (agrupamento por “clusters”) ... 42
viii
3.6.1 Análise do coeficiente de determinação ... 46
3.6.2 Análise do comportamento dos resíduos ... 47
3.6.3 Retransformação do desvio padrão dos resíduos para as unidades originais e percentagem e aplicação de um fator de correção aos modelos logaritmizados. ... 48
4. Resultados ... 51
4.1. Avaliação da influência do local para as espécies comuns ... 51
4.2. Aplicação de modelos volumétricos ... 54
4.3. Agrupamento de espécies pelo método de “clusters” ... 66
5. Discussão de resultados ... 75
ix
Índice de figuras
Figura 1 - Mapa da localização do território timorense e dos dois distritos inventariados que fundamentam este relatório (Fonte: www 1). ... 16
Figura 2 - Representação gráfica do padrão esperado para duas espécies mediante a aplicação das equações de volume relativas aos modelos 18 e 19. ... 40
Figura 3 - Representação gráfica do padrão esperado para duas espécies mediante a aplicação da equação de volume relativa ao modelo 20. ... 41
Figura 4 - Representação gráfica do padrão esperado para duas espécies mediante a aplicação da equação de volume relativa ao modelo 21. ... 41
Figura 5 - Diagrama do processo de separação das espécies de acordo com as
xi
Índice de quadros
Quadro 1 - Compilação de modelos volumétricos utilizados na estimação do volume do tronco de espécies florestais (Adaptado de Fonte, 2000). ... 21
Quadro 2 - Resumo das metodologias usadas por vários autores no agrupamento de espécies florestais (Adaptado de Akindele, 2005)... 27
Quadro 3 - Sumário da distribuição das espécies por número de observações no terreno. ... 30
Quadro 4 - Resumo da distribuição das observações por classe de diâmetro para as espécies do distrito de Bobonaro (≥ 30 observações)... 31
Quadro 5 - Resumo da distribuição das observações por classe de diâmetro para as espécies do distrito de Covalima (≥ 30 observações). ... 33
Quadro 6 - Resumo da distribuição das observações por classe de diâmetro para as espécies do distrito de Covalima (≥ 30 observações). Continuação... 35
Quadro 7 - Resultados do efeito da proveniência com recurso à aplicação da variável binária para as espécies comuns a ambos os distritos. ... 53
Quadro 8 - Resultados da aplicação de um modelo de regrassão linear simples (Eq. 2) sem diferenciação da proveniência dos dados para as espécies comuns aos dois distritos e que mostraram não haver influência do local no seu comportamento. .. 55
Quadro 9 - Resultados da aplicação de um modelo de regressão múltipla (Eq. 3) sem diferenciação da proveniência dos dados para as espécies comuns aos dois distritos e que mostraram não haver influência do local no seu comportamento. ... 57
xii Quadro 10 - Aplicação do modelo de regressão linear simples (Eq. 2) para as espécies
de Bobonaro que falharam o teste de influência de proveniência e as restantes com mais de 30 observações. ... 59
Quadro 11 - Aplicação do modelo de regressão múltipla (Eq. 3) para as espécies de Bobonaro que falharam o teste de influência de proveniência e as restantes com mais de 30 observações. ... 59
Quadro 12 - Aplicação do modelo de regressão linear simples (Eq. 2) para as espécies de Covalima que falharam o teste de influência de proveniência e as restantes com mais de 30 observações. ... 61
Quadro 13 - Aplicação do modelo de regressão múltipla (Eq. 3) para as espécies de Covalima que falharam o teste de influência de proveniência e as restantes com mais de 30 observações. ... 63
Quadro 14 – Aplicação do modelo de regressão linear simples (Eq. 2) para as espécies sem identificação ou desconhecidas. Considera exemplares de ambos os distritos. ... 65
Quadro 15 - Aplicação do modelo de regressão múltipla (Eq. 3) para as espécies sem identificação ou desconhecidas. Considera exemplares de ambos os distritos. ... 65
Quadro 16 - Distribuição das espécies pelos diferentes grupos após aplicação da
metodologia de “clusters” (Grupos 1 a 5). ... 67
Quadro 17 - Distribuição das espécies pelos diferentes grupos após aplicação da
metodologia de “clusters” (Grupos 6 a 8). ... 69
Quadro 18 - Distribuição das espécies pelos diferentes grupos após aplicação da
metodologia de “clusters” (Grupos 9 e 10). ... 71
Quadro 19 – Resultados da aplicação do modelo de regressão linear simples (Eq. 2) para os grupos recém formados com as espécies menos representadas. ... 73
xiii Quadro 20 - Resultados da aplicação do modelo de regressão múltipla (Eq. 3) para os
15
1.
Introdução
As florestas tropicais são responsáveis por 47% de toda a área florestal do planeta, apresentando um sem número de espécie vegetais e animais que dependem do seu bioma (FAO, 2003; FAO, 2012; Richards, 1996; www2). Toda esta riqueza tem sido largamente explorada pelo Homem na ânsia pelos seus recursos. Estima-se que a floresta tropical esteja a ser derrubada a um ritmo de 127.300km2 por ano por culpa do seu riquíssimo valor madeireiro e consequência de uma sobrexploração e de uma ilegalidade nos processos desta mesma exploração (FAO, 1999).
Numa era de globalização, de crescimento populacional e industrial, o papel que estes maciços florestais representam na manutenção do equilíbrio global é enorme. E a sua destruição faz apenas com que a capacidade das florestas para combater o aquecimento global e a fixação de carbono seja decrescida (Köhler e Huth, 1998). E não é apenas esta capacidade que se perde mas também se verifica uma diminuição na qualidade e quantidade de habitats de espécies, resultando na extinção destas (Tucker e Townshend, 2000).
Por isto mesmo, o Ministério da Agricultura, Florestas e Pesca da República Democrática de Timor-Leste preparou um documento onde foi feita uma análise ao setor florestal (“Prioridades e proposta de investimento sectorial”), no qual se previa a necessidade de realizar um Inventário Florestal Nacional (IFN) e a avaliação rigorosa dos recursos existentes. O primeiro IFN foi realizado entre 2008 e 2010 em dois distritos: Bobonaro e Covalima (Figura 1) e permitiu um primeiro cadastro florestal e setorial (Marques et al., 2010). Para suporte à avaliação das existências florestais, ao nível do volume, foi realizada a colheita de dados de volume em árvores em pé, num conjunto numeroso de árvores (cerca de 5000). Com base na informação recolhida, Marques et al. (2010) desenvolveram um conjunto inicial de equações de volume, para a porção mercantil do tronco, numa primeira fase para as árvores amostradas em Bobonaro e, posteriormente para as inventariadas em Covalima.
Para a inventariação dos recursos e como suporte à gestão florestal, é extremamente útil a existência de funções matemáticas que possam dar informação sobre os padrões de crescimento deste tipo de floresta e que permita ao gestor florestal tomar conhecimento da quantidade e do valor da madeira (Akindele et al., 2006).
16
Figura 1 - Mapa da localização do território timorense e dos dois distritos inventariados que
fundamentam este relatório (Fonte: www 1).
No entanto, tal como referem autores como Akindele e LeMay (2006), Vanclay (1991), Leech et al. (1991), a enorme diversidade de espécies associada às florestas tropicais torna-se um problema aquando do estabelecimento de equações de volume. Este facto complica a tarefa de sumariar toda a informação e de a aplicar num estudo científico (Phillips et al., 2002) dada a complexidade de hábitos de crescimento e de interações deste ambiente.
Segundo Akindele (2005), existem três possibilidades para se desenvolverem equações de volume para as espécies das florestas tropicais: (i) usando a informação individual de cada espécie para criar a sua própria equação de volume; (ii) empregando toda a informação colhida sobre todas as espécies inventariadas e desenvolver um conjunto de equações gerais ou ainda; (iii) dividir as espécies por grau de semelhança e colocá-las num grupo para o qual se estabelece uma equação que será aplicável a todas as espécies nele contidas. Claro que a situação ideal seria a primeira, na qual cada
17 espécie teria a sua própria equação, mas as circunstâncias dadas pela floresta tropical impossibilitam essa situação (Akindele e LeMay, 2006).
A bibliografia consultada para a elaboração deste trabalho sustenta que a hipótese mais apropriada é criar grupos de espécies com caraterísticas de crescimento semelhantes, sendo impraticável a criação de uma equação específica para cada espécie (Vanclay, 1991; Akindele e LeMay, 2006; Akindele, 2005; Leech et al., 1991). E apesar do tema do agrupamento de dados já ter sido abordado por vários autores, não existe uma abordagem universal que possa ser aplicada para fazer o agrupamento de espécies (Gitay e Noble, 1997). Existem, no entanto, algumas formas para agrupar as espécies com base em diversas caraterísticas das plantas. Através da sua taxonomia, que consiste em juntar espécies pelos níveis género ou família; com base nos comportamentos ecofisiológicos, tais como fotoperíodo, tolerância à sombra, vigor da regeneração natural ou capacidade potencial de atingir os níveis mais elevados; baseada no seu valor e utilização comercial, ou seja, apoiada nas suas propriedades físicas e mecânicas; e ainda, suportada em análises estatísticas, como a aplicação de “clusters”, que funciona através da análise do grau de associação entre dois objetos, neste caso espécies, juntando-os no mesmo grupo se as diferenças forem mínimas ou separando-os caso contrário. Desde logo, surgem críticas a alguns dos métodos por parte dos autores da bibliografia consultada para este trabalho, não estando definido um método que seja adequado a todas as situações.
Esta dissertação dá continuidade ao trabalho desenvolvido por Marques et al. (2010) e surge na perspetiva de desenvolver equações de volume do fuste, sobre a casca, para as espécies suficientemente representadas nas parcelas inventariadas nos distritos de Bobonaro e de Covalima tendo como suporte o Inventário Florestal realizado em Timor-Leste. É ainda objetivo ensaiar métodos para agrupar as espécies por forma a desenvolver equações mais gerais e que permitam um bom ajustamento aos dados para as espécies menos representadas. Estas espécies foram inicialmente agrupadas, em cada distrito, com base na taxonomia, segundo o género ou a família ou ainda num conjunto diverso que foi designado por “outras espécies” (Marques et al., 2010).
É intenção do trabalho criarem-se equações volumétricas que abranjam não apenas uma região associada a um distrito mas também outras regiões que se apresentem como semelhantes na influência que exercem no desenvolvimento das espécies. Se verificada esta situação poderá dispor-se de uma mesma equação para aquelas espécies florestais que lhes sejam comuns, diminuindo significativamente a
18 quantidade de dados a recolher pelas equipas de pesquisa e de trabalho em situações futuras (Brandeis et al., 2005).
Em suma, os objectivos deste trabalho são:
a. Desenvolvimento de equações de volume com casca para as espécies com informação suficiente para suportar um bom ajustamento;
b. Avaliação de possíveis semelhanças volumétrica para a mesma espécie nos dois distritos inventariados com vista à construção de uma mesma equação de volume;
c. Síntese e ensaio de metodologias de agrupamento de espécies e identificação da (s) mais adequada (s);
d. Desenvolvimento de equações de volume para as espécies com informação insuficiente, quanto à respetiva representatividade, e/ou identificação, para as quais não se pôde recorrer ao método de agrupamento referido em c.
19
2.
Desenvolvimento de equações de volume para espécies
florestais
A utilização de equações de volume ou modelos de regressão é bastante comum aquando da quantificação do material lenhoso de determinada mancha florestal. Este processo envolve a estimativa de uma variável dependente, geralmente o volume, em função de uma ou mais variáveis independentes que estejam relacionadas com esta (Marques et al., 2007).
Assim sendo, torna-se importante que este processo de cálculo seja simplificado, sem prejuízo da sua precisão, de modo a responder às necessidades dos gestores florestais (Fonte, 2000). Visto que a gestão florestal envolve um investimento considerável de recursos económicos e logísticos, faz todo o sentido que seja programada de forma eficiente, por forma ao investidor recuperar o que foi aplicado. Os modelos matemáticos desenvolvidos têm um papel importante nesse aspeto, ajudando na predição dos acontecimentos, fornecendo hipóteses de gestão. Estando, é claro, dependente da existência de informação adequada (Vanclay et al., 1993).
No caso específico deste trabalho recorreu-se à aplicação de modelos de regressão que estimam o valor do volume individual de uma árvore com base em duas variáveis independentes, ou mais propriamente, dois regressores, neste caso o diâmetro a 1,30 metros (d) e a altura do fuste até aos primeiros ramos vivos (hbc).
20
2.1.
Modelos comuns de regressão
Como modelos mais comuns nas análises de regressão estatística, para determinação do volume, temos os modelos lineares simples, lineares múltiplos e os não lineares.
Os modelos de regressão lineares simples ou de simples entrada estudam a relação linear entre duas variáveis X e Y, sendo a primeira a variável independente ou regressor e a segunda a dependente ou resposta (Fonseca, 2009). Aplicando ao exemplo florestal e ao problema estudado traduzir-se-ia na relação entre o diâmetro a 1,30 metros e o volume. Estes possuem a vantagem de serem fáceis e rápidos de elaborar, o que os torna economicamente mais atractivos. No entanto, induzem o pressuposto de que todas as árvores com determinado diâmetro possuem a mesma altura e forma, criando-se um afastamento à realidade (Marques et al, 2007).
Como alternativa aos modelos simples temos os modelos de regressão múltipla que combinam mais que uma variável independente para estimar o comportamento da variável dependente (Fonseca, 2009). Com a aplicação deste tipo de modelos pode advir um incremente da precisão na estimativa da variável resposta e é deixado de parte o erro de considerar que todas as árvores da mesma classe de diâmetro sejam iguais em forma e tamanho, como era feito nos modelos anteriores. Este aumento é tanto maior quanto mais heterogénea for a estrutura da floresta em estudo (Fonte, 2000).
Surgem ainda como opção os modelos não lineares, que frequentemente diferem dos lineares apenas na forma de apresentação dos seus parâmetros. Associada a estes modelos temos a grande capacidade de tradução a formas distintas o que se torna vantajoso na tradução do modelo face à realidade (Fonseca, 2009).
Marques et al. (2010) utilizaram a relação alométrica geral no ajustamento das equações de volume para os dados de Bobonaro e de Covalima (Equação 1), onde v representa o volume do tronco desde a base até à base da copa.
21 Dos modelos mais referidos na bibliografia, pelo bom desempenho que apresentam, constam os modelos de Spurr e de Shumacher-Hall, cujas formas linearizadas estão indicadas nas equações 2 e 3. A equação 2 corresponde a um modelo cuja forma funcional coincide com um modelo de regressão linear simples e a equação 3 a um modelo de regressão múltipla (Equações 2 e 3).
log v = β0 + β1×log (d2hbc) (2)
log v = β0 + β1×log (d) + β2× log (hbc) (3)
No quadro 1 apresenta-se uma síntese de outros modelos que têm sido propostos e utilizados por diversos autores para estimação do volume das árvores.
Quadro 1 - Compilação de modelos volumétricos utilizados na estimação do volume do
tronco de espécies florestais (Adaptado de Fonte, 2000).
Modelo Referência Equação
4 Kopezky-Gehrhardt = + 5 Dissescu-Meyer = + 6 Berkhout = 7 Brenac = 8 Shumacher-Hall (1933) = ℎ 9 Spurr (1952) = ℎ 10 Spurr (1952) = ( ℎ) 11 Dwight e Thornber = ℎ( ) 12 Spurr (1952) = + ℎ 13 Honer (1965) = + ℎ + ℎ 14 Ogaya (1968) = + ℎ 15 Goulding e Murray (1976) = ℎ + ℎ 16 Buckhart (1977) = + ℎ + ℎ 17 Akindele (2005) = + ℎ
22
2.2.
Aplicação de uma variável binária para agrupar dados de uma
espécie provenientes de locais distintos
Como já foi enunciado na secção 1, um dos objetivos para este trabalho é o desenvolvimento de equações que possam traduzir o comportamento do volume para determinada espécie, ou grupo de espécies, para mais que uma região/distrito de Timor-Leste. Isto não só significaria uma poupança de recursos e de dinheiro, aquando da recolha dos dados de campo, como também um tratamento de dados mais simples e mais rápido no gabinete.
Neste campo, a utilização de variáveis binárias surge como uma solução matemática para a avaliação da possibilidade de considerar que dados colhidos nos distritos timorenses de Bobonaro e Covalima podem ser conjugados. O processo pode ser aplicado a qualquer das formas funcionais já descritas, procedendo-se à generalização do (s) modelo (s) seleccionado (s) para o efeito adicionando-se uma variável binária referente à localização. Esta variável deve ser incorporada de forma a permitir contemplar diferenças ao nível da ordenada na origem e da inclinação da reta ou, no caso de modelos de regressão linear múltipla, da superfície traduzida pelo modelo.
23
2.3.
Metodologia para o agrupamento de espécies
Do trabalho de campo realizado no IFN nos dois distritos timorenses resultou uma base de dados composta por uma elevada quantidade de espécies e um conjunto numeroso de observações. Esta secção tem como objetivo descrever o que foi feito por forma a organizar essa informação com vista aos objetivos finais.
Tal como a muitos outros autores que outrora trabalharam com dados provenientes de floresta do tipo tropical, surgiram diversos problemas que dificultam o processamento da informação. Um desses problemas advém exatamente do grande número de espécies inventariadas que origina uma distribuição irregular das espécies ou uma presença pouco pronunciada que leva a uma insuficiência ou deficiência dos dados para se desenvolverem modelos assertivos (Vanclay, 1991; Phillips et al., 2002). Tendo este facto em conta, e de que é impraticável desenvolver uma equação individual para cada uma das espécies inventariadas, a melhor solução é o agrupamento das espécies (Vanclay, 1991; Akindele, 2005; Akindele e LeMay, 2006).
O agrupamento consiste em colocar as espécies em grupos homogéneos através da análise de um qualquer parâmetro que as distinga ou aproxime, com o intuito de que o número de equações específicas diminua ao ponto de se ficar apenas com um número de modelos manejável (Vanclay, 1991; Akindele, 2005) (Quadro 2).
Autores como Lischke et al. (1996) questionam qual será o nível mínimo de observações para o agrupamento de modo a retratar a enorme variabilidade de uma floresta, não havendo, para já, uma resposta concreta. E existe também a dúvida de qual será a melhor metodologia para realizar o agrupamento (Gitay e Noble, 1997). Akindele (2005) enumera alguns métodos de agrupamento, como o agrupamento pela taxonomia, pelo comportamento ecofisiológico, aplicação comercial, análise estatística, forma e ou adelgaçamento do tronco e ainda através das caraterísticas do solo. Estes aspetos serão apresentados de forma individualizada nas subsecções seguintes.
24
2.3.1
Agrupamento pela taxonomia da espécie
O agrupamento taxonómico consiste numa distribuição das espécies por grupos onde o critério de distinção é a classificação ao nível da família ou género. Este tipo de metodologia é, em geral, usada nos estudos de biodiversidade mas pode ser uma das abordagens a tomar no agrupamento de espécies para outros fins. No entanto, este método não agrada a alguns autores por não ser o ideal para retratar os padrões de crescimento e produção das espécies florestais (Vanclay, 1991; Phillips et al., 2002).
2.3.2
Agrupamento pela ecofisiologia da espécie
Este tipo de agrupamento tem como critérios de separação os comportamentos ecofisiológicos referentes à espécie, seja a exigência de radiação solar, a tolerância à sombra, necessidade de clareiras, capacidade e vigor da regeneração natural, capacidade de atingir o topo do copado, etc. Tal como o método anterior, surgem problemas na sua aplicação. Akindele (2005) refere que para muitas das espécies florestais tropicais carece a informação relativa a estes mesmos critérios de separação.
2.3.3
Agrupamento através do fim comercial
Outra das metodologias usada para classificar e agrupar espécies florestais em alguns países tropicais é baseada na possível aplicação da madeira que cada espécie teria quando chegasse ao transformador ou ao mercado. Muitas vezes, este tipo de classificação era baseado nos valores de mercado da época para o tipo de madeira em questão. Mais uma vez, o método não era o mais correto, pois considerava apenas as propriedades da madeira e os seus usos e não os hábitos de crescimento da espécie (Akindele, 2005).
25
2.3.4
Agrupamento por análises estatísticas
Uma outra hipótese considera a agregação de espécies por grupos homogéneos em que o critério de distribuição assenta em parâmetros estatísticos ou num processo matemático que as una por grau de semelhança. Isto pode ser realizado através da aplicação de análise de “clusters”, análise discriminatória, etc. Com este tipo de abordagem ao problema, parecem ser ultrapassadas as dificuldades ou fraquezas que os outros métodos de agrupamento apresentavam, visto que neste caso em concreto é assegurado que as espécies são comparadas e agrupadas com base no seu comportamento de crescimento e/ou produtivo (Akindele, 2005).
O método de agrupamento por análise de “clusters”é um procedimento de estatística multivariada que organiza um conjunto de dados conhecidos em grupos homogéneos. Este processo envolve a análise dos dados e o seu agrupamento tomando em conta apenas as semelhanças entre eles, sem definir critérios de agrupamento (Reis, 2001).
A grande questão deste processo é a definição do critério de partição que levará à formação dos grupos e à separação das espécies, já que diferentes abordagens poderão levar a resultados diferentes sem que nenhum esteja errado. É, por isso, importante ter especial atenção na escolha da variável de comparação para resultados coerentes. Como refere Reis (2001), os métodos são exploratórios e estamos a gerar hipóteses de abordagem.
O estudo das variáveis dendrométricas do tronco é muito comum na avaliação de volumes e em outros estudos biométricos. No entanto, as variáveis mais frequentemente disponíveis são o diâmetro a 1,3 metros e a alturas, total ou do fuste, as quais, se consideradas isoladamente, não são as mais adequadas ao ensaio de critérios de partição. Segundo Duarte (2001), a forma corresponde ao perfil do tronco e a sua forma geométrica, e a taxa de variação do diâmetro ao longo do tronco corresponde ao adelgaçamento do mesmo. Combinados, estes dois conceitos formam um único, o perfil do tronco.Como hipótese de agrupamento de espécies surge, neste trabalho, o recurso ao quociente de forma do tronco, isto é, à razão entre dois diâmetros avaliados a alturas diferentes do perfil do tronco.
27
Quadro 2 - Resumo das metodologias usadas por vários autores no agrupamento de espécies florestais (Adaptado de Akindele, 2005).
Método Local Autores
Teste F para amostras emparelhadas Floresta tropical australiana Vanclay (1991)
Agrupamento ecofisiológico Floresta mista da Malásia Chai e LeMay (1993)
Agrupamento ecofisiológico Floresta tropical da Malásia Kohler et al. (2000)
Teste F para amostras emparelhadas (modificado) Floresta tropical da Guiana Francesa Gourlet-Fleury e Houllier (2000)
Análise por “clusters” e análise discriminante Floresta tropical mista do Borneo Phillips et al. (2002)
Análise por “clusters” e minimização de erros de agregação Floresta tropical da Guiana Francesa Picard and Franc (2003)
Análise por “clusters” Floresta mista no estado de Mississipi Zhao et al. (2004)
29
3.
Material e métodos
3.1.
Descrição do material
A base de dados que sustenta este trabalho foi colhida, como referido, no decorrer do 1º Inventário Florestal de Timor-Leste, no distrito de Bobonaro, em 2008, e no distrito de Covalima, em 2009.
Para a inventariação dos dois distritos usou-se uma amostragem em vários graus ou subamostragem, distribuída sistematicamente numa grelha de 4×4 km2 (Marques et
al., 2010). Nas parcelas de estudo procedeu-se à enumeração das árvores vivas e à
medição de parâmetros dendrométricos. Em todas as árvores com altura igual ou superior a 1,30m foi medido o diâmetro a esse nível. Num subconjunto, por espécie e por classe de d, foram realizadas avaliações complementares das variáveis: altura do fuste (altura até à zona do tronco onde se encontram os primeiros ramos vivos), diâmetro a esse nível e altura total. Estas avaliações foram realizadas com o relascópio de Bitterlich, modelo CP. Foi ainda realizada a avaliação da espessura da casca nas árvores amostra. O procedimento encontra-se detalhado em Marques et al., 2010.
Como resultado, em ambos os distritos, foram inventariadas cerca de 150 espécies, para o caso do distrito de Bobonaro, e à volta de 190 para Covalima, num total de mais de 5000 observações individuais (Quadro 3).
30
Quadro 3 - Sumário da distribuição das espécies por número de observações no
terreno.
Distrito de Bobonaro Distrito de Covalima
Observações Espécies Observações Espécies
1 a 10 105 1 a 10 124 11 a 20 16 11 a 20 29 21 a 30 14 21 a 30 9 31 a 40 2 31 a 40 6 41 a 50 1 41 a 50 2 51 a 60 1 51 a 60 3 61 a 70 3 61 a 70 2 71 a 80 1 71 a 80 2 81 a 90 1 81 a 90 1 91 a 100 2 91 a 100 2 >100 4 >100 6 Total 150 Total 186
A distribuição das espécies pelas unidades de amostragem e pelo território não é semelhante, o que torna ainda mais complicado o tratamento de dados e a criação de equações de volume. Algumas destas espécies surgem com um número de observações insuficiente para se obter uma base de dados cabal e que sustente o desenvolvimento de uma equação individual, daí haver a necessidade de se proceder ao agrupamento. É também importante que as espécies inventariadas tenham valores de diâmetro bem distribuídos pelas diferentes classes, de modo a que se possa ter informação para todas as suas fases de desenvolvimento e que a equação seja a mais adequada possível (Quadros 4, 5 e 6). Note-se que as espécies se encontram representadas nos Quadros seguintes por um código. Cada um corresponde a uma espécie, conforme consta no anexo deste trabalho.
No âmbito deste trabalho foram ensaiados os modelos não lineares indicados na secção 2.2 (Eq. 2 e 3) usando como regressores as variáveis diâmetro a 1.30m, com casca (d, cm), e altura até aos primeiros ramos vivos (hbc, m). Estes modelos têm mostrado grande flexibilidade no ajustamento a dados de espécies florestais diversas, dada a alometria que lhes está subjacente, razão pela qual foram os modelos selecionados para suporte a este estudo.
31
Quadro 4 - Resumo da distribuição das observações por classe de diâmetro para as espécies do distrito de Bobonaro (≥ 30 observações).
Esp. Obs. 10 a 15 15-20 20-25 25-30 30-35 35-40 40-45 45-50 50-55 55-60 60-65 65-70 70-75 75-80 80-85 85-90 90-95 95-100 >100 101 59 9 7 11 10 3 4 5 3 3 1 - 1 - - 1 - - - - 106 67 8 7 11 4 4 8 5 6 3 - 3 5 2 1 - - - - - 108 73 16 17 16 9 9 5 - - 1 - - - - 111 41 8 7 5 3 6 2 1 4 2 1 2 - - - - 1 - - - 140 38 14 4 7 9 2 2 - - - - 145 91 8 9 10 13 9 9 3 9 3 8 6 2 1 1 - - - - - 169 230 34 37 39 30 30 19 17 8 3 7 2 - - - 2 2 - - - 223 38 5 3 12 3 5 2 3 1 2 1 - 1 - - - - 239 124 17 13 27 19 20 11 4 4 4 2 - 1 - 1 1 - - - - 247 196 33 28 32 28 18 15 13 8 7 7 4 1 2 - - - - 252 137 12 17 25 26 15 20 9 5 4 1 1 - 1 - 1 - - - - 258 66 12 7 16 10 9 3 2 3 1 2 - - - 1 267 91 31 22 19 6 6 5 0 1 1 - - - - 308 67 18 15 14 13 3 2 1 - - - 1 - - Totais 1318 225 193 244 183 139 107 63 52 34 30 18 11 6 3 5 3 1 0 1
33
Quadro 5 - Resumo da distribuição das observações por classe de diâmetro para as espécies do distrito de Covalima (≥ 30 observações).
Esp. Obs. 10 a 15 15-20 20-25 25-30 30-35 35-40 40-45 45-50 50-55 55-60 60-65 65-70 70-75 75-80 80-85 85-90 90-95 95-100 >100 101 79 11 13 21 10 9 9 2 1 - 3 - - - - 107 62 7 8 7 11 2 6 3 3 5 2 3 2 3 - - - - 109 32 7 5 14 3 2 1 - - - - 110 96 18 18 21 18 10 6 3 2 - - - - 111 96 8 9 12 14 9 12 10 9 2 1 4 2 1 1 2 - - - - 112 40 4 2 8 2 3 3 - 1 3 2 3 3 - 1 3 - - - 2 127 57 9 8 9 14 8 6 1 2 - - - - 145 33 8 8 10 5 - 1 - 1 - - - - 149 152 8 9 13 19 18 21 15 10 12 8 6 4 3 1 2 2 1 - - 150 53 6 4 8 12 5 6 4 5 - 2 - 1 - - - - 151 51 13 8 14 10 4 2 - - - - 162 30 13 6 8 1 1 - 1 - - - - 185 125 13 10 20 14 18 12 15 9 5 5 1 1 - - - - 1 - -
35
Quadro 6 - Resumo da distribuição das observações por classe de diâmetro para as espécies do distrito de Covalima (≥ 30 observações).
Continuação. Esp. Obs. 10 a 15 15-20 20-25 25-30 30-35 35-40 40-45 45-50 50-55 55-60 60-65 65-70 70-75 75-80 80-85 85-90 90-95 95-100 >100 216 33 13 5 6 4 2 - 1 - 1 1 - - - - 234 43 18 10 9 2 4 - - - - 238 39 20 13 3 1 2 - - - - 277 142 20 18 29 24 14 10 8 5 5 3 2 1 2 - 1 - - - - 285 182 29 19 29 29 23 16 14 9 6 4 - 1 1 1 - 1 - - - 286 42 9 10 15 3 4 1 - - - - 294 113 14 7 19 15 22 17 6 4 6 1 1 1 - - - - 295 70 18 16 20 9 5 2 - - - - 301 102 20 13 24 20 8 7 6 1 2 - 1 - - - - 314 42 26 14 2 - - - - 318 30 3 6 7 8 3 3 - - - - 327 30 10 9 7 3 - 1 - - - - Totais 1774 325 248 335 251 176 142 89 62 47 26 14 16 10 4 8 3 2 0 2
36
3.2.
Tratamento da base de dados
A recolha dos dados no campo dá-se, em geral, através da utilização de fichas de campo que enumeram diversos parâmetros dendrométricos a serem avaliados. Posteriormente, estes dados são passados para uma base de dados informática ordenada que facilita o trabalho com a informação. Foi a partir deste suporte informático que este trabalho foi desenvolvido e fundamentado. Como tal, e em primeira instância, o trabalho teve início numa análise e consequente reordenação dos dados dendrométricos dos exemplares medidos por forma a permitir uma maior facilidade no estudo de certos parâmetros e relações, tais como a relação do desenvolvimento da mesma espécie em diferentes locais de estudo, o comportamento da espécie em relação a outras, etc. Serviu também para detetar valores pouco verosímeis ou incorrectos e que possam ter sido mal escritos.
A avaliação do comportamento em diferentes distritos para a mesma espécie é um dos pontos mais importantes desta investigação, pois uma espécie que apresente caraterísticas fenotípicas semelhantes tanto em Bobonaro como em Covalima poderá vir a ter associada uma única equação de volume, diminuindo a logística de inventários futuros. Foi então decidido que este seria o primeiro passo na elaboração do trabalho.
Criou-se então uma base de dados por espécie que se apresentasse como comum a ambos os distritos e preenchesse o requisito mínimo de 30 observações para cada local de estudo (secção 3.2). Dentro desta compilação informativa estavam presentes os dados colhidos para as espécies de Bobonaro e para Covalima. Para testar a influência do local usou-se uma variável binária (secção 2.2) associada ao modelo base da equação volumétrica. A análise do efeito do local foi feita com o auxílio das capacidades de estimação de modelos do programa Eviews (versão 5.1) que permite testar, em simultâneo, a significância dos coeficientes associados aos termos que contêm possíveis diferenças de comportamento associadas à origem dos dados.
A reordenação da base de dados não se limitou à seleção das espécies que ambos os distritos partilhavam. Após serem filtradas as espécies comuns, permaneceram as que eram específicas de cada distrito. Ora, estas foram então separadas em duas categorias diferentes, as que correspondiam às espécies com pelo menos trinta observações (e que sustentariam um equação individual) e as espécies cujas observações não perfaziam este
37 valor mínimo para poderem fundamentar uma equação. As últimas foram referenciadas num mesmo lote para serem associadas em grupos, no momento da agregação.
3.3.
Agrupamento de dados de ambos os distritos
A simplificação dos processos de cálculo e a redução dos custos e da logística de gabinete e de campo são dois aspetos tidos em conta na elaboração deste trabalho. Considerando este facto, abordou-se o método de aplicação de uma variável binária (secção 2.2) para a determinação dos comportamentos de crescimento para as espécies inventariadas nos dois distritos timorenses estudados. Caso o comportamento se mostrasse idêntico em ambos os distritos os dados poderiam ser tratados em simultâneo e sem discriminação de proveniência (Figura 5).
Como requisito mínimo para poderem suportar uma equação individual, impôs-se um número mínimo de 30 obimpôs-servações para cada espécie (impôs-secção 3.3.1). No entanto, existem espécies que não completavam esse valor mínimo num dos distritos ou em nenhum. Para casos como este adotou-se outra abordagem (secções 3.3.2 e 3.3.3).
3.3.1 Espécies com pelo menos 30 observações em cada distrito
Para espécies que preenchiam este número de observações e que haviam passado com sucesso nos testes com a variável binária, que estudava o comportamento nos dois locais diferentes de estudo, não foi feita qualquer modificação. Os dados foram agrupados e desenvolveu-se uma única equação de volume para a espécie para poder ser usada tanto para Bobonaro e Covalima (Figura 5).
3.3.2 Espécies com pelo menos 60 observações em ambos os distritos
Na maioria dos casos, as espécies não possuíam o valor mínimo das 30 observações num dos distritos de maneira a que pudessem suportar uma equação individual. No entanto, em alguns casos, as espécies possuíam um número superior a 30 observações num dos dois distritos mas não perfaziam o número mínimo no outro. Estabeleceu-se, então, que para estes casos as espécies que perfizessem 60 observações no total das observações de Bobonaro e Covalima podiam ser consideradas aptas para38 suportar e desenvolver uma equação individual. Isto, nos casos em que o ensaio quanto ao agrupamento (assente na aplicação de variáveis binárias e na análise gráfica) não evidenciou uma tendência em contrário (Figura 5).
3.3.3 Restantes casos
Após a aplicação das duas opções expostas nas secções 3.3.1 e 3.3.2, houve espécies que não preenchiamm qualquer dos requisitos. Sendo assim, e como seriam incapazes de suportar uma equação que fosse representativa, foram remetidas para o processo de agrupamento, onde foram distribuídas por grupos homogéneos para os quais se desenvolveram equações gerais e com potencialidade de serem aplicadas a um rol de espécies florestais para aquele tipo de floresta e localização (Figura 5).
39
3.4.
Aplicação de uma variável binária para estudo da
influência do local
Para averiguar a influência do local no desenvolvimento das espécies usou-se um conjunto de quatro modelos às quais foi adicionada uma variável binária referente à localização.
log v = β0 + β1×log (d2hbc) (18)
log v = β0 + β1× L + β2×log (d2hbc) (19)
log v = β0 + β1×log (d2hbc) + β2×log (d2hbc) × L (20)
log v = β0 + β1× L + β2×log (d2hbc) + β3×log (d2hbc) × L (21)
Sendo:
v = Volume individual de cada árvore (m3)
d = Valor de diâmetro sobre casca de cada árvore, a 1,30 metros de altura (cm) hbc = Altura até aos primeiros ramos vivos de cada árvore (m)
β0 = Constante que indica a ordenada na origem
β1, β2 e β3= Coeficientes parciais da regressão
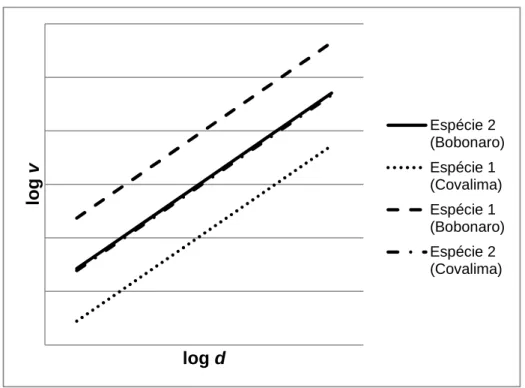
40 O modelo 18 presume, sem a aplicação da variável binária, a hipótese de ambos os locais apresentarem igual padrão comportamental na ordenada na origem e na inclinação da reta (Figura 2). Consideram-se os dados de ambos os distritos como agrupados. Na aplicação do modelo 19, já com a inclusão da variável binária, testa-se a possibilidade de aos dados de Bobonaro e de Covalima corresponderem diferenças na ordenada na origem e, como tal, não poderem ser tratados como idênticos (Figura 2).
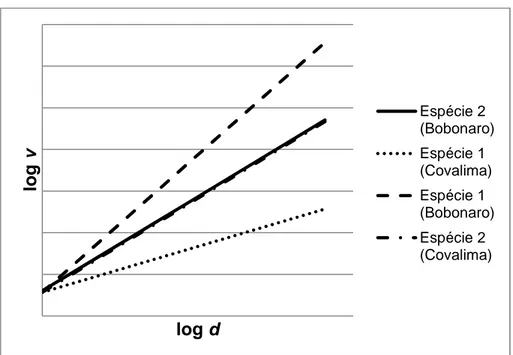
Com a aplicação do modelo 20, presume-se que não existem diferenças quanto à ordenada na origem e avalia-se o comportamento em relação à inclinação da reta. Caso haja diferença no padrão gráfico e no comportamento estatístico os dados não podem ser agrupados num mesmo conjunto (Figura 3). Por último, com a aplicação do modelo número 21, avaliam-se os dois parâmetros em simultâneo. Isto é, na mesma equação inclui-se o efeito da variável binária na ordenada na origem e na inclinação da recta (Figura 4).
Figura 2 - Representação gráfica do padrão esperado para duas espécies mediante a
aplicação das equações de volume relativas aos modelos 18 e 19.
lo g v log d Espécie 2 (Bobonaro) Espécie 1 (Covalima) Espécie 1 (Bobonaro) Espécie 2 (Covalima)
41
Figura 3 - Representação gráfica do padrão esperado para duas espécies mediante a
aplicação da equação de volume relativa ao modelo 20.
Figura 4 - Representação gráfica do padrão esperado para duas espécies mediante a
aplicação da equação de volume relativa ao modelo 21.
lo g v log d Espécie 2 (Bobonaro) Espécie 1 (Covalima) Espécie 1 (Bobonaro) Espécie 2 (Covalima) lo g v log d Espécie 2 (Bobonaro) Espécie 1 (Covalima) Espécie 1 (Bobonaro) Espécie 2 (Covalima)
42
3.5.
Estimação de modelos para as espécies menos
representadas (agrupamento por “clusters”)
Uma das problemáticas da investigação é a estimação de modelos acurados para as espécies que não apresentem o valor mínimo de trinta observações para determinado distrito. Este facto condiciona a construção de uma equação volumétrica individual suficientemente representativa para a espécie. Como solução a este problema, e visto que é situação comum a várias dezenas das espécies inventariadas, surge o agrupamento em grupos homogéneos de espécies (“clusters”) e a posterior criação de equações para os grupos (secção 2.3).
A análise por “clusters” está dependente de um leque de variáveis e dos seus valores para cada uma das espécies que se pretenda agrupar. Por exemplo, num estudo realizado na floresta amazónica usaram-se as caraterísticas do solo para distinguir as espécies inventariadas (Lima et al., 2003).
Não existe uma regra que estipule as variáveis ideais para determinado estudo mas, para criação de modelos de crescimento ou produção, estas devem traduzir processos como o aumento de diâmetro, vigor da regeneração, mortalidade, etc. (Phillips e van Gardingen, 1999). Segundo Zhao et al. (2004), variáveis como a idade, a altura máxima e o índice de qualidade de estação possuem pouca importância em povoamentos como os em estudo nesta dissertação, referindo como mais adequados os parâmetros relacionados com a dimensão.
Tendo isto em conta e a base de dados disponível, considerou-se que a melhor solução seria a aplicação de um método separativo através da relação de diâmetros e da forma do fuste. Para isso utilizou-se um conjunto de dois quocientes, o v/vcilindro e o di/d. Ou seja, o primeiro quociente divide as espécies comparando ao volume do cilindro imaginário, que vai desde o cepo aos primeiros ramos vivos e com diâmetro igual ao diâmetro medido a 1,30 metros, o volume médio da espécie quantificado com o relascópio. O segundo quociente separa as espécies com base no quociente entre o diâmetro a um nível superior (di) medido à altura dos primeiros ramos vivos pelo diâmetro a 1,30 metros.
Em ambos, um resultado inferior a 1 demonstra que o tronco possui a tradicional forma cónica. Para valores iguais a 1 temos um tronco de forma cilíndrica e para valores superiores a 1 temos os casos em que a espécie apresenta um tronco cónico invertido,
43 em que a parte superior e mais larga que a base. Quocientes deste género podem corresponder a árvores que apresentam um alargamento no fuste junto à base da copa, provocado pelo desenvolvimento dos ramos.
Optou-se por utilizar dois quocientes, um baseado na média e outro na mediana dos valores de diâmetro e volume, por forma a evitar os casos em que existam valores demasiado díspares e que a média se pudesse ver afectada, e com isso adulterar a separação das espécies. Para isto, a informação de campo foi organizada por espécies e calculou-se a média e mediana dos diâmetros e volumes para a espécie com base nas medições individuais. O valor dos quocientes foi mais tarde obtido através da divisão das médias e medianas calculadas (Quadros 16, 17 e 18).
Após o cálculo de cada um destes quocientes e das médias de cada um para cada uma das espécies alvo do processo de agrupamento recorreu-se às capacidades do JMP 7.0 para as juntar por grau de similitude. Este programa estatístico disponibiliza dentro de um dos seus menus a opção de análise multivariante e dentro desta encontra-se a opção “Cluster”. Depois de selecionar esta opção toca ao operador eleger que variáveis pretende utilizar para separar as espécies, bem como o método pelo qual quer que seja feita a separação. No caso deste trabalho já havia sido referido que seriam utilizados os quocientes v/vcilindro e o d/d (1,30m) e como método de separação elegeu-se o método de Ward. Neste método os diferentes elementos são juntos através da análise ANOVA da soma de quadrados das variáveis utilizadas na avaliação. O programa junta-os pelo valor mínimo de variância entre eles. E, após ter encontrado um par que seja compatível, funde-os e vai em busca de um outro elemento que esteja próximo destes que volta a fundir e volta a procurar outro elemento próximo. Este processo termina quando cada elemento tem um grupo e quando não há mais modificações possíveis. Ao fazê-lo assim aumenta a fiabilidade dos grupos pois torna as junções mais precisas e aumenta a sensibilidade a “outliers”.
44 A fórmula de distância para este método é:
=| − |
+
Onde:
DKL = Distância entre cluster K e L = Vector médio para o cluster K = Vector médio para o cluster L
= Número de observações para K = Número de observações para L
45
Figura 5 - Diagrama do processo de separação das espécies de acordo com as características de proveniência e número de observações.
Espécies de Bobonaro Espécies de Covalima
Espécies exclusivas de Covalima
Espécies exclusivas de Bobonaro Espécies comuns a ambos os distritos
≥ 60 Observações ≤ 60 Observações ≥ 30 Observações ≤ 30 Observações ≤30 Observações ≥ 30 Observações Equação Individual S. 3.2.1 Equação Individual S. 3.2.1 Não conjugáveis Possível de conjugar Equação de grupo S. 3.2.3 Eq. Suportada para
ambos os locais S. 3.2 a 3.2.2 ≤30 Observações ≥ 30 Observações ≤30 Observações ≥30 Observações
46
3.6.
Análise da qualidade de estimação e obtenção das estimativas
de volume
Todos os cálculos que foram falados nas secções anteriores são estimativas ou previsões baseadas em resultados reais das medições que foram feitas durante o 1º Inventário Florestal de Timor Leste. Ao serem considerados como estimativas significa que vêm acompanhados de um erro associado ao desvio natural que se observa de indivíduo para indivíduo na natureza. Como tal, há que considerar este desvio aquando da análise ou aplicação dos modelos aqui estimados. De seguida serão apresentadas algumas variáveis ou aplicações matemáticas que nos permitem ajuizar os resultados das nossas estimações com maior rigor.
3.6.1 Análise do coeficiente de determinação
O coeficiente de determinação (R2) será o primeiro dos parâmetros de medição de qualidade do modelo a ter em consideração numa análise sumária. Apresenta-se como um valor numérico entre 0 e 1 e indica a percentagem de valores explicados da variável dependente (no caso deste trabalho será sempre o volume) em relação à variabilidade demonstrada pela (s) variável (s) independente (s), ou seja, as variáveis diâmetro e altura do fuste. Isto é o mesmo que dizer que para um valor de R2 igual a 0,9 o modelo explica 90% da variância para a variável dependente (Hair et al., 1998; Rawlings et al., 1998).
Tal como foi feito para este trabalho, muitos autores aconselham a utilização não do coeficiente de determinação mas sim do coeficiente de determinação ajustado (R2 ajustado). Este é uma versão modificada do anterior e considera o número de variáveis explicativas e o tamanho da amostra considerada. Fazendo, em geral, um melhor ajuste do modelo caso as variáveis independentes utilizadas tenham grande poder de explicação, passando exactamente o contrário caso não possuam forte poder de explicação (Hair et al., 1998).
Como valor mínimo para considerar qualquer um dos modelos adequados para o cálculo do volume para qualquer uma das espécies optou-se por um valor de R2 ajustado igual a 0,7. Ou seja, um modelo deverá explicar pelo menos 70% da variância
47 da variável dependente para ser considerado um modelo apropriado e susceptível de poder vir a ser analisado com mais detalhe. Este valor torna-se realmente importante para as espécies que serão agrupadas em grupos e onde os valores de variância serão maiores e terão mais influência na construção de um bom ajuste.
3.6.2 Análise do comportamento dos resíduos
A utilização de dados amostrais colhidos no terreno para fazer estimativas de comportamento ou produção para um qualquer momento do ciclo de crescimento de um exemplar florestal está associada a um erro ou resíduo (ε). É muito difícil fazer estimativas em que o valor deste erro seja zero. O que acontece é considerar-se que o erro da amostra existe mas que é aleatório (Hair et al., 1998).
Uma fase muito importante na análise de modelos é exactamente a avaliação do comportamento gráfico dos resíduos em busca de comportamentos não desejados ou anómalos, como são por exemplo a heterocedasticidade e a especificação incorreta.
A heterocedasticidade é um fenómeno que traduz um comportamento flutuante ou de crescente variância dos resíduos para o intervalo de valores analisado. Sendo portanto um comportamento que não deve ser considerado válido (Fonseca et al., 1998).
A logaritmização dos modelos é um processo proposto para lidar com este fenómeno, mas nem sempre o resolve, sendo por isso importante complementar a avaliação com a análise gráfica do comportamento dos resíduos em relação aos valores estimados.
48
3.6.3 Retransformação do desvio padrão dos resíduos para as unidades
originais e percentagem e aplicação de um fator de correção aos
modelos logaritmizados.
Por forma a facilitar a análise dos modelos e a perceção da sua qualidade as tabelas de resultados vêm acompanhadas por um conjunto de variáveis, algumas abordadas dentro desta secção, que dão uma imagem mais esclarecedora do comportamento do modelo. Uma dessas variáveis diz respeito ao valor dos resíduos ou desvios da estimativa realizada.
Para que a ordem de grandeza dos erros seja mais facilmente visualizada existe a possibilidade de converter as unidades em que surgem para outras mais tangíveis e mais comuns para o operador como são a percentagem e a unidade de medida original, neste caso, o metro cúbico. Para isto usaram-se as fórmulas:
!"# (m³) = &12*(. 4º (.6,á(),*∑(( ( )*+,-./0)
!"# (%) = 9!v ; × 100"#
O presente relatório tem muito do seu fundamento nas aplicações dos modelos logaritmizados como forma de contrariar os elevados padrões de variância heterogénea observados na natureza. O que a logaritmização faz é homogenizar esta enorme variância ao longo da amostra analisada. Sendo este um processo bastante comum é importante saber que ao homogenizar-se a amostra está a introduzir-se um erro nas estimativas feitas e, como tal, este erro por mais pequeno que seja deve ser contrariado (Sprugel, 1983).
De maneira a eliminar o desvio introduzido pela logaritmização o valor final do cálculo deve ser multiplicado por um factor de correção, que é calculado a partir do desvio padrão dos resíduos da regressão original (ou seja, do modelo logaritmizado). Este, por sua vez, é calculado através da fórmula:
49
!"#= ?∑(log C − F,− log CE )D
Em que:
Log yi = Valores para a variável Y
GHI CJ E =Valores para a variável Y obtidos com a equação
n = Número de observações
p = Número de parâmetros do modelo.
Após o valor do desvio padrão da regressão ter sido calculado, pode estimar-se o factor de conversão (Correction Factor, CF) a aplicar:
51
4.
Resultados
4.1.
Avaliação da influência do local para as espécies comuns
No estudo do efeito do local usou-se o teste de Wald para averiguar a veracidade da hipótese nula, relativa à equação 21 da secção 3.2 (log v = β0 + β1 × L + β2 × ln
(d2hbc) + β3 × log (d2hbc) × L) formulada da seguinte forma:
H0: β1 = β3=0
Ha: Pelo menos uma das restrições em H0 é falsa.
Considerando os resultados da análise do efeito do local, expostos no Quadro 7, verificou-se que a hipótese nula podia ser aceite para um grupo dessas espécies. Para as espécies que apresentavam valores que dessem garantia que a influência do local no seu desenvolvimento era nula ou insignificante, procedeu-se à aplicação de um modelo que considerava os valores colhidos sem qualquer distinção, tomando-os como provenientes do mesmo local. Ou seja, das dezanove espécies consideradas para este estudo, apenas cinco não poderão ser tomadas como iguais entre os dois distritos. São elas, Cassia
fistula, Casuarina junghuhniana, Eucalyptus alba, Schleichera oleosa e Sesbania grandiflora.
No ajustamento dos modelos usaram-se estimadores robustos que atendessem a uma eventual situação de variância não homogénea ou de autocorrelação do termo do erro, caraterísticas que poderiam colocar em causa a inferência estatística realizada.
53
Quadro 7 - Resultados do efeito da proveniência com recurso à aplicação da variável binária para as espécies comuns a ambos os distritos.
Teste de Wald Espécie Obs. B/C* R² (aj.) T₀ T₁ T₂ T₃ XY₀ XY XY XY P ( T₁) P ( T₃) P (F) P (χ²) Acacia leucophloea 138 59/79 0,989 -4,076 0,025 0,975 -0,004 0,040 0,058 0,012 0,018 0,686 0,845 0,217 0,214 Albizia chinensis 129 67/62 0,647 -1,807 -0,067 1,338 -0,002 0,119 0,160 0,109 0,145 0,675 0,988 0,447 0,445 Albizia lebbeck 105 61/44 0,751 -1,982 -0,230 1,200 0,252 0,076 0,162 0,087 0,165 0,159 0,130 0,310 0,306 Albizia procera 114 18/96 0,985 -4,013 -0,012 0,955 0,001 0,051 0,062 0,015 0,018 0,848 0,997 0,486 0,484 Aleurites moluccana 124 28/96 0,981 -4,193 0,150 1,004 -0,042 0,145 0,151 0,043 0,044 0,324 0,340 0,580 0,579 Alstonia scholaris 83 43/40 0,741 -2,038 0,248 1,499 -0,179 0,083 0,275 0,080 -0,179 0,369 0,423 0,646 0,645 Cassia fistula 71 38/33 0,646 -2,061 0,379 1,398 -0,522 0,068 0,163 0,087 0,202 0,023 0,012 0,037 0,032 Casuarina equisetifolia 175 23/152 0,989 -3,930 -0,105 0,930 0,024 0,062 0,071 0,017 0,019 0,145 0,206 0,288 0,286 Casuarina junghuhniana 144 91/53 0,704 -1,851 -0,139 1,353 -0,004 0,097 0,150 0,086 0,127 0,358 0,975 0,005 0,004 Eucalyptus alba 355 230/125 0,729 -1,977 0,183 1,439 -0,143 0,042 0,077 0,049 0,085 0,019 0,091 0,027 0,026 Leucaena leucocephala 71 28/43 0,985 -4,083 0,005 0,976 -0,003 0,082 0,094 0,025 0,029 0,958 0,921 0,923 0,923 Peltophorum pterocarpum 60 39/21 0,989 -3,968 -0,011 0,943 -0,001 0,041 0,084 0,012 0,026 0,892 0,983 0,641 0,639 Pterocarpus indicus 266 124/142 0,668 -1,929 0,080 1,319 -0,111 0,069 0,121 0,065 0,110 0,511 0,314 0,304 0,303 Schleichera oleosa 378 84/182 0,663 -1,598 -0,220 1,192 0,150 0,063 0,083 0,085 0,115 0,009 0,193 0,000 0,000 Sesbania grandiflora 64 22/42 0,982 -3,850 -0,475 0,888 0,149 0,070 0,109 0,022 0,032 0,000 0,000 0,000 0,000 Tamarindus indica 251 137/114 0,654 -1,796 0,100 1,294 -0,159 0,057 0,010 0,066 0,109 0,339 0,146 0,128 0,126 Tectona grandis 92 22/70 0,977 -3,920 -0,066 0,909 0,029 0,137 0,142 0,043 0,045 0,644 0,520 0,370 0,366 Timonius timon 168 66/102 0,613 -1,970 0,207 1,377 -0,306 0,107 0,136 0,133 0,164 0,129 0,064 0,132 0,129 Ziziphus mauritiana 153 91/63 0,980 -4,053 -0,038 0,979 0,010 0,037 0,046 0,013 0,016 0,409 0,532 0,374 0,372
54
4.2.
Aplicação de modelos volumétricos
Após identificar que espécies podiam ser tratadas como iguais em ambos os distritos e considerando o número de observações para as restantes dentro de cada distrito, organizou-se a base de dados para permitir cumprir o segundo objectivo deste relatório, a aplicação de modelos matemáticos que permitissem obter indicação do volume individual para cada espécie da maneira mais rápida e precisa possível.
Numa primeira fase, os modelos matemáticos 2 e 3 foram aplicados usando os dados das espécies que na secção 4.1 haviam sido classificadas como conjugáveis. Com os resultados desta análise construíram-se os Quadros 8 e 9. Posteriormente compilaram-se para cada distrito as espécies com pelo menos trinta observações e aplicaram-se os dois modelos propostos na secção 2.1 (Quadros 10 a 13).
Durante o inventário da floresta timorense houve problemas em identificar algumas das espécies sendo nesses casos atribuída a designação de “desconhecida” ou era registada, na ficha de campo, com o nome local. Para esses casos foi também estabelecida uma equação de volume baseada nos dois modelos usados nos casos anteriores com o objetivo de permitir futuros casos de avaliação volumétrica ou para estudos de biomassa. Nesta situação, juntaram-se todas as espécies sem identificação num mesmo conjunto, independentemente do distrito de origem (Quadros 14 e 15).
55
Quadro 8 - Resultados da aplicação de um modelo de regrassão linear simples (Eq. 2) sem diferenciação da proveniência dos dados para as
espécies comuns aos dois distritos e que mostraram não haver influência do local no seu comportamento.
Aplicação do modelo 2 log v = β₀ + β₁ × log (d²hbc) para as espécies comuns
Espécie Obs. R² (aj.) T₀ T₁ XY₀ XY Syx Syx (m3) Syx (%)
Acacia leucophloea 138 0,989 -4,056 0,971 0,029 0,009 0,047 0,053 20,368 Albizia chinensis 129 0,649 -1,823 1,320 0,085 0,086 0,323 0,407 77,463 Albizia lebbeck 105 0,750 -2,054 1,293 0,068 0,073 0,207 0,138 70,909 Albizia procera 114 0,985 -4,022 0,954 0,038 0,011 0,052 0,035 14,746 Aleurites moluccana 124 0,982 -4,058 0,965 0,043 0,012 0,063 0,085 19,284 Alstonia scholaris 83 0,742 -1,974 1,457 0,099 0,095 0,328 0,488 82,773 Casuarina equisetifolia 175 0,988 -4,002 0,947 0,031 0,008 0,060 0,318 24,525 Leucaena leucocephala 71 0,986 -4,083 0,975 0,043 0,014 0,044 0,013 11,319 Peltophorum pterocarpum 60 0,990 -3,977 0,945 0,043 0,013 0,052 0,036 11,116 Pterocarpus indicus 266 0,669 -1,891 1,261 0,056 0,054 0,273 0,301 79,811 Tamarindus indica 250 0,653 -1,748 1,215 0,048 0,056 0,025 0,160 62,157 Tectona grandis 92 0,976 -3,979 0,934 0,049 0,015 0,062 0,028 18,216 Timonius timon 168 0,605 -1,850 1,200 0,065 0,075 0,271 0,406 16,976 Ziziphus mauritiana 153 0,979 -4,071 0,984 0,033 0,012 0,052 0,008 11,352
57
Quadro 9 - Resultados da aplicação de um modelo de regressão múltipla (Eq. 3) sem diferenciação da proveniência dos dados para as espécies
comuns aos dois distritos e que mostraram não haver influência do local no seu comportamento.
Aplicação do modelo 3 log v = β₀ + β₁ × log d + β2 × log (hbc) para as espécies comuns
Espécie Obs. R² (aj.) T₀ T₁ T₂ XY₀ XY XY Syx Syx (m3) Syx (%)
Acacia leucophloea 138 0,993 -4,112 2,021 0,871 0,023 0,016 0,013 0,037 0,038 14,438 Albizia chinensis 129 0,994 -4,123 2,034 0,849 0,024 0,016 0,012 0,041 0,083 15,765 Albizia lebbeck 105 0,966 -4,098 2,017 0,847 0,063 0,049 0,032 0,077 0,06 31,052 Albizia procera 114 0,991 -4,117 2,040 0,836 0,031 0,023 0,016 0,040 0,027 11,365 Aleurites moluccana 124 0,988 -4,147 2,037 0,862 0,037 0,023 0,016 0,051 0,074 16,829 Alstonia scholaris 83 0,995 -4,117 2,019 0,850 0,029 0,020 0,016 0,045 0,170 19,881 Casuarina equisetifolia 175 0,992 -4,105 2,021 0,846 0,030 0,021 0,014 0,051 0,330 25,444 Leucaena leucocephala 71 0,989 -4,134 2,035 0,888 0,039 0,030 0,022 0,038 0,012 10,524 Peltophorum pterocarpum 60 0,994 -4,066 1,991 0,853 0,036 0,025 0,017 0,039 0,035 10,766 Pterocarpus indicus 266 0,981 -4,088 2,009 0,866 0,030 0,020 0,014 0,065 0,061 16,154 Tamarindus indica 250 0,991 -4,103 2,010 0,890 0,021 0,014 0,009 0,040 0,039 15,227 Tectona grandis 92 0,981 -4,092 2,003 0,836 0,049 0,038 0,024 0,055 0,031 20,167 Timonius timon 168 0,984 -4,097 2,009 0,852 0,033 0,022 0,016 0,055 0,063 26,186 Ziziphus mauritiana 153 0,984 -4,119 2,034 0,880 0,030 0,023 0,019 0,046 0,008 10,473
![Referências técnicas para atuação de psicólogas(os) em Programas de Atenção à Mulher em situação de Violência [2013] - CREPOP CREPOP](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)