Influence of Quercus ilex trees on herbaceous production and nutrient
concentrations in southern Portugal
Elena Cubera1, 2, Jorge Manuel Nunes3, Manuel Madeira1*, and Luíz Gazarini3
1Instituto Superior de Agronomia, Universidade Técnica de Lisboa. Tapada da Ajuda, 1349-017, Lisboa, Portugal 2Ingeniería Técnica Forestal. Universidad de Extremadura. Avenida Virgen del Puerto 2, 10600 Plasencia, Spain 3Universidade de Évora, Departamento de Biología, 7002–554, Évora, Portugal
Abstract
In an open woodland in Portugal, the nature of interactions between Quercus ilex trees and her-baceous plants was assessed during 2 years by studying how manipulation of incident solar radiation, water and nutrient supply affect the herbaceous biomass and N, K, P, Ca, Mg, and Mn concentrations. Measurements were carried out in three environments consisting of (1) open grassland, (2) beneath the tree canopy, and (3) under artificial shade. Each of these environ-ments was subjected to two regimes of fertilization and two water levels in a factorial design. The fertilizer treatment consisted of application of no fertilizer or a combination of 200 kg calcium ammonium nitrate ha–1(26% N) and 350 kg superphosphate ha–1(8% P), while the water-supply
treatment consisted of either no irrigation or irrigation fortnightly from February 1 to April 30. Grasses showed significantly lower nutrient concentrations than forbs. However, nutrient con-centrations of the whole herbaceous community were within the recommended ranges for cattle nutrition. A negative effect of shade on herbaceous biomass production was observed. The effect of watering on herbaceous biomass was less prominent than the effect of fertilization, irre-spective of the environment, suggesting that Q. ilex does not compete for soil-water resources with herbaceous biomass in this ecosystem. Fertilization increased total biomass by 106%, 49%, and 97% in the open grassland, beneath the tree canopy, and under artificial shade, re-spectively. During the first and second year, fertilization increased herbaceous P concentrations by 24% and 83%, respectively, if compared with concentrations obtained at the unfertilized plots. Higher K and Mg concentrations were observed in herbaceous plants beneath the tree canopy than in the open areas, indicating a positive effect of trees on pasture quality. The positive and negative effects of trees on understory forage are discussed.
Key words: herbaceous biomass / shade / fertilization / irrigation Accepted March 4, 2009
1 Introduction
Trees change environmental conditions in the understory in terms of reduced light, less extreme temperatures, altered water availability, and evapotranspiration, soil nutrient avail-ability, and organic-matter status (Joffre and Rambal, 1988;
Gallardo, 2003; Moreno et al., 2007a). The composition of
grassland species differs under tree canopies compared to adjacent open grassland, and this increases species biodi-versity of Mediterranean silvopastoral systems. The presence of trees can reduce (Marañón and Bartolome, 1994; Scholes and Archer, 1997; González-Hernández et al., 1998) or increase (Holland, 1980; Moreno, 2008) productivity of under-storey vegetation compared to surrounding open areas. The overall effect of canopy cover on understorey plant produc-tion is the result of competitive or facilitatory processes occur-ring between the trees and pasture, both above and below the ground level. The positive response is generally attributed to more favorable physical and chemical soil properties and to more favorable soil and air temperatures under trees (Moreno et al., 2007a). However, the relative importance of each factor is not well-known as it varies with tree age, spe-cies, phenological status, soil quality, and annual rainfall
(McPherson, 1997; Garrett et al., 2004). It is necessary to consider to what degree each resource limits growth in a par-ticular environment and how the species involved are suited to compete for or to facilitate the use of such resource. Previous studies in semiarid areas of western Spain have determined that the quality and production of the grassland vary with temporal and spatial gradients (Pérez-Corona et al., 1994, 1998; Vázquez-de-Aldana et al., 2000), producing a complex environment, which is difficult to manage for optimal livestock production. From energetic and economic points of view, herbaceous quality determines the rational use of these silvopastoral ecosystems. Protein concentration and digest-ibility of grasslands have been emphasized as the main determinants of forage quality (Pérez-Corona et al., 1994). However, much less attention has been paid to mineral ele-ments even though they also influence forage quality and can depress feed intake when levels are low (Provenza, 1995). The mineral composition of botanical constituents of the her-bage community is of paramount significance as it is a reli-able indicator of nutritional value. More information about the
effect of trees on understory production and nutritional quality is necessary for improving and determining the sustainability and profitability of silvopastoral systems.
In Europe, the most extended agroforestry systems are the Iberian “montados” (“dehesas” in Spanish), with about 3 million hectares in Portugal and in southwestern Spain (Eichhorn et al., 2006). These man-made savannah-type ecosystems are characterized by a stratum of well-spaced oak trees (10–60 trees ha–1) associated with an herbaceous
understorey. In these systems, livestock grazing meets 60%–70% of cattle annual energetic needs (Campos et al., 2001). The aim of the present study is to assess the nature of interactions between trees and herbaceous plants in monta-dos by studying how manipulation of incident solar radiation, water and nutrient supply affect the herbaceous biomass and herbaceous nutrient concentrations over 2 years with differ-ent rainfall amounts. The positive and negative effects of trees on understorey forage are discussed.
2 Material and methods
2.1 Study area
The study was conducted in an open oak woodland (mon-tado) at the “Herdade da Mitra” experimental area (38°32′ N, 8°01′ W; 243 m above sea level) located near Évora, some 150 km southeast of Lisbon, Portugal. The climate is Mediter-ranean, with dry and hot summers contrasting with mild and rainy winters. Long-term (1951–1980) mean rainfall is around 665 mm per year, with considerable variation throughout the year (rain mainly falls from October to May), and mean annual air temperature is 15°C, ranging from 8.6°C to 23.1°C in January and August, respectively. Mean annual potential evapotranspiration estimated following the approach of
Thornthwaite (1948) is 794 mm. Soils are mainly Haplic
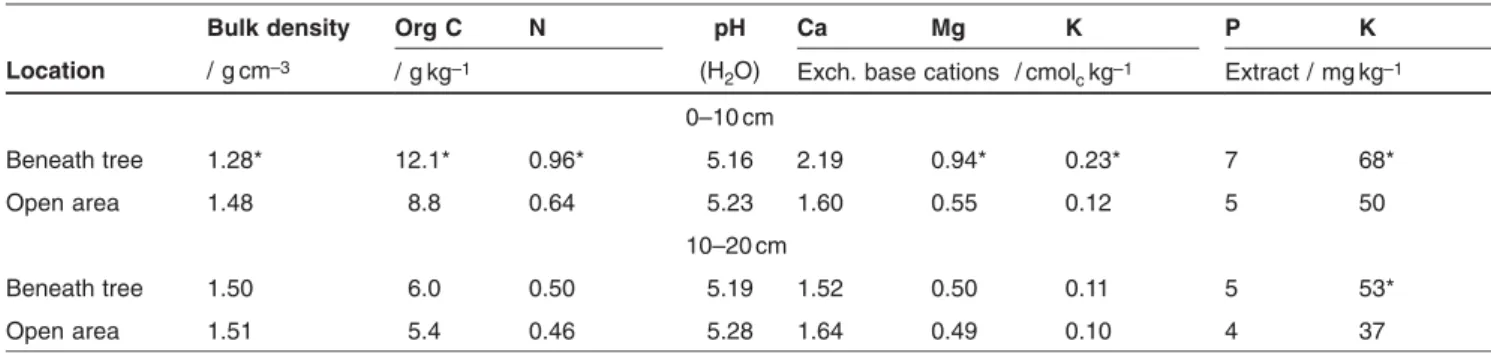
Leptosol (Dystric) derived from gneiss (IUSS Working Group, 2006), with a sandy-loam texture, low organic-matter content and P status, and acidic. The main chemical properties of soils are shown in Tab. 1.
About 0.2 ha of montado (10% slope) covered by natural annual grassland and scattered Quercus ilex trees was selected as the study area. Within the studied area, Quercus
ilex subsp. rotundifolia had an average tree density of 35–45
trees ha–1(61.6% tree-canopy cover). Average tree-canopy
diameter was 14 m, and mean diameter at breast height was 0.59 m. The trees had not been pruned for the previous 6 years. The herbaceous understorey layers comprised annual grasses such as Vulpia bromoides (L.) S.F. Gray, Bromus rigidum Gaudin, Hordeum murinum L., and Briza maxima L., and forbs such as Rumex bucephalophorus L., Silene gallica L.,
Geranium purpureum Vill., Tolpis barbata (L.) Gaertner, Tuberaria guttata L. Fourr. Legumes included Ornithopus compressus L. and Ornithopus pinnatus (Miller) Druce.
2.2 Experimental layout
Herbaceous production and nutrient concentrations were measured around four trees, in twelve 1 m × 1 m plots estab-lished in each of the following three environments: (1) in the open grassland (25 m from the closest tree), (2) beneath the tree canopy (2 m from the trunk), and (3) in the open area under artificial shade. Tree canopy intercepted 70% of the full sunlight, and artificial shade consisted of a plastic mesh (16 threads cm–1) that intercepted 70%–80% of the sunlight.
Plots were protected from grazing by a 1 m high fence. At the end of the summer of 2001, all vegetation inside the enclo-sures was cut at ground level in order to simulate plant con-sumption by grazing herbivores outside the enclosure. In each environment, plots were subjected to fertilization and watering treatments, distributed in a factorial design, with an average distance of 50 cm between them.
Fertilization treatments consisted of (1) a surface application of 200 kg calcium ammonium nitrate ha–1(CAN, 26% N) and
350 kg superphosphate ha–1(8% P) at the beginning of the
growing season (October) (F), as commonly used in the area to regenerate natural pastures (R. Freixial, personal commu-nication), and (2) an unfertilized plot (nF). The watering treat-ments were (1) water applied fortnightly from February 1 to April 30, by applying at each date 10 L m–2by hand and thus
simulating an increase of 70 L m–2of total precipitation from
February to April (W) and (2) a nonwatered plot (nW). Her-baceous biomass was measured during two vegetative peri-ods with different rainfall amounts. From September 2000 to April 2001 and from September 2001 to April 2002, total pre-cipitation was 962 and 641 mm, respectively (Fig. 1). Sam-pling was done in early May, when plants were at the flower-ing-fruiting stage. Herbaceous plants were clipped inside quadrants of 0.25 m2 situated at the center of each plot.
Table 1: The main characteristics of the soils at the experimental site. Asterisks indicate significant differences between the average values
beneath tree canopies and in the open areas (p<5%).
Bulk density Org C N pH Ca Mg K P K
Location / g cm–3 / g kg–1 (H
2O) Exch. base cations / cmolckg–1 Extract / mg kg–1
0–10 cm Beneath tree 1.28* 12.1* 0.96* 5.16 2.19 0.94* 0.23* 7 68* Open area 1.48 8.8 0.64 5.23 1.60 0.55 0.12 5 50 10–20 cm Beneath tree 1.50 6.0 0.50 5.19 1.52 0.50 0.11 5 53* Open area 1.51 5.4 0.46 5.28 1.64 0.49 0.10 4 37
Three replicates per environment and per treatment were considered.
Plants were clipped at ground level and manually sorted into grasses and forbs. Legumes were excluded because their cover values inside the plots were below 8%. Dry matter (DM) was determined by drying samples at 60°C for 48 h in a forced-air oven. The dried plant material was ground with a ball mill prior to analyzing the concentrations of N, P, K, Ca, Mg, and Mn, in grasses and forbs separately. Nitrogen con-centration was analyzed with the Kjeldahl method (Digestion System 40, Kjeltec Auto 1030 Analyzer), P was measured colorimetrically, and K, Ca, Mg, and Mn concentrations were determined using atomic-absorption spectrophotometry.
In November 2000, soil samples were taken around five trees at two distances from the trunk: beneath the tree canopy and in the open area (2 and 20 m from the tree trunk, respective-ly). For each tree and distance, four samples from the 0–10 and 10–20 cm soil layers were combined into one composite sample per soil layer. Samples were sieved (
<
2 mm), air-dried, and analyzed for organic C, total N, exchangeable Ca, Mg, and K, and extractable P and K. Two undisturbed sam-ples selected at both distances per tree were used for the determination of soil bulk density. Organic C was determined after wet oxidation. Total N was analyzed using the Kjeldhal method (Digestion System 40, Kjeltec Auto 1030 Analyzer). Soil pH was determined potentiometrically in distilled water. Exchangeable Ca, Mg, and K were extracted with ammoniumacetate at pH 7 (Chapman, 1965). Extractable P and K were determined using the Egnér-Riehm method (Egnér et al., 1960) and the molybdate-blue method.
2.3 Data analysis
Normality of data was checked with Kolmogorov-Smirnov tests. In order to analyze the effect of trees on herbaceous production and herbaceous nutrient concentrations, several 4-way ANOVAs were undertaken, using total herbaceous bio-mass, grasses biobio-mass, and forbs biomass and herbaceous nutrient concentrations as dependent variables, and year (2001 and 2002), environment (open grassland, beneath canopy, and beneath artificial shade), fertilization treatment (F and nF) and watering treatment (W and nW) as factors. Since 3- and 4-way interactions were not significant, two levels of interaction were considered. Least significant differ-ences (LSD) were used to determine differdiffer-ences between means when significant ANOVA results occurred. Relation-ships between grasses and forbs biomass and total herbac-eous N, P, K, Ca, Mg, and Mn concentrations (weighted aver-age) were analyzed by correlation analyses. For statistical analysis, the program STATGRAPHICS Plus v.4.1 was used.
3 Results
Soils beneath tree canopies showed, in the top 10 cm of the soil profile, higher values of organic C, total N, exchangeable Mg and K, and extractable K than soils in the open areas (Tab. 1). Other soil properties in the 0–10 cm and all proper-ties in the 10–20 cm layer except extractable K were not affected by the tree canopy.
3.1 Herbaceous biomass
Total herbaceous biomass was significantly higher in 2001 than in 2002 (p
<
1%; Tab. 2), with mean values of 283 and 221 g DM m–2, respectively. Differences in biomass betweenyears were only detected for forbs (p = 0.1%) and not for grasses (Figs. 2, 3). The interaction between environment and fertilization (p
<
0.1%; Tab. 2) showed that differences between nF and F were lower beneath tree canopy than in open grassland and under artificial shade. The fertilization treatment in the open area, beneath the tree canopy, and under artificial shade increased total biomass by 106%, 49%, and 97%, respectively. The fertilization treatment increased significantly the biomass of grasses and forbs (p<
0.1% and0 50 100 150 200 250 300 350 Sep Nov Ja n Mar May Ju l Sep Nov Ja n Mar May Ju l Pr ec ip itat io n / mm 0 5 10 15 20 25 T e mper a tur e / ºC Precipitation Temperature 2000 2001 2002
Figure 1: Monthly accumulated precipitation and monthly mean
temperature at the experimental site during the study period. Arrows indicate dates during which herbaceous vegetation was clipped.
2001 0 100 200 300 400 500 600 F nF F nF F nF
Open grassland Beneath canopy Artificial shade
Biom a s s / g D M m –2 Grasses Forbs 2002 0 100 200 300 400 500 600 F nF F nF F nF
Open grassland Beneath canopy Artificial shade
Bioma s s / g D M m –2
a
b
Figure 2: Biomass production of
grasses and forbs measured during 2001 (a) and 2002 (b), under three environments (on the open grassland, beneath the tree canopy, and under artificial shade) and two fertilization treatments (F and nF). Vertical bars represent standard errors of total biomass.
p = 1%, respectively; Fig. 2). The interaction between
envir-onment and fertilization was also observed for the biomass values of grasses (Fig. 2). During both years, unfertilized plots showed higher herbaceous biomass in open areas than beneath the tree canopy or under the artificial shade (254, 139, and 131 g DM m–2, respectively, p
<
0.1%). Fertilizedplots also showed higher herbaceous biomass in open areas than beneath the tree canopy or under the artificial shade (523, 208, and 257 g DM m–2, respectively, p
<
0.1%). Inferti-lized and unfertiferti-lized plots, no differences in biomass produc-tion were observed beneath the tree canopy and under artifi-cial shade.
The effect of watering on total herbaceous biomass was also significant, irrespective of the environment (no interaction environment × watering; Fig. 3). Total herbaceous biomass showed significant differences between W and nW only dur-ing the humid year 2001 (interaction year × waterdur-ing), with mean values of 329 and 238 g DM m–2, respectively. In 2001,
these differences were mainly due to the higher biomass val-ues of forbs under W than under nW (186 and 114 g DM m–2,
respectively).
Considering only the untreated plots (unsheltered, unferti-lized, and not irrigated), total biomass during both years was significantly higher on the open area than beneath the tree canopy (258 and 97 g DM m–2, p
<
0.1%; respectively). Thistrend was observed for both grasses (p = 1%) and forbs (p = 2%). The plot submitted to artificial shade (unfertilized and not irrigated) showed similar total herbaceous-biomass values to the untreated plot beneath the tree canopy.
3.2 Herbaceous nutrient concentrations
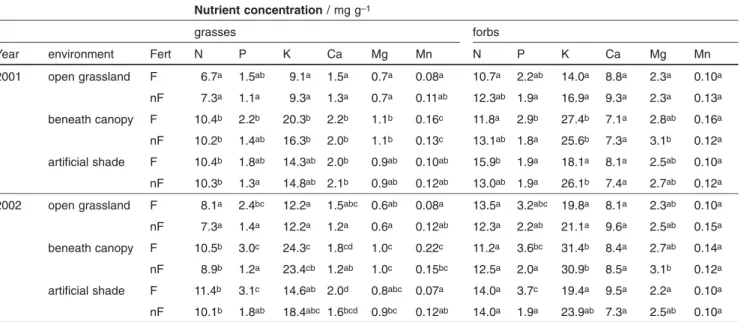
Nutrient concentrations of herbaceous plants did not differ between 2001 and 2002 except for P (Tab. 2), which was 1.9 and 2.3 mg g–1, respectively. Phosphorus concentrations of
grasses and P concentrations of forbs were similar for the three environments studied (Tab. 3). The average N, K, Mg, and Mn concentrations of grasses were higher beneath the tree canopy than on the open area. Grasses under artificial shade showed similar N concentration to grasses under the tree canopy, and intermediate K and Mg values between those observed under the tree canopy and on the open area. Fertilization treatments only affected the P concentration of herbaceous plants (p
<
0.1%, Tab. 2). Differences of herbac-eous P concentrations between F and nF were higher in 2002 than in 2001, as indicated by the interaction between year and fertilization (Tab. 2). In 2001 and 2002, fertilization increased herbaceous P concentration by 24% and 83%, re-spectively. Watering treatments did not affect any of the nutri-ent concnutri-entrations analyzed (Tab. 2). Forbs showed higher N, P, K, Ca, and Mg concentrations than grasses (p<
0.1%), but similar values of Mn (Tab. 3).In untreated plots, higher herbaceous K and Mg concentra-tions were observed beneath the tree canopy than in the open grassland (p = 0.36% and p = 4% for K and Mg, respec-tively, Tab. 3). Phosphorus, Ca, and Mn concentrations were similar, however, in both untreated environments. The corre-lation analysis pointed to a decrease of N, K, Ca, and Mg concentrations in the plant community with increasing bio-mass of grasses (Tab. 4). Calcium concentration was posi-tively correlated with forbs biomass.
4 Discussion
Herbaceous biomass significantly increased with fertilization and irrigation, indicating that herbaceous plants in montados Table 2: Significance levels of multifactorial ANOVAs analyzing the
significance of the herbaceous production and herbaceous nutrient concentrations for 2 years (2001 and 2002), under three environ-ments (on the open grassland, beneath the tree canopy, and under artificial shade), two fertilization treatments (F and nF) and two watering regimens (W and nW). Asterisks indicate levels of significance (***p<0.1%; **p<1%; *p<5%), and ns indicates not significant. 3- and 4-way interactions were not significant.
Factors Biomass N P K Ca Mg Mn Year ** ns ** ns ns ns ns Environment *** *** ns *** ns ** *** Fertilization *** ns *** ns ns ns ns Watering * ns ns ns ns ns ns Year × environment ns ns ns ns ns ns ns Year × fertilization ns ns ** ns ns ns ns Year × watering * ns ns ns ns ns ns Environment × fertilization *** ns ns ns ns ns ns Environment × watering ns ns ns ns ns ns ns Fertilization × watering ns ns ns ns ns ns ns 2001 0 100 200 300 400 500 600 W nW W nW W nW
Open grassland Beneath canopy Artificial shade
Bioma s s / g D M m –2 Grasses Forbs 2002 0 100 200 300 400 500 600 W nW W nW W nW
Open grassland Beneath canopy Artificial shade
Bioma s s / g D M m –2
a
b
Figure 3: Biomass production of
grasses and forbs measured during 2001 (a) and 2002 (b), under three environments (on the open grassland, beneath the tree canopy, and under artificial shade) and two watering regimens (W and nW). Vertical bars represent standard errors of total biomass.
experience deficit of nutrients and soil water. The increase of biomass due to fertilization, albeit lower beneath tree canopy than in the open area and under artificial shade, could be explained by the higher values of organic C, N, exchangeable K, and extractable K observed in soils beneath canopy than in adjacent soils. A positive effect of trees on soil fertility has also been described for grazed “dehesas” (Gallardo, 2003;
Moreno et al., 2007b), North American oak savannas
(Jackson et al., 1990; McPherson, 1997), and other agro-forestry systems (Young, 1997).
At the doses used here, the response of herbaceous biomass to fertilization was more pronounced than to watering. Even in drier “dehesas”, Moreno (2008) reported that pasture pro-duction was more nutrient-limited than water-limited. The similar herbaceous biomass observed beneath the tree canopy and under the artificial shade indicate, however, that grasses and forbs were not strongly competing with trees for soil nutrients. Higher herbaceous biomass in the W plots than in the nW plots were only detected during the humid year 2001. It seems that the effect of watering treatment on her-baceous biomass was more influenced by the precipitation accumulated during April than by the total precipitation accu-mulated during the months prior to clipping. In 2001 and 2002, April precipitation was 13 and 103 mm, respectively (Fig. 1), and W treatments increased these values by 61%
and 16%, respectively. This may explain why additional water applied during 2002 did not produce higher herbaceous bio-mass. Herbaceous plants in montados may depend on the available water accumulated in the first centimeters of the soil, a hypothesis sustained by the fact that shallow herbaceous root systems have been reported for “dehesas” of Q. ilex (Joffre et al., 1987; Moreno et al., 2005). Since watering had a similar effect in the three environments stu-died, it seems that trees and herbaceous plants must have explored different soil layers and do not strongly compete for soil water resources in montados, as stated in previous stud-ies (Moreno et al., 2005; Cubera and Moreno, 2007).
The lower understory biomass registered under tree canopy and artificial shade compared to open areas could be attribu-ted to light reduction. Limiting radiation was a main factor determining density and productivity of herbaceous species in the understorey of similar oak woodlands in California (Parker and Muller, 1982; Marañón and Bartolome, 1993). In our study, interception of 70%–80% of the incident radiation exceeded the value assumed to produce the maximum pas-ture biomass in Mediterranean systems, i.e., around 50% (Étienne, 1996). In the Midwestern U.S., many cool-season forage species benefit from shading equivalent to 40%–60% of light interception (Garrett et al., 2004) and in Spain, a posi-tive effect of 50% light interception on pasture yield was Table 3: Nutrient concentrations (N, P, K, Ca, Mg, and Mn) of grasses and forbs measured during 2001 and 2002, in fertilized (F) and
unfertilized (nF) plots under three environments (open grassland, beneath the tree canopy, and under artificial shade). For each year within columns, different letters indicate significant differences (p<5%).
Nutrient concentration / mg g–1
grasses forbs
Year environment Fert N P K Ca Mg Mn N P K Ca Mg Mn
2001 open grassland F 6.7a 1.5ab 9.1a 1.5a 0.7a 0.08a 10.7a 2.2ab 14.0a 8.8a 2.3a 0.10a nF 7.3a 1.1a 9.3a 1.3a 0.7a 0.11ab 12.3ab 1.9a 16.9a 9.3a 2.3a 0.13a beneath canopy F 10.4b 2.2b 20.3b 2.2b 1.1b 0.16c 11.8a 2.9b 27.4b 7.1a 2.8ab 0.16a nF 10.2b 1.4ab 16.3b 2.0b 1.1b 0.13c 13.1ab 1.8a 25.6b 7.3a 3.1b 0.12a artificial shade F 10.4b 1.8ab 14.3ab 2.0b 0.9ab 0.10ab 15.9b 1.9a 18.1a 8.1a 2.5ab 0.10a nF 10.3b 1.3a 14.8ab 2.1b 0.9ab 0.12ab 13.0ab 1.9a 26.1b 7.4a 2.7ab 0.12a
2002 open grassland F 8.1a 2.4bc 12.2a 1.5abc 0.6ab 0.08a 13.5a 3.2abc 19.8a 8.1a 2.3ab 0.10a
nF 7.3a 1.4a 12.2a 1.2a 0.6a 0.12ab 12.3a 2.2ab 21.1a 9.6a 2.5ab 0.15a
beneath canopy F 10.5b 3.0c 24.3c 1.8cd 1.0c 0.22c 11.2a 3.6bc 31.4b 8.4a 2.7ab 0.14a
nF 8.9b 1.2a 23.4cb 1.2ab 1.0c 0.15bc 12.5a 2.0a 30.9b 8.5a 3.1b 0.12a
artificial shade F 11.4b 3.1c 14.6ab 2.0d 0.8abc 0.07a 14.0a 3.7c 19.4a 9.5a 2.2a 0.10a
nF 10.1b 1.8ab 18.4abc 1.6bcd 0.9bc 0.12ab 14.0a 1.9a 23.9ab 7.3a 2.5ab 0.10a
Table 4: Correlation coefficients between biomass of grasses or forbs and N, P, K, Ca, Mg, or Mn concentrations of the plant community
(grasses + forbs). Asterisks indicate levels of significance (***p<0.1%; **p<1%; *p<5%).
Plant nutrient concentration
Factors N P K Ca Mg Mn
Biomass of grasses –0.61** 0.05 –0.62** –0.53** –0.71** –0.20
recently confirmed (Moreno, 2008). Pruning our trees would reduce light interception and probably would avoid the negative effect of shade on herbaceous production.
Interannual differences in the nutrient concentrations of the plant community were small, suggesting that in semiarid montados, factors other than those related to interannual cli-matic variation are more important in determining the nutrient concentrations of the herbaceous plants. A similar conclusion has been previously drawn for “dehesas” in western Spain (Vázquez-de-Aldana et al., 1996; García-Ciudad et al., 1997). Vázquez-de-Aldana et al. (1996) showed that mineral concentration in plant tissues varies along the topographic slope through differences in soil nutrient status and the tex-ture of the soil. Trees improved the herbaceous quality by increasing their K and Mg concentrations, and this effect may be related to the high exchangeable-K and -Mg values ob-served in the soil (0–10 cm depth) beneath the tree canopy. According to observations of Escudero et al. (1985) for “dehesas”, greater K and Mg concentrations observed in the soil beneath the tree may be associated with additional inputs through litterfall (0.61 kg m–2y–1), throughfall (1.9 and
0.53 g m–2 y–1 for K and Mg, respectively), and stemflow
(0.09 and 0.007 g m–2y–1for K and Mg, respectively).
Varia-tions of herbaceous nutrient concentraVaria-tions depending on the distance from the trunk of Q. ilex have been recently reported (Gea et al., 2007; Moreno et al., 2007b). Herbaceous plants may depend significantly on the nutrients located in the uppermost soil layer, where most of their roots are located. However, in deciduous Q. douglasii savanna similar plant N accumulation was observed beneath and beyond the tree canopy despite the higher N availability of soils under the trees (Jackson et al., 1990), as was also found in our study. Caution must be taken when comparing and interpreting nutrient concentrations of herbaceous plants, especially if plant densities are excluded from the calculations. If the nutri-ent concnutri-entrations of plants were expressed per surface area, herbaceous K and Mg would show similar values in the open areas, beneath the tree canopy, and under artificial shade (3.6, 3.1, and 2.9 g K m–2and 0.37, 0.24, and 0.27 g
Mg m–2, respectively). Therefore, to study the effect of tree ×
fertilization on pasture quality a similar range of plant densi-ties under and beyond the tree canopy should be considered. Since pasture production and nutrient deficiencies limit feed intake (Christian, 1987; Provenza, 1995), both biomass and nutrient concentrations of herbaceous plants should be included in guidelines for efficient pasture management. In our study, herbaceous production in the open untreated areas were around 2500 kg of DM ha–1y–1, far above the minimum
required for cattle maintenance (500–600 kg of DM ha–1y–1;
Christian, 1987), although during drier years lower
herbac-eous production would be obtained. Although minor N and P deficiencies were observed, nutrient concentrations of the whole herbaceous community were mostly within the range recommended for cattle nutrition (ARC, 1980). In any year or treatment, N and Ca concentrations of grasses (Tab. 3) did reach the level recommended for cattle nutrition (11.2 and 3 mg g–1, respectively; ARC, 1980). However, N and Ca
con-centrations of forbs were above the established require-ments. In the first year of study, grasses and forbs in all plots
showed P concentrations below the recommended level (2.5 mg g–1; ARC, 1980), while in the second year, P
concen-trations of fertilized plots were above this level. Similar effects of fertilization on P concentrations of Iberian pastures have been reported (Moreira and Ribeiro, 1990; García-Ciudad et al., 1997). In both study years, K and Mn concentrations in grasses and forbs were above the recommended level (6.0 and 0.04 mg g–1; ARC, 1980). Grasses contained
signifi-cantly lower nutrient concentrations than forbs, which coin-cides with the findings of a study carried out in Spanish “dehesas” (García-Ciudad et al., 1997).
5 Conclusions
Although grasses showed significantly lower nutrient concen-trations than forbs, herbaceous plants in Q. ilex montados appear to be suitable for cattle feeding, especially if the soil is fertilized for two consecutive years. A negative effect of shade on herbaceous-biomass production was observed, suggesting that pruning would favor herbaceous production. Water supply increased total herbaceous biomass irrespec-tive of the environment, suggesting that Q. ilex does not com-pete for soil-water resources with herbaceous plants in this ecosystem. Nutrient supply significantly increased total her-baceous biomass and herher-baceous P concentrations, espe-cially on the open grassland, supporting the traditional fertili-zation practices that farmers apply to the montados. Finally, higher K and Mg concentrations were observed in herbac-eous plants beneath the tree canopy than in those in the open areas, suggesting a positive effect of trees on pasture quality.
Acknowledgments
We thank the staff of Instituto de Ciências Agrárias
Mediterrâ-nicas (ICAM, Évora) for their valuable collaboration in field
work and Departamento de Ciências do Ambiente and Centro
de Pedología (Instituto Superior de Agronomia, Lisboa) for
analyzing the herbaceous nutrient concentrations and Dougal
Quin for the English revision. Comments by Profs. G. Moreno
and A. Solla greatly improved the manuscript. This study was supported by the project “Impacto da árvore e da pastagem melhorada na qualidade do solo e sustentabilidade dos mon-tados: Quantidades e dinâmicas do C e do N” (POCTI/AGG/
48914/2002), and by the activities of the Centre of Pedology (FCT, Plurianual Programme), through the subproject “Soil
management and sustainability of forestry and agroforestry systems” (SUSTAINFOR). Elena Cubera was awarded a postdoctoral fellowship by Consejería de Infraestructuras y
Desarrollo Tecnológico (Junta de Extremadura) and Fondo Social Europeo.
References
ARC (Agricultural Research Council) (1980): The Nutrient Require-ments of Ruminant Livestock. CAB, Farnham Royal, UK.
Campos, P., Rodríguez, Y., Caparrós, A. (2001): Towards the dehesa total income accounting: theory and operative Monfragüe study cases. Invest. Agrar.: Sist. Recur. For. Fuera de Serie 1, 43–62.
Chapman, H. D. (1965): Cation-exchange capacity, in Black, C. A.: Methods of Soil Analysis – Chemical and Microbiological Prop-erties. Agronomy 9, Am. Soc. Agron., Madison, WI, pp. 891–901. Christian, K. R. (1987): Matching pasture production and animal
requirements, in Wheeler, J. L., Pearson, C. J., Robards, G. E.: Temperate Pastures; Their Production, Use and Management. Australian Wool Corporation/CSIRO, Melbourne, pp. 463–476. Cubera, E., Moreno, G. (2007): Effect of single Quercus ilex trees
upon spatial and seasonal changes in soil water content in dehesas of central western Spain. Ann. For. Sci. 64, 355–364. Egnér, H., Riehm, H., Domingo, W. R. (1960): Untersuchungen über
die chemische Bodenanalyse als Grundlage für die Beurteilung des Nährstoffzustandes der Böden. II. Chemische Extraktions-methoden zur Phosphor- und Kaliumbestimmung. Kungl. Lantbr. Högsk. Ann. 26, 199–215.
Eichhorn, M. P., Paris, P., Herzog, F., Incoll, L. D., Liagre, F., Mantzanas K., Mayus, M., Moreno, G., Pilbeam, D. J. (2006): Silvoarable agriculture in Europe – past, present and future. Agrofor. Syst. 67, 29–50.
Escudero, A., García, B., Gomez, J. M., Luis, E. (1985): The nutrient cycling in Quercus rotundifolia and Quercus pyrenaica ecosystems (“dehesas”) of Spain. Acta Oecol.- Oec. Plant. 20, 73–86. Étienne, M. (1996): Western European Silvopastoral Systems.
Science Update series. INRA, Paris.
Gallardo, A. (2003): Effect of tree canopy on the spatial distribution of soil nutrients in a Mediterranean dehesa. Pedobiol. 47, 117–125. García-Ciudad, A., Ruano-Ramos, A., Vázquez-de-Aldana, B. R.,
García-Criado, B. (1997): Interannual variations of nutrients concentrations in botanical fractions from extensively managed grassland. Anim. Feed Sci. Technol. 66, 257–269.
Garrett, H. E., Kerley, M. S., Ladyman, K. P., Walter, W. D., Godsey, L. D., Van sambeek, J. W., Brauer, D. K. (2004): Hardwood silvo-pasture management in North America. Agrofor. Syst. 61, 21–33. Gea, G., Cañellas, I., Peña, M. B., Montero, G. (2007): Influencia de
la encina sobre la calidad bromatológica de comunidades herbáceas anuales en dehesas: Consecuencias sobre el manejo silvopastoral. Los Sistemas Forrajeros: entre la Producción y el Paisaje. Reunión científica XLVI, SEEP, Vitoria 4–8 Junio. González-Hernández, M. P., Silva-Pando, F. J., Casal Jiménez, M.
(1998): Production patterns of understory layers in several Galician (NW Spain) woodlands. Seasonality, net productivity and renewal rates. For. Ecol. Manage. 109, 251–259.
Holland, V. L. (1980): Effect of blue oak on rangeland forage production in central California, in Plumb, T. R. (tech. coord.): Ecology, Management and Utilization of California Oaks. USDA Gen. Tech. Report PSW-44, pp. 314–318.
IUSS Working Group (2006): World Reference Base for Soil Resources 2006 – A framework for international classification, correlation and communications. World Soil Resources, Reports 103, FAO, Rome.
Jackson, L. E., Strauss, R. B., Firestone, M. K., Bartolome, J. W. (1990): Influence of tree canopies on grassland productivity and nitrogen dynamics in deciduous oak savanna. Agric. Ecosyst. Environ. 32, 89–105.
Joffre, R., Rambal, S. (1988): Soil water improvement by trees in the rangelands of southern Spain. Oecol. Plant. 9, 405–422.
Joffre, R., Leiva Morales, M. J., Rambal, S., Fernandez Ales, R. (1987): Dynamique racinaire et extraction de l’eau du sol par des graminées pérennes et annuelles méditerranéennes. Acta Oecol.-Oec. Plant. 8, 181–194.
Marañón, T., Bartolome, J. W. (1993): Reciprocal transplants of herbaceous communities between Quercus agrifolia woodland and adjacent grassland. J. Ecol. 81, 673–682.
Marañón, T., Bartolome, J. W. (1994): Coast live oak (Quercus agri-folia) effects on grassland biomass and diversity. Madroño 41, 39–52.
Mcpherson, G. R. (1997): Ecology and Management of North American Savannas. University of Arizona Press, Tucson, p. 208. Moreno, G. (2008): Response of understorey forage to multiple tree
effects in Iberian dehesas. Agric. Ecosyst. Environ. 123, 239–244. Moreno, G., Obrador, J. J., Cubera, E., Dupraz, C. (2005): Fine root
distribution in Dehesas of central-western Spain. Plant Soil 277, 153–162.
Moreno, G., Obrador, J. J., García, E., Cubera, E., Montero, M. J., Pulido, F. J., Dupraz, C. (2007a): Driving competitive and facili-tative interactions in oak dehesas with management practices. Agrofor. Syst. 70, 25–40.
Moreno, G., Obrador, J., García, A. (2007b): Impact of evergreen oaks on soil fertility and crop production in intercropped dehesas. Agric. Ecosyst. Environ. 119, 270–280.
Moreira, O. M. S. C., Ribeiro, J. M. C. R. (1990): Mineral composition of some Portuguese pastures and its value for grazing ruminants. World Rev. Anim. Prod. 25, 69–72.
Parker, V. T., Muller, C. H. (1982): Vegetational and environment changes beneath isolated live oak trees (Quercus agrifolia) in a California annual grassland. Am. Midl. Nat. 107, 69–81.
Pérez-Corona, M. E., García-Criado, B., Vázquez-de-Aldana, B. R., García-Ciudad, A. (1994): Effect of topographic and temporal (maturity) gradients on the nutritive quality of semiarid herbaceous communities. Commu. Soil Sci. Plant Anal. 25, 2047–2061. Pérez-Corona, M. E., Vázquez-de-Aldana B. R., García-Criado, B.,
García-Ciudad, A. (1998): Variations in nutritional quality and biomass production of semiarid grasslands. J. Range Managem. 51, 570–576.
Provenza, F. D. (1995): Postingestive feedback as an elementary determinant of food preference and intake in ruminants. J. Range Manage. 48, 2–17.
Scholes, R. J., Archer, S. R. (1997): Tree-grass interactions in savannas. Annu. Rev. Ecol. Syst. 28, 517–544.
Thornthwaite, C. W. (1948): An approach towards a rational classifi-cation of climate. Geogr. Rev. 38, 55
Vázquez-de-Aldana, B. R., Pérez-Corona, M. E., García-Ciudad, A., García-Criado, B. (1996): Mineral content in semiarid grassland systems as affected by community structure and soil character-istics. Acta Oecol. 17, 245–259.
Vázquez-de-Aldana, B. R., García-Ciudad, A., Pérez-Corona, M. E., García-Criado, B. (2000): Nutritional quality of semi-arid grassland in western Spain over 10-year period: changes in chemical compo-sition of grasses, legumes and forbs. Grass Forage Sci. 55, 209–220.
Young, A. (1997): Agroforestry for Soil Management. CAB Interna-tional, Wallingford, p. 320.