Efeito do laser diodo sobre as características de motilidade, de integridade das...
Texto
Imagem



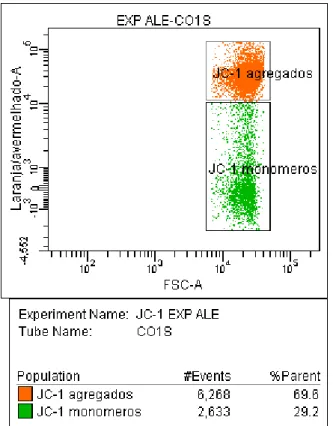




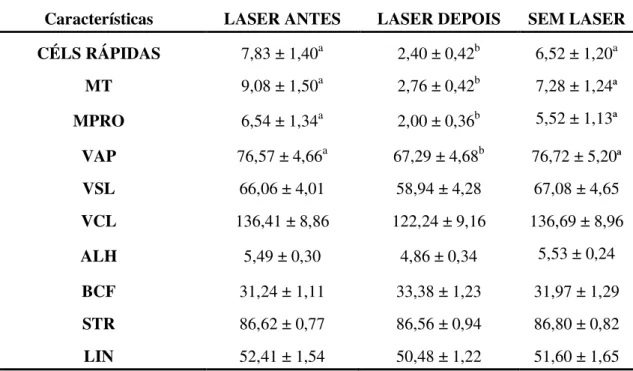
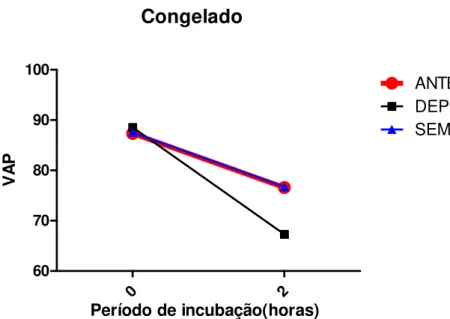
Documentos relacionados
A partir da junção da proposta teórica de Frank Esser (ESSER apud ZIPSER, 2002) e Christiane Nord (1991), passamos, então, a considerar o texto jornalístico como
Isto demonstra mais uma vez o quanto é importante à educação inclusiva para o favorecimento da aprendizagem de ambas as partes, tanto crianças ditas “normais” quanto crianças
In the in vitro study the propolis extract and dentifrice showed efficacy against S.mutans up to the concentration of 0.3906 μg/mL and the Parodontax dentifrice
Seminário Complexidade, Coerência de Políticas e Agenda Global 2030 para o Desenvolvimento Sustentável?. Núcleo de Estudos em EcoPolíticas e EConsCiencias Instituto de
De forma a sustentar esta ideia, recorro a Costa et.al (1996, p.9) quando afirmam que “A aprendizagem da profissão docente não principia com a frequência de um
Nesse mesmo período, foi feito um pedido (Processo do Conjunto da Avenida Rio Branco, no Rio de Janeiro nº 860-T-72) pelo Instituto dos Arquitetos do Brasil e pelo Clube de
Dessa forma, esta pesquisa visa determinar o comportamento dos metais em uma celula experimental, lisimetro, de residuos solidos urbanos (RSU) da cidade de Campina Grande-PB com
Para ambas as amostras não foi observado fases cristalinas na região soldada, indicando que a temperatura não excedeu a temperatura de cristalização das
