Estudo do potencial adaptativo de diferentes espécies florestais em resposta
às alterações climáticas
Dissertação de Mestrado em Engenharia Florestal
Alexandra Manuela Silva Barros
Orientadora: Professora Doutora Maria João Magalhães Gaspar
Coorientadora: Professora Doutora Maria do Rosário Alves Ferreira dos Anjos
Estudo do potencial adaptativo de diferentes espécies florestais em resposta
às alterações climáticas
Dissertação de Mestrado em Engenharia Florestal
Alexandra Manuela Silva Barros
Orientadora: Professora Doutora Maria João Magalhães Gaspar
Coorientadora: Professora Doutora Maria do Rosário Alves Ferreira dos Anjos
Composição do Júri:
_____________________________________ _____________________________________ _____________________________________
“As doutrinas apresentadas neste trabalho são da exclusiva responsabilidade do seu autor.”
“Não há nenhum caminho tranquilizador à nossa espera. Se o queremos, teremos de construí-lo com as nossas mãos.”
pessoas, deixo, aqui, uma palavra de agradecimento a todos os que permitiram que este trabalho se realizasse, em especial:
À Professora Doutora Maria João Gaspar, minha orientadora, um sincero agradecimento pela disponibilidade, simpatia, incentivo e confiança que sempre depositou em mim ao longo de todo o trabalho, pela amizade, pelos ensinamentos transmitidos, por todas as palavras de conforto e pela atenção que sempre me deu.
À Professora Doutora Rosário Anjos, na qualidade de coorientadora, pela boa disposição que sempre demonstrou, pelos ensinamentos que me transmitiu, pela disponibilidade no acompanhamento do trabalho e pela constante preocupação.
Ao Engenheiro Délio e ao Paulo Ventura, técnicos do Departamento de Engenharia Florestal, por todo o apoio, pela cedência de bibliografia, por estarem sempre disponíveis a ajudar-me e pelo carinho que sempre demonstraram. Agradeço também ao Carlos Fernandes pela sua prontidão em colaborar nas idas ao campo.
À Mestre Helena Ferreira, pela paciência, pela valiosa ajuda e pelos esclarecimentos prestados no laboratório, assim como à Rita e ao Rodrigo, pelas risadas e pelos bons momentos passados no campo.
Ao Engenheiro António Correia, um agradecimento especial, pela ajuda disponibilizada no tratamento estatístico dos dados e por estar sempre disponível para me esclarecer qualquer duvida.
Ao Doutor João Paulo Coutinho, pelo importante contributo na versão final desta dissertação.
Aos meus pais, em especial à minha mãe, por todo o apoio e compreensão que sempre demonstrou, pelos conselhos e por fazer com que me tornasse na pessoa que sou hoje.
Aos meus irmãos e avós, por sempre terem acreditado que chegaria ao fim desta etapa com sucesso, pelo orgulho que sempre tiveram em mim e pelas palavras de encorajamento nos momentos de fraqueza.
Ao meu namorado, pela paciência, pelo companheirismo, pela força e apoio que sempre me transmitiu.
Ao meu amigo Sérgio por ter estado sempre presente nos bons e nos maus momentos ao longo destes cinco anos e à Tatiana pela pessoa fantástica que é e por me ter ajudado sempre.
Para finalizar, gostaria de agradecer a todos os meus amigos, que por fazerem parte da minha vida contribuíram significativamente para a realização desta dissertação.
xi
século XXI. Em consequência dessas alterações são de esperar fortes impactos nas espécies florestais, nomeadamente: elevada taxa de mortalidade nos limites da atual distribuição geográfica; alteração na produtividade e qualidade; maior incidência de pragas e doenças, bem como migração e extinção de espécies importantes. Assim, a capacidade adaptativa torna-se num pré-requisito para a sobrevivência sob condições climáticas mais adversas no futuro. Neste contexto, surgiu o projeto REINFFORCE, que ofereceu a oportunidade de instalar uma rede de arboretos capaz de monitorizar o efeito das alterações climáticas na floresta Atlântica e testar a eficiência de medidas de adaptação em 35 espécies florestais. Para o efeito, foi instalado o mesmo material genético em 38 climas distintos desde a Escócia até Portugal Continental e Açores. No presente trabalho apresentam-se os resultados obtidos, três anos após a instalação do arboreto na Universidade de Trás-os-Montes e Alto Douro. O principal objetivo foi avaliar o comportamento de nove proveniências de três espécies florestais (Betula pendula Roth,
Quercus robur L. e Quercus suber L.) quanto à taxa de sobrevivência, crescimento, fenologia
e parâmetros bioquímicos, relacionando o seu comportamento com a origem do material de propagação. Os resultados obtidos mostraram variabilidade na adaptabilidade entre as espécies e proveniências. Ao nível da sobrevivência, destaca-se a espécie Q. suber com maior percentagem de plantas vivas. Contudo, a proveniência de origem portuguesa (Alca) revelou-se menos adaptada às condições edafoclimáticas do local com maior percentagem de plantas mortas. Adicionalmente, esta espécie, apresentou acréscimos médios anuais em altura e em diâmetro mais baixos, bem como as suas proveniências, designadamente as proveniências espanhola (Esp) e francesa (Pyre) na variável altura e as proveniências portuguesa (Alca) e espanhola (Esp) na variável diâmetro. Por sua vez, a B. pendula e respetivas proveniências registaram os valores mais elevados de acréscimos em altura e em diâmetro, sobretudo a proveniência francesa (Nord). Além disso, demonstraram um maior avanço fenológico, especialmente a proveniência eslovaca (Kral). A espécie mais tardia no abrolhamento e na maturação foliar foi a Q. suber, bem como as proveniências portuguesa (Alca) e francesa (Land). No que respeita aos parâmetros bioquímicos verificou-se que a B. pendula apresentou teores mais elevados em todos os compostos quantificados, à exceção dos compostos fenólicos que foram superiores na Q. suber, particularmente na proveniência francesa (Land). Por conseguinte, este estudo permitiu perceber quais as respostas das diferentes proveniências de cada espécie às condições edafoclimáticas atuais no arboreto. Estes resultados serão uma
xii
ferramenta importante na gestão sustentável da floresta mediterrânica, principalmente em ações de reflorestação.
Palavras-chave: alterações climáticas, proveniências, sobrevivência, crescimento, fenologia,
xiii
As a result of these changes, strong impacts on forest species are expected, in particular: high mortality rate within the limits of the current geographic distribution; changes in productivity and quality; higher incidence of pests and diseases, as well as migration and extinction of important species. Therefore, the adaptive capacity becomes a prerequisite for survival under adverse weather conditions in the future. In this context, REINFFORCE project was implemented, offering the opportunity to install an arboretum network capable of monitoring the effect of climate changes on the Atlantic forest and test the efficiency of adaptation measures in 35 forest species. For this purpose, the same genetic material was installed in 38 different climates from Scotland to Continental Portugal and Azores. This study presents the results obtained three years after the arboretum installation at University of Trás-os-Montes e Alto Douro. The main objective was to evaluate the behaviour of nine provenances of three forest species (Betula pendula Roth, Quercus robur L. and Quercus suber L.) in terms of survival rate, growth, phenology and biochemical parameters, in order to relate their behaviour to the origin of the propagating material. The obtained results showed variability in the adaptability between species and provenances. Regarding the level of survival, Q. suber species stands out with the highest percentage of live plants. However, the Portuguese provenance (Alca) was less adapted to the edaphoclimatic of the site, with higher percentage of dead plants. In addition, this species presented lower annual average increases in height and diameter, as well as its provenances, namely the Spanish (Esp) and French (Pyre) for diameter. On the other hand, B. pendula and its provenances registered the highest increases in height and diameter, especially the French provenance (Nord). In addition, these species presented a greater phenological advance, especially the Slovak provenance (Kral). The species with the later sprouting and maturation was Q. suber, namely the Portuguese (Alca) and French (Land) provenances. Concerning the biochemical parameters, it was verified that B. pendula had higher levels in all the quantified compounds, except for the phenolic compounds that were superior in Q. suber, particularly in the French provenance (Land). Therefore, this study allowed us to perceive the responses of the different provenances of each species to the current edaphoclimatic conditions in the arboretum. These results will be an important tool in the sustainable management of the Mediterranean forest, especially in reforestation actions.
xiv
Keywords: climate changes, provenances, survival, growth, phenology, biochemical parameters.
xv
2. Caracterização das espécies em estudo ... 5
2.1. Betula pendula Roth. ... 5
2.1.1. Características ecológicas ... 7 2.1.2. Distribuição geográfica ... 8 2.2. Quercus robur L. ... 8 2.2.1. Características ecológicas ... 10 2.2.2. Distribuição geográfica ... 10 2.3. Quercus suber L. ... 12 2.3.1. Características ecológicas ... 14 2.3.2. Distribuição geográfica ... 14 3. Fenologia ... 17 4. Parâmetros bioquímicos ... 19 4.1. Pigmentos fotossintéticos ... 19 4.2. Compostos fenólicos ... 20
4.3. Açúcares solúveis e amido ... 21
4.4. Proteínas solúveis ... 21
5. Material e métodos ... 23
5.1. Desenho experimental e material vegetal ... 23
5.2. Parâmetros avaliados ... 26
5.2.1. Taxa de sobrevivência ... 26
5.2.2. Crescimento ... 26
5.2.3. Fenologia foliar ... 28
5.2.4. Análises bioquímicas ... 28
5.2.4.1. Recolha e conservação do material vegetal ... 28
5.2.4.2. Determinação do teor em pigmentos fotossintéticos ... 29
5.2.4.3. Determinação dos compostos fenólicos totais ... 30
5.2.4.4. Determinação dos açúcares solúveis e amido ... 30
5.2.4.5. Determinação das proteínas solúveis ... 31
5.3. Análise estatística ... 32
6. Análise e discussão dos resultados ... 35
6.1. Sobrevivência ... 35
xvi
6.1.2. Taxa de sobrevivência por proveniência ... 36
6.2. Acréscimo em altura ... 38
6.2.1. Acréscimo médio anual em altura por espécie ... 38
6.2.2. Acréscimo médio anual em altura por proveniência ... 40
6.3. Acréscimo em diâmetro ... 41
6.3.1. Acréscimo médio anual em diâmetro por espécie ... 41
6.3.2. Acréscimo médio anual em diâmetro por proveniência ... 41
6.4. Fenologia foliar ... 42
6.4.1. Evolução fenológica por espécie ... 42
6.4.2. Evolução fenológica por proveniência ... 46
6.5. Análises bioquímicas ... 49
6.5.1. Análise bioquímica dos compostos quantificados por espécie ... 49
6.5.2. Análise bioquímica dos compostos quantificados por proveniência ... 52
7. Conclusões ... 55
Referências bibliográficas ... 57
xvii
Figura 2 - Detalhes da espécie B. pendula com três anos: (a) folhas; (b) amentilhos; (c)
pormenor da coloração esbranquiçada do ritidoma. ... 6
Figura 3 - Exemplar da Q. robur. ... 9
Figura 4 - Distribuição da Q. robur em Portugal.. ... 11
Figura 5 - Exemplar da Q. suber no estado adulto. ... 12
Figura 6 - Área de distribuição da Q. suber com destaque para a distribuição em Portugal.. . 15
Figura 7 - Algumas etapas do ciclo fenológico da Q. suber num período de seis semanas. ... 17
Figura 8 - Localização das parcelas do arboreto REINFFORCE na UTAD. ... 23
Figura 9 - Localização geográfica das nove proveniências em estudo. ... 24
Figura 10 - Medições efetuadas em Q. suber: (a) em altura (cm) com recurso a uma vara graduada (b) em diâmetro (mm) com auxílio de um paquímetro digital. ... 27
Figura 11 - Ramo da Q. suber marcado para avaliação da fenologia. ... 28
Figura 12 - Taxa de sobrevivência (%), por espécie, observada em outubro de 2015, três anos após a instalação do arboreto. ... 36
Figura 13 - Taxa de sobrevivência (%), por proveniência, observada em outubro de 2015, três anos após a instalação do arboreto.. ... 37
Figura 14 - Acréscimo médio anual em altura (cm) e respetivo erro padrão, por espécie, três anos após a plantação. ... 39
Figura 15 - Acréscimo médio anual em altura (cm) e respetivo erro padrão, por proveniência, três anos após a plantação. ... 40
Figura 16 - Acréscimo médio anual em diâmetro (mm) e respetivo erro padrão, por espécie, três anos após a plantação.. ... 41
Figura 17 - Acréscimo médio anual em diâmetro (mm) e respetivo erro padrão, por proveniência, três anos após a plantação. ... 42
Figura 18 - Número de dias e respetivo erro padrão, observado no campo, que as espécies em estudo demoraram a atingir o estádio 19 (maturação foliar). ... 45
Figura 19 - Número de dias e respetivo erro padrão, observado no campo, que as proveniências da B. pendula demoraram a atingir o estádio 19 (maturação foliar). ... 46
Figura 20 - Número de dias e respetivo erro padrão, observado no campo, que as proveniências da Q. robur demoraram a atingir o estádio 19 (maturação foliar). ... 47
xviii
Figura 21 - Número de dias e respetivo erro padrão, observado no campo, que as proveniências da Q. suber demoraram a atingir o estádio 19 (maturação foliar). ... 48 Figura 22 - Teores médios e respetivo erro padrão em: (a) pigmentos clorofilinos (Cl a, Cl b e carotenoides); (b) açúcares solúveis, amido, proteínas solúveis e compostos fenólicos, por espécie... ... 51 Figura 23 - Teores médios e respetivo erro padrão de pigmentos fotossintéticos (a, b, c), açúcares solúveis (d), amido (e), proteínas solúveis (f) e compostos fenólicos (g), por proveniência. ... 53
xix
e proveniência, país da colheita das sementes, região de proveniência, variáveis geográficas e principais características climáticas.. ... 25 Tabela 2 - Resumo da análise de variância da taxa de sobrevivência por espécie e por proveniência. ... 35 Tabela 3 - Resumo da análise de variância dos acréscimos médios anuais em altura (cm) e em diâmetro (mm) por espécie e por proveniência. ... 39 Tabela 4 - Resumo da análise de variância por espécie e por proveniência referente aos estádios 02, 07, 10 e 19. ... 43 Tabela 5 - Número médio de dias ± erro padrão para os estádios 02, 07, 10 e 19 por espécie e por proveniência. ... 44 Tabela 6 - Resumo da análise de variância para os pigmentos fotossintéticos (Cl a, Cl b e carotenoides), açúcares solúveis, amido, proteínas solúveis e compostos fenólicos, por espécie e por proveniência. ... 50
xxi Ø - diâmetro ºC - graus Celsius μg - microgramas μL - microlitros Abs - absorvância
ANOVA - Analysis of variance
BSA - Bovine serum albumin
Car - carotenoides
Cl - clorofila
cm - centímetros
CO2 - dióxido de carbono
EDTA - Ethylenediamine tetraacetic acid
g - gramas
ha - hectares
IFN - Inventário Florestal Nacional
K - potássio km - quilómetros m - metros mL - mililitros mm - milímetros N - azoto nm - nanómetros
xxii P - fósforo
pH - potencial de hidrogénio
PMSF - phenylmethylsulfonyl fluoride
ppm - partes por milhão
PS - peso seco
PVPP - Polyvinylpyrrolidone insoluble
REINFFORCE - REsource INFrastructures for monitoring, adapting and protecting european atlantic FORests under Changing climatE
RNAP - Rede Nacional de Áreas Protegidas
rpm - rotações por minuto
UTAD - Universidade de Trás-os-Montes e Alto Douro
1
1. Introdução
As florestas cobrem aproximadamente 42 milhões de km2 em regiões tropicais, temperadas e boreais, ou seja, cerca de 30% da superfície terrestre (Bonan, 2008). Reconhecemo-las como um recurso natural que sustém numerosos setores socioeconómicos e providencia uma larga extensão de bens e serviços culturais (Davidson et al., 2003; Morison & Morecroft, 2006; Ashraf et al., 2015), incluindo refúgio para a biodiversidade, fornecimento de comida, produtos florestais e medicinais, regulação do ciclo hidrológico, usos recreativos, valores estéticos, etc. (Bonan, 2008). Além da importância para a ecologia do planeta, as florestas sempre tiveram grande valor para a humanidade. A madeira é o material de construção por excelência, sendo utilizada como matéria-prima industrial e artesanal e como combustível (Pereira, 2014).
As alterações climáticas tornaram-se num dos maiores desafios que o setor florestal enfrenta no século XXI (DGRF, 2006; Hanewinkel & Peyron, 2014). Muitos fatores, bióticos e abióticos, podem determinar a resiliência das florestas a estas mudanças de clima (Dale et al., 2009), podendo levar a modificações significativas em habitats naturais e consequentemente, no desenvolvimento normal de espécies localmente adaptadas (Alfaro et al., 2014). Assim, o clima está entre os fatores mais importantes que controlam a distribuição das espécies florestais (Gonzalez et al., 2010; Lindner et al., 2010). As florestas são particularmente sensíveis às alterações climáticas porque as árvores têm vida longa e crescimento lento, o que dificulta adaptações rápidas (Lindner et al., 2010). Porém, ao longo dos milénios, as florestas têm-se adaptado continuamente a estas mudanças (Davis et al., 2005; Carcaillet et al., 2011).De acordo com Araújo & Rahbek (2006), são diversas as respostas que os organismos adotam face a estas alterações: mudanças na distribuição geográfica (Chuine & Beaubien, 2001) e migrações (Pereira, 2014), na dinâmica populacional (Stenseth et al., 2002), no melhoramento dos processos evolutivos (Bradshaw & Holzapfel, 2006) ou no ajuste dos seus ciclos de vida (Sparks & Menzel, 2002). A migração (processo lento) e a adaptação (mais rápida) são as respostas alternativas a estes desafios, uma vez que esta evolução permite às populações adaptarem-se às novas condições (Aitken et al., 2008).
As alterações climáticas também podem danificar as florestas, especialmente aquelas que já se encontram sob condições de stresse, ao alterarem o regime de fogo (Guiomar et al., 2015), desencadeando pragas e doenças (Dukes et al., 2009; Sturrock et al., 2011), favorecendo o estabelecimento de espécies invasoras ou induzindo perdas na biodiversidade (Afreen et al., 2011). Por outro lado, as florestas podem ajudar a mitigar as alterações climáticas, através de
1. Introdução
2
trocas de energia, fixando o carbono, suportando biocombustíveis ou prevenindo a erosão e desertificação do solo (Bonan, 2008; Aaheim et al., 2011). De facto, as emissões de gases atmosféricos são a principal causa antropogénica das alterações climáticas atuais (Koskela et
al., 2007). Desde o início da revolução industrial, em meados do século XVIII, que as emissões
de CO2 para a atmosfera, resultantes da combustão dos combustíveis fósseis (carvão, petróleo
e gás natural) e da desflorestação, contribuíram para um aumento de 30% na concentração atmosférica de CO2 (Pereira et al., 2005). A taxa de aumento na concentração do teor de CO2
na atmosfera é de aproximadamente 0,5% ao ano, tendo passado de 315,7 ppm (partes por milhão) em 1958 para 400 ppm em 2013 (Pereira, 2014). Atualmente, as florestas armazenam cerca de 45% do carbono terrestre e, ao contribuírem com cerca de 50% da produção primária líquida, podem sequestrar grandes quantidades de carbono (Bonan, 2008).
Portugal é um dos países europeus com maior área de floresta em relação à área total do território (Pereira, 2014). De acordo com os dados publicados em 2010 pelo 6.º Inventário Florestal Nacional (IFN), a floresta continental ocupa cerca de 35,4% do território, tendo neste século sofrido um decréscimo a uma taxa de 0,3% por ano. Importa referir que, atualmente, as espécies florestais com maior representatividade e interesse económico são o pinheiro-bravo (Pinus pinaster Ait.), o eucalipto (Eucalyptus globulus Labill.) e o sobreiro (Quercus suber L.), que representam cerca de 74% da área florestal existente.
Em Portugal predominam quatro tipos de florestas: (1) os montados, em clima tipicamente mediterrânico, composto por espécies autóctones (sobreiro, azinheira e pinheiro-manso), com baixa densidade de árvores e uso agroflorestal; (2) as florestas plantadas para produção de madeira com espécies autóctones, como o pinheiro-bravo em regime de alto fuste; (3) os povoamentos submetidos a uma silvicultura intensiva, explorados em regime de talhadia de rotações curtas e obtidos artificialmente através da instalação de espécies exóticas como o eucalipto ou de espécies nativas como o castanheiro; e (4) as florestas de regeneração natural. Alguns destes espaços apresentam alto valor de conservação da biodiversidade sendo que aproximadamente 18,7% da área de floresta do continente está integrada na Rede Nacional de Áreas Protegidas (RNAP) e 23% na Rede Natura 2000 (Pereira, 2014).
No que se refere ao clima português, as características mediterrânicas prevalecem ao longo do país com forte sazonalidade (estações quentes e secas e frias e húmidas), precipitação irregular e seca no verão. Na região Norte do país predominam as florestas e os bosques com maior densidade, enquanto a parte mais a Sul é dominada pela agricultura e por áreas extensas de montado (Godinho-Ferreira et al., 2005).
3
As florestas localizadas no Sul da Europa deverão ser afetadas pelas alterações climáticas, uma vez que é apontada como uma das regiões mais vulneráveis do planeta (IPCC, 2007). Em Portugal Continental as temperaturas máximas e mínimas subiram, desde os anos setenta, cerca de 0,5ºC por década, o que corresponde a mais do dobro do aumento verificado na temperatura média mundial. Ao mesmo tempo, esta alteração foi acompanhada pela diminuição da precipitação total e de um acréscimo da frequência e intensidade de secas. Outra das alterações climáticas previstas e já observada é o aumento da frequência dos fenómenos extremos, como cheias, tempestades, secas e ondas de calor (Santos & Miranda, 2006).
Neste contexto surge o projeto REINFFORCE (REsource INFrastructures for
monitoring, adapting and protecting European Atlantic FORests under Changing climatE), que
ofereceu a oportunidade de instalar uma rede única no mundo, com o intuito de monitorizar o efeito das alterações climáticas na floresta Atlântica e melhorar o conhecimento sobre a capacidade de adaptação das espécies florestais. Para o efeito, foi instalado o mesmo material genético em 38 arboretos com diferentes climas desde a Escócia até Portugal Continental e Açores (entre as Latitudes 37º até 58ºN). Em Portugal foram instalados três arboretos no Continente (Lisboa - Tapada da Ajuda; Sintra - Parque Natural; e Vila Real - Quinta de Prados) e três no arquipélago dos Açores (ilha de S. Miguel). Este projeto torna-se fundamental para a sustentabilidade dos recursos florestais atlânticos, já que se está a plantar o que irá ser colhido no futuro, sob novas condições climáticas (Orazio et al., 2013).
Nesta perspetiva, este estudo apresenta os resultados obtidos, três anos após a instalação do arboreto, tendo como principal objetivo estudar a capacidade adaptativa de nove proveniências de três espécies florestais, nomeadamente Betula pendula, Quercus robur e
Quercus suber, instaladas num arboreto da Universidade de Trás-os-Montes e Alto Douro
(UTAD), Vila Real. O trabalho realizado consistiu na avaliação da taxa de sobrevivência, crescimento, fenologia e parâmetros bioquímicos, cujo objetivo final é determinar quais as proveniências de cada espécie que melhor se adaptam às condições edafoclimáticas atuais do arboreto, contribuindo, assim, para o estabelecimento de povoamentos sãos e produtivos no futuro.
1. Introdução
5
2. Caracterização das espécies em estudo
2.1. Betula pendula Roth.
A espécie Betula pendula, representada na Figura 1, foi reconhecida em 1788 por Roth e é vulgarmente conhecida por bétula, bidoeiro ou vidoeiro (Bingre et al., 2007). Esta espécie pertence à divisão Spermatophyta, subdivisão Magnoliophyta, classe Magnoliopsida, ordem Fagales, família Betulaceae e ao género Betula (Franco, 1971).
Figura 1 - Exemplar da B. pendula, com três anos, no arboreto da UTAD.
Quanto às suas características botânicas é uma árvore de médio porte, atinge alturas de 30 m e vive até cerca dos 100 anos (Humphries & More, 1996). O seu crescimento é relativamente rápido até cerca dos 30 anos e depois diminui até cessar por volta dos 50 anos (Ferreira & Jiménez, 2003). Depois disso a vitalidade das árvores diminui e tornam-se mais suscetíveis à deterioração e a outros defeitos (Hynynen et al., 2010). Esta espécie exibe uma copa estreita e aguda quando jovem, tendo tendência a alargar com a idade. Nas árvores jovens, os ramos são eretos, tornando-se pendentes com o aumento da idade, e os raminhos pendentes, delgados, glabros e castanhos, com inúmeras glândulas (Humphries & More, 1996).
As suas folhas (Figura 2a) medem 2-7 cm de comprimento, são ovado triangulares, agudas e acuminadas, finas e mais ou menos glabras quando maturas, exibindo margens
2. Caracterização das espécies em estudo
6
duplamente serradas, com dentes primários salientes separados por numerosos dentes secundários. Com cerca de 3-6 cm de comprimento, os amentilhos (Figura 2b) encontram-se agregados de dois a quatro na extremidade dos raminhos. No inverno são acastanhados, tornando-se amarelados e maturando com as folhas jovens. Os amentilhos femininos, inicialmente esverdeados, surgem nas axilas das folhas, tornando-se castanhos. As infrutescências (com 1,5-3,5 cm de comprimento) apresentam escamas com dois lobos laterais largos e curvos e um lobo central triangular. A asa da sâmara é 2-3 vezes mais larga que o corpo (Humphries & More, 1996).
O ritidoma (Figura 2c) é liso, branco prateado em cima e escuro na base, onde surge fissurado, formando placas retangulares. Por vezes, apresenta ranhuras escamosas evidenciando uma coloração cinza claro (Humphries & More, 1996).
(a)
(b) (c)
Figura 2 - Detalhes da espécie B. pendula com três anos: (a) folhas; (b) amentilhos; (c) pormenor da coloração
esbranquiçada do ritidoma.
A floração acontece de abril a maio e a maturação dos amentilhos ocorre entre o final do verão e o início do outono. A frutificação inicia-se a partir dos 8-10 anos, no entanto, em alguns casos, pode expressar-se num prazo de cinco anos (Reyes et al., 1997). A dispersão do pólen, que é principalmente anemófila, mas podendo ser também entomófila, decorre até final do
7
outono. A semente alcança grandes distâncias e é favorecida pelos tegumentos impermeáveis que permitem a sua flutuação durante meses antes da germinação. Quando começa a produzir semente, a tendência é uma frutificação anual estável, a não ser que as condições meteorológicas sejam desfavoráveis, e na época da polinização haja perdas. No entanto, a produção de semente varia de ano para ano, ocorrendo a cada três anos um pico na quantidade produzida. Em anos favoráveis, a quantidade de sementes é muito elevada, embora a sua viabilidade tenha tendência a ser baixa, bem como a sobrevivência dos novos rebentos (Reyes
et al., 1997).
Os fatores que influenciam o número de sementes produzidas, bem como a sua qualidade são diversos. Entre eles podem citar-se: (1) características da população, devido à existência de uma relação direta entre o diâmetro e a altura média, quantidade de sementes produzida e percentagem de germinação (Reyes et al., 1997); (2) altitude das populações, porque à medida que a altitude aumenta, diminui o número de sementes em cada amentilho, a qualidade antes da dispersão e, por conseguinte, a capacidade germinativa; (3) clima, visto que a germinação no campo depende da existência de um período de frio no inverno e de um aumento da temperatura na primavera, pois normalmente a qualidade das sementes é maior nos anos em que as primaveras são mais quentes (Perala & Alm, 1990); e (4) período de dispersão, porque à medida que o período de frutificação e disseminação avançam, a quantidade de sementes e a sua viabilidade diminui (Reyes et al., 1997).
O sistema radicular varia de acordo com as condições edáficas, ou seja, em locais secos desenvolvem-se raízes profundas, enquanto em solos húmidos o sistema radicular é mais superficial (Reyes et al., 1997).
2.1.1. Características ecológicas
Tendo frequentemente um papel de colonizadora, a B. pendula comporta-se de forma muito positiva como espécie pioneira, melhorando as condições edáficas do solo com vista à instalação posterior de outras espécies mais exigentes. É uma espécie muito resistente ao frio, neve, gelo e ventos fortes. Pode resistir, também, a temperaturas muito elevadas no verão, sempre que haja uma compensação hídrica freática. Normalmente é uma espécie bastante plástica quanto à natureza do solo e possui um amplo espectro ecológico sendo, contudo, exigente no que respeita à humidade (Loureiro, 2005). Tem a capacidade de colonizar rapidamente áreas onde o solo é pobre em nutrientes, mas preferindo solos com pH ácido (Lewis
2. Caracterização das espécies em estudo
8
et al. 2015). No entanto, sendo esta uma espécie de luz (não cresce à sombra) pode ser
eventualmente substituída por outras espécies (Bingre et al., 2007).
2.1.2. Distribuição geográfica
De acordo com Correia & Oliveira (2003) a B. pendula tem uma vasta área de origem, abrangendo toda a Europa, com exceção da Grécia e todo o Norte da Ásia até à península de Kamchatka (região oriental da Rússia). No Sul da Europa a sua distribuição é local e restringida às regiões de montanha (Humphries & More, 1996). Em Portugal aparece principalmente nas serras do Norte e Centro marginando as linhas de água e nas encostas húmidas, onde forma pequenos bosques (Loureiro, 2005).
2.2. Quercus robur L.
A espécie Quercus robur L. (Figura 3), validada por Lineu em 1753, é usualmente denominada carvalho-roble, carvalho-alvarinho ou carvalho-comum (Silva, 2007a). Pertence à divisão Spermatophyta, à subdivisão Magnoliophyta, à classe Magnoliopsida, à ordem Fagales, à família Fagacea e ao género Quercus (Franco, 1971).
No que respeita às características botânicas, é um dos carvalhos com maior porte, atingindo uma altura de 30-40 m, de copa ampla e regular (Humphries & More, 1996) com ramificação tortuosa e com diversos ramos grossos, sendo que a sua forma está dependente das condições de crescimento. Quando juvenil, a casca é lisa e de coloração acinzentada, com tendência a tornar-se grossa e fendida longitudinalmente com o passar dos anos. Por vezes, cobre-se de rebentos por todo o tronco. As folhas membranosas não contêm tricomas e têm habitualmente dimensões que rondam os 6-12 × 3-6 cm. Estas possuem 4-8 pares de lóbulos redondos e um pecíolo reduzido (cerca de 2-7 mm de diâmetro). Inicialmente axial, o seu sistema radicular forma, com a idade, raízes secundárias a níveis profundos, concedendo-lhe grande capacidade de resistência ao vento (Silva, 2007a).
9
Figura 3 - Exemplar da Q. robur [1].
A floração monoica ocorre em abril e é coincidente com o aparecimento das folhas. Contudo, de modo a evitar a autofecundação, as florações (feminina e masculina) estão desfasadas no tempo. As flores femininas são arredondadas e de pequenas dimensões, estendem-se sobre um longo pedúnculo, contendo geralmente três estigmas de coloração arroxeada. Estas flores não são atrativas para os insetos, uma vez que a sua polinização é feita por ação do vento (anemófila). As flores masculinas dispõem-se em amentilhos delgados, agrupados na extremidade dos ramos. A maturação do fruto ocorre no outono do mesmo ano da floração e consiste numa glande ovoide de cor acastanhada inserida numa cápsula basilar, vulgarmente conhecida por bolota.
Esta espécie só consegue produzir fruto em abundância e com qualidade a partir dos 60 anos, ou seja, quando atinge a maturidade, refletindo-se no desenvolvimento das suas inflorescências. O facto da qualidade e quantidade do fruto diferir ao longo dos anos deve- -se não só às alterações climáticas, mas também a outros fatores externos, nomeadamente à disponibilidade de reservas, às características genéticas e aos ciclos de predação de insetos. A cada 2-6 anos a Q. robur frutifica abundantemente, chegando a produzir ao longo do seu ciclo de vida, mais de três milhões de bolotas. A disseminação das sementes é feita através de aves e outros animais, que as dispersam noutros locais.
2. Caracterização das espécies em estudo
10
As florestas de carvalhos desempenham um papel fundamental para a manutenção da fauna silvestre, contribuindo, não só como fonte de alimento, como também na criação de múltiplos recantos para vários animais que vivem associados aos carvalhais. Além disso, albergam variadas plantas produtoras de frutos de interesse para a fauna (Silva, 2007a).
2.2.1. Características ecológicas
Segundo Silva (2007a), aquando do estabelecimento de uma floresta de carvalho e do seu respetivo modo de gestão é fundamental ter em consideração as características do clima e do solo nesse mesmo local.
Trata-se de uma espécie dependente de condições climáticas temperadas, que prospera em baixas altitudes, podendo em alguns locais subir até aos 800-1000 m como nas regiões do Gerês e Barroso. Necessita de uma pluviosidade anual entre os 1000 e 1800 mm podendo, em certas regiões mais quentes, beneficiar de precipitações acima dos 2000 mm. Outra característica climática que se associa a esta espécie é a baixa tolerância a frios invernais intensos ou a geadas tardias. De acordo com Monteiro (2010), o carvalho roble vegeta numa larga gama de solos, desde que sejam frescos, não calcários e com boa capacidade de retenção de água.
2.2.2. Distribuição geográfica
Segundo Bevia & Resines (2005), o carvalho-alvarinho é o carvalho mais difundido na Europa. Distribui-se vastamente na Europa atlântica e central, abrangendo as zonas temperadas a leste do Cáucaso. Na Península Ibérica ocorre sobretudo na metade Noroeste, principalmente em regiões com influência climática atlântica (Silva, 2007a).
Em Portugal Continental, conforme se pode observar na Figura 4, esta espécie centra-se essencialmente nas regiões do noroeste português, abrangendo as províncias do Minho, Douro litoral e Beira litoral.
11
Figura 4 - Distribuição da Q. robur em Portugal. Fonte: Silva (2007a). Dada a escala em que o mapa foi
cartografado existem pequenas manchas que não são observáveis.
A sua área de ocorrência potencial (representada a verde-claro na Figura 4) é onde esta espécie encontra condições ecológicas ótimas para surgir de forma espontânea, aparecendo frequentemente dispersa ou em pequenos núcleos, por exemplo em terrenos pedregosos ou com declives acentuados, junto a linhas de água, caminhos e berma de estradas, ou ainda em povoamentos dominados por outras espécies florestais.
Ainda na Figura 4, as manchas verde-escuras referem-se a povoamentos puros ou mistos com Quercus pyrenaica Willd. ou Quercus faginea Lam. nalgumas manchas mais a sul. Noutros casos, existem zonas de sobreposição onde as espécies podem coexistir numa mesma área. No nosso país existem dois bosques de referência para esta espécie, na Mata de Albergaria (Parque Nacional da Peneda-Gerês) e em Corno do Bico (Paredes de Coura). A Mata de Albergaria é constituída predominantemente por carvalhal, tratando-se de uma encosta extensa e abrigada com exposição a norte. Face às suas características ecológicas (estrutura, composição
2. Caracterização das espécies em estudo
12
florística e grau de preservação), pode-se assumir como sendo um bosque antigo, albergando uma enorme diversidade em fauna. Corno do Bico é outra área com grande relevância que, apesar de no passado ter sofrido algum grau de intervenção silvícola, nomeadamente a plantação de Quercus borealis L., é onde ainda se pode encontrar uma das maiores manchas contínuas e bem conservadas de carvalho-alvarinho. Dada a elevada importância para a conservação de uma variedade de habitats e espécies ameaçados, atualmente é uma área de Paisagem Protegida e um Sítio de Importância Comunitária da Rede Natura 2000 (Silva, 2007a).
2.3. Quercus suber L.
O sobreiro (Figura 5), que Lineu batizou com o nome latino de Quercus suber pertence à família Fagacea e ao género Quercus. Entre indivíduos são verificadas diferenças morfológicas muito marcadas, em consequência da sua complexa constituição genética (AGRO.GES, 2000). Esta espécie de grande longevidade e de crescimento lento pode, em alguns casos, atingir 250-350 anos, ainda que se considere que a idade limite para a produção de cortiça seja de 150-200 anos (Fortes et al., 2006). Normalmente, um sobreiro adulto tem um porte natural de 10-15 m de altura, possuindo um ritidoma suberoso, espesso e fendido que se bifurca a uma altura reduzida do solo, cerca de 4 m (Franco, 1971).
13
É uma espécie de folha persistente, assim como a azinheira (Q. rotundifólia L.), distinguindo-se de outros carvalhos portugueses de folha caduca, como Q. faginea Lam. (carvalho-português), Q. pyrenaica Willd. (carvalho-negral) e Q. robur L. (carvalho-alvarinho). A página superior das folhas apresenta uma cor verde-escura e na página inferior, tende a ser esbranquiçada e tomentosa. De árvore para árvore, a forma e a dimensão das folhas é variável, podendo ser ovadas, ovadas-lanceoladas, oblongas ou serradas. No início da primavera nascem as primeiras folhas, que se desenvolvem até junho. Geralmente, as folhas têm uma duração média de 1-2 anos, mas grande parte da folhagem cai na primavera (Silva, 2007b).
A época principal de floração dá-se durante um período relativamente longo, de abril até junho conforme as regiões e as condições edafoclimáticas do local. Em Portugal, nas regiões do centro e sul do país a floração normalmente inicia-se em abril/maio, podendo, em algumas regiões do norte do país, estender-se até junho/julho. Sendo uma espécie que apresenta dicogamia, as inflorescências femininas surgem mais tarde que as masculinas, sendo que as primeiras se encontram nas axilas das folhas, na parte média dos lançamentos anuais dos ramos dentro de um invólucro de escamas que dará origem à cúpula do fruto. As flores masculinas situam-se nas extremidades dos lançamentos dos ramos do ano anterior ou na base de raminhos do próprio ano, formando amentilhos pendentes e em grupos (Silva, 2007b).
Na primavera ocorre a polinização, com pólen proveniente dos próprios indivíduos e de árvores vizinhas. O sobreiro, sendo uma espécie anemófila, a dispersão do pólen é muitas vezes condicionada por fatores climáticos, como a precipitação, humidade relativa, insolação, temperatura e intensidade do vento durante a antese. A frutificação tem início pelos 15 a 20 anos e, de árvore para árvore, o fruto (glande ou bolota) vai variando de forma e dimensão. Dado o longo período de floração, a maturação da bolota também se alonga no tempo, podendo existir mais do que uma época de frutificação. O que significa que no mesmo ramo podem observar-se frutificações com diferentes graus de maturação. A época de frutificação mais abundante ocorre entre novembro e janeiro (Silva, 2007b).
O sistema radicular é caracterizado por uma raiz aprumada e profunda, que facilita a sua fixação em solos leves e arenosos, que se molda às fendas e espaços disponíveis em solos rochosos. O crescimento radicular inicial é muito superior ao crescimento da parte aérea e as raízes mais grossas a descoberto são revestidas de cortiça (Silva, 2007b).
2. Caracterização das espécies em estudo
14 2.3.1. Características ecológicas
Segundo Silva (2007b), o sobreiro é uma árvore vigorosa que se adapta facilmente a condições adversas de clima e de solo. Em relação ao clima, esta espécie suporta condições de baixa pluviosidade (valor médio anual de 500 mm), sendo que o valor ótimo ronda os 600-800 mm. No entanto, como sucedeu em algumas regiões nacionais, nos anos de 1995 e 2003, as árvores conseguiram sobreviver durante períodos de seca com pluviosidades inferiores a 500 mm. O valor ótimo de temperatura anual para esta espécie situa-se entre 15 e 19ºC, no entanto, Fortes et al. (2006) refere o intervalo de -5 a 40ºC como limites para a sua sobrevivência.
Esta espécie prefere altitudes pouco elevadas, entre 300 e 500 m (Torre, 2006). Contudo, é nas bacias do Tejo e do Sado (altitudes inferiores a 200 m) que se encontram os melhores povoamentos de sobreiro nacionais. Pode ainda vegetar em altitudes superiores, desde que as exposições mais ensolaradas e abrigadas permitam mínimas térmicas absolutas mais elevadas, uma vez que a ocorrência do sobreiro nas nossas regiões montanhosas é condicionada pelas temperaturas mínimas invernais (Silva, 2007b).
Em termos de solos, predomina em solos com valores de pH entre 4,8 e 7,0 e com baixo teor em calcário (Aronson et al., 2009). Exige solos frescos e com alguma humidade em profundidade, mas não encharcados (Torre, 2006). De um modo geral, a espécie é muito tolerante, desenvolve-se bem em solos derivados dos granitos e dos xistos, pobres em matéria orgânica e não compactos (Silva, 2007b).
2.3.2. Distribuição geográfica
O sobreiro encontra as melhores condições de crescimento sob a influência atlântica. Atualmente está circunscrito à região Mediterrânica Ocidental, surgindo de forma natural em Portugal e Espanha, como também no Norte de Marrocos, Argélia e Tunísia. Esta espécie também ocorre numa pequena faixa do Sul de França e na costa ocidental de Itália, incluindo a Sicília, Córsega e Sardenha, como se observa na Figura 6. Portugal é o país que detém a maior área ocupada por esta espécie, sendo que as maiores e mais densas manchas de sobreiro se encontram a Sul do rio Tejo (Silva, 2007b).
Em Portugal, logo a seguir ao eucalipto, o sobreiro é a espécie florestal com maior área, cerca de 737 000 ha correspondentes a 23% da área total de floresta (dados de 2010; ICNF, 2013), na sua maioria em povoamentos de baixa densidade ou em montados (Pereira, 2014).
15
Figura 6 - Área de distribuição da Q. suber com destaque para a distribuição em Portugal. Fonte: Pereira et al.
2. Caracterização das espécies em estudo
17
3. Fenologia
A fenologia define-se como “o estudo da cronometragem de eventos biológicos recorrentes, considerando como causas dessa cronometragem, forças bióticas e abióticas, e a inter-relação entre fases da mesma espécie ou de espécies diferentes” (Lieth, 1974), incluindo eventos como o abrolhamento, a floração, a abertura e queda das folhas (Van Vliet et al., 2003; Cleland et al., 2012) (Figura 7). Contudo, definições mais recentes sublinham que "fenologia inclui o estudo de eventos periódicos influenciados pelo meio ambiente, especialmente mudanças de temperatura provocadas pelo tempo e clima" (Schwartz, 2003).
Figura 7 - Algumas etapas do ciclo fenológico da Q. suber num período de seis semanas.
As alterações na fenologia e as alterações climáticas estão intimamente ligadas (Visser & Both, 2005; Oteros et al., 2013; Richardson et al., 2013). Estas alterações podem variar dentro da comunidade, de indivíduo para indivíduo (Cleland et al., 2007), e de espécie para espécie (Schwartz & Reiter, 2000), determinando a estrutura e o funcionamento dos ecossistemas (Cleland et al., 2007). No entanto, outros autores referem que nem todas as espécies ou populações apresentam alterações fenológicas perante as alterações climáticas (Visser et al., 2003; Both et al., 2004). Nas últimas décadas, a fenologia tem surgido como um foco importante para pesquisas ecológicas (Schwartz, 1999), dada a sua pertinência em abordar questões relacionadas com a capacidade que os organismos possuem em adaptar os seus ciclos de vida como resposta às alterações climáticas (Chmielewski & Rötzer, 2001; Rosenzweig et
al., 2008; Gordo & Sanz, 2009). Uma vez que a fenologia da planta é fortemente controlada
1 2 3
3. Fenologia
18
pelo clima (Gordo & Sanz, 2010) existem inúmeros fatores que a podem modificar, nomeadamente: (1) fatores meteorológicos (luz, fotoperíodo, temperatura, precipitação, humidade, vento, etc.); (2) fatores edáficos (topografia, declive, exposição, etc.); e (3) fatores bióticos (pragas, doenças, competição entre espécies, etc.) (Menzel 2002). De entre os fatores meteorológicos, a temperatura é o que mais contribui para as alterações fenológicas, especialmente nas espécies mediterrânicas (Peñuelas & Filella, 2001; Root et al., 2005; Menzel
et al., 2006; Pau et al., 2011; Diez et al., 2012) pois influencia a duração do tempo de
desenvolvimento das plantas (Partanen et al., 1998; Saxe et al., 2001; Peñuelas et al., 2002; Lu
et al., 2006) por acelerar o abrolhamento, a floração e o alongamento do pedúnculo na
primavera (Vitasse et al., 2009; Richardson et al., 2010). O segundo condicionante mais importante é o fotoperíodo (Morin et al., 2009; Körner & Basler, 2010; Migliavacca et al., 2012) uma vez que controla a indução (formação de gomos no inverno, queda das folhas e resistência ao congelamento), a quebra de dormência, o início do crescimento, eventos reprodutivos e a sincronia da floração (Keller & Körner, 2003; Jackson, 2009). Estes acontecimentos podem originar consequências no ecossistema terrestre (Peñuelas & Filella, 2001; Badeck et al., 2004). Por exemplo, o prolongamento da estação de crescimento afeta a absorção e o armazenamento a longo prazo de CO2 atmosférico pelas florestas caducifólias
(Linderholm, 2006). Por outro lado, pode afetar a sincronização entre herbívoros e/ou polinizadores, trazendo consequências nas interações tróficas e na viabilidade das populações (Visser & Both, 2005; Memmott et al., 2007). Além disso, pode gerar consequências económicas, uma vez que uma alteração no calendário de eventos fenológicos pode afetar o rendimento final de uma cultura (Jones et al., 2005; Porter & Semenov, 2005).
Estudos realizados por Menzel et al. (2006) e Gordo & Sanz (2009) relataram que a abertura das folhas foi antecipada, em média, 2-3 dias por década nas últimas cinco décadas e que a senescência foliar e, consequentemente a sua queda, foi adiada 1-2,5 dias por década. No entanto, estas alterações são mais lentas, mais heterogéneas e menos consistentes do que mudanças na data de abertura das folhas. Além de que, o facto de a fenologia ser extremamente sensível às alterações climáticas, pode, por vezes, trazer implicações no ordenamento do território (agricultura, silvicultura, plantas invasoras, etc.), na saúde humana (transporte de alergénios e vetores de doenças), bem como para numerosos serviços ecossistémicos dos quais a sociedade é dependente (Schröter et al., 2005).
19
4. Parâmetros bioquímicos
4.1. Pigmentos fotossintéticos
A fotossíntese é o processo essencial para toda a vida, produzindo oxigénio e material orgânico (Nelson & Yocum, 2006). Dentro desse processo, o tecido fotossinteticamente ativo das plantas capta energia proveniente da luz solar e armazena-a como energia química na forma de hidratos de carbono (Govindjee & Krogmann, 2004). A cadeia de transferência de eletrões que ocorre nas membranas tilacoides dos cloroplastos, durante a fotossíntese, é altamente dependente de vários pigmentos vegetais, incluindo as clorofilas (pigmentos verdes), os carotenoides (pigmentos amarelos) e as antocianinas (pigmentos vermelhos) (Lichtenthaler, 1987).
As clorofilas a e b são responsáveis por absorver a luz e transferir a sua energia mais tarde (Sims & Gamon, 2002), sendo que a clorofila a é o único tipo de clorofila que se encontra em todos os grupos fotossintéticos das plantas (Campos, 1998). A clorofila foliar controla a quantidade de radiação solar que as folhas conseguem absorver. Assim, o teor em clorofila está diretamente relacionado com o potencial fotossintético da planta e, por conseguinte, com a sua produção primária (Blackburn, 2007). O teor em clorofila também serve como estimativa indireta do estado dos nutrientes, dado que uma parte substancial do azoto das folhas é incorporado na clorofila (Moran et al., 2000). Além disso, o teor em clorofila está intimamente relacionado com o stresse e com a senescência da planta (Hatfield et al., 2008), sendo normalmente condicionado pelas condições de luminosidade e severidade do stresse hídrico e/ou térmico (Pereira, 2000). Na fase de senescência, a clorofila é destruída mais rapidamente do que os carotenoides, modificando a coloração das folhas para tons de laranja, vermelho ou amarelo (Lefsrud et al., 2007). As concentrações relativas aos pigmentos são conhecidas por se alterarem com fatores abióticos como a luz, e assim quantificar essas proporções pode fornecer informações importantes sobre a relação entre as plantas e o ambiente (Richardson et al., 2002). A segunda classe mais importante dos pigmentos é representada pelos carotenoides. Estes são parte das estruturas essenciais da antena fotossintética e do centro de reação. Além disso, desempenham um papel relevante no ciclo da xantofila, importante na proteção das células a partir da foto-oxidação e da foto-inibição (Bartley & Scolnik, 1995) que pode ocorrer se a intensidade de luz for superior à capacidade de transferência de eletrões na cadeia fotossintética. Deste modo, os carotenoides representam, em simultâneo, um papel fotossintético e foto-protetor (Hatfield et al., 2008).
4. Parâmetros bioquímicos
20
Fang et al. (1998) e Richardson et al. (2002) afirmaram que mudanças nos teores das clorofilas e dos carotenoides podem ser ligados à fenologia e a sintomas de stresse. A redução dos teores de clorofila durante o período vegetativo geralmente indica stresse, sendo que o teor de carotenoides providencia informação sobre o estado fisiológico das plantas. Assim sendo, a variação do teor em carotenoides e a proporção de clorofilas é, portanto, vulgarmente utilizada como análise do estado fisiológico das plantas.
4.2. Compostos fenólicos
Como organismos imóveis, as plantas necessitam de produzir um vasto repertório de substâncias com significado adaptativo de forma a sobreviverem nos diferentes nichos ecológicos terrestres (Daayf & Lattanzio, 2008). No seu ambiente natural, as plantas estão expostas à radiação ultravioleta-B do Sol, a qual afeta negativamente não só o ADN, como também as proteínas e as membranas, levando a uma alteração do seu metabolismo. Com o intuito de se protegerem dessa radiação, as plantas sintetizam compostos fenólicos que atuam absorvendo a radiação nas camadas epidérmicas dos tecidos e regulando o sistema antioxidante nas células ou no próprio organismo (Daayf & Lattanzio, 2008). Segundo Dai & Mumper (2010), todas as plantas produzem uma extraordinária diversidade de metabolitos secundários. Os compostos fenólicos são um grupo de metabolitos secundários que se classificam de acordo com as características estruturais e vias biossintéticas (Michalak, 2006). Estes são caracterizados por possuírem, pelo menos, um anel aromático que comporta um ou mais grupos hidroxilo (Naczk & Shahidi, 2006; Harborne, 2009).
Os ácidos hidroxicinâmicos, os flavonóides, as antocianinas e os taninos representam, em conjunto, a maior classe de compostos fenólicos, com aproximadamente 40% do carbono orgânico da biosfera. Embora os compostos fenólicos estruturais, como a lenhina e a suberina compreendam parte desse reservatório de carbono, a incrível variedade de compostos fenólicos não estruturais apresenta diversas funções nas plantas: (1) de defesa (contra herbívoros, micróbios, vírus ou plantas competidoras); (2) de sinalização visual através da pigmentação (atraindo agentes polinizadores ou dispersores de sementes); (3) de proteção da radiação ultravioleta (UV) (Asami et al., 2003; Michalak, 2006); e (4) atuando como antioxidantes (Kähkönen et al., 1999; Ainsworth & Gillespie, 2007). Localizam-se, preferencialmente, nos tecidos mais externos das plantas ou nas paredes celulares que contêm lenhina ou nos vacúolos (Macheix et al., 1990). Os fatores ambientais e genéticos afetam qualitativa e quantitativamente a composição fenólica das folhas (Andreotti et al., 2006). De um modo geral, os compostos
21
fenólicos fornecem informação acerca da capacidade de adaptação das espécies a condições de stresse ambiental (Farah & Donangelo, 2006).
4.3. Açúcares solúveis e amido
As árvores necessitam de conter reservas para garantir a sua sobrevivência durante o inverno e permitir o abrolhamento, para que se possam desenvolver na primavera seguinte (Regier et al., 2010). Os hidratos de carbono não estruturais (açúcares solúveis e amido) são essenciais para a sobrevivência de plantas lenhosas, após um período de stresse ou de dormência (Kaelke & Dawson, 2005) e representam 35% da matéria seca (Nuñez et al., 2010). Além da função de reserva, podem ainda atuar como reguladores osmóticos, protegendo contra o congelamento (Yun et al., 2014), auxiliando na redução das perdas de água (Han et al., 2013), bem como ajudando a mitigar o stresse provocado pelas baixas temperaturas (Xi et al., 2013).
O amido, para além da sacarose, é também um produto primário da fotossíntese e o principal polissacárido de reserva, encontrando-se em todos os órgãos principais das plantas (Ribeiro et al., 2014). É composto por polímeros de glicose, nomeadamente amilose e amilopectina (Orzechowski, 2008). A síntese do amido processa-se quer nas folhas, sob a presença de luz, quer nos órgãos de reserva (sementes e raízes). Nas folhas, que têm capacidade limitada de armazenamento, é sintetizado nos cloroplastos, formando aí reservas a curto prazo. Nas sementes e raízes chega a armazenar-se por alguns meses, e destina-se a constituir reservas a longo prazo que poderão ser mobilizadas na estação seguinte para fornecer carbono e energia durante a germinação e para o crescimento (Campos, 1998).
4.4. Proteínas solúveis
As plantas desenvolveram uma complexa e elaborada rede de sistemas de defesa que interferem na sua interação com o ambiente (Ferreira et al., 2007). Stresses abióticos, como os metais pesados, podem exercer efeitos nas plantas, provocando alterações no teor em proteínas solúveis. O conhecimento da concentração de proteínas solúveis é fundamental para compreender o impacto dos fatores de stresse na proteólise celular e na síntese proteica (Santos & Caldeira, 1999).
Lepeduš et al. (2011) referem uma diminuição do teor em proteínas solúveis em etapas mais avançadas da ontogenia foliar. Bacelar et al. (2009) acrescentaram que, em concentrações mais elevadas, as proteínas podem representar um aumento de enzimas de defesa contra o stresse oxidativo e também uma reserva para a recuperação pós stresse.
4. Parâmetros bioquímicos
23
5. Material e métodos
5.1. Desenho experimental e material vegetal
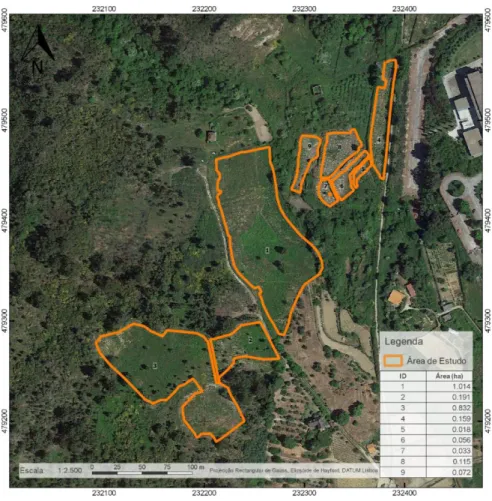
Os trabalhos de campo decorreram em Vila Real, na Universidade de Trás-os-Montes e Alto Douro (Latitude: 41°19'N; Longitude: 7°44'W; Altitude: 450 m), localizada na freguesia de Folhadela. O delineamento experimental utilizado (blocos incompletos) é comum a toda a rede de arboretos REINFFORCE. O arboreto com aproximadamente 3 ha (Figura 8) possui três parcelas onde foram instaladas, em abril de 2012, 35 espécies florestais de interesse económico com três proveniências obrigatórias, sendo cada uma delas avaliada através de 12 plantas.
Figura 8 - Localização das parcelas do arboreto REINFFORCE na UTAD.
O compasso utilizado foi de 3 × 3 m e, aquando da instalação, efetuaram-se adubações de fundo com azoto (N), fósforo (P) e potássio (K). Para proteção das plantas foram colocados tubos protetores de rede da Sinorgan (80 cm de altura e 10 cm de diâmetro), com duas finalidades: evitar possíveis danos causados por animais e permitir a circulação do ar para que as plantas não sofressem sobreaquecimento (Orazio et al., 2013). Para além das espécies
5. Material e métodos
24
instaladas, a vegetação presente no arboreto é constituída na sua maioria por plantas do género
Pteridium e por herbáceas.
O clima é tipicamente temperado mediterrânico com influência continental. Segundo o IPMA (2016; dados de 1981 a 2010) a precipitação média anual da cidade de Vila Real foi de 1128 mm com um máximo de 168 mm registado em dezembro e um mínimo de 12 mm registado em julho. A temperatura média anual foi de 13,4ºC, em que a temperatura média do mês mais quente se verificou em agosto (28, 6ºC) e a do mês mais frio ocorreu em janeiro (2,8ºC). O número de dias de geada por ano é 30, e o principal tipo de solos presente no local são os leptosolos êutricos com pH de 4,5 (Orazio et al., 2013).
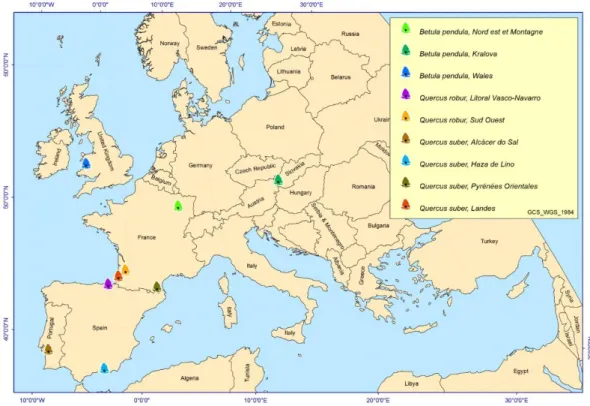
A escolha das espécies teve como critério a taxa de sobrevivência, sendo todas espécies de folhosas (Betula pendula, Quercus robur e Quercus suber), oriundas de nove proveniências, nomeadamente quatro francesas, duas espanholas, uma portuguesa, uma inglesa e uma eslovaca. A B. pendula é representada por três proveniências (Nord, Unit e Kral), a Q. robur por duas (Pago e Sud) e a Q. suber por quatro (Alca, Esp, Pyre e Land). A localização das diferentes proveniências está ilustrada na Figura 9, e na Tabela 1 encontra-se informação mais detalhada sobre a origem das mesmas e alguns elementos climatológicos dos locais de onde a semente foi colhida.
25
Tabela 1 - Identificação das proveniências da B. pendula, Q. robur e Q. suber: código da espécie e proveniência, país da colheita das sementes, região de proveniência,
variáveis geográficas e principais características climáticas. As informações climáticas para cada proveniência foram obtidas no período de 1961-1990 através do software ClimateEU v4.63. Espécie Código da espécie e proveniência País Região de
proveniência Latitude Longitude
Altitude (m) *Temperatura média máxima estival (ºC) *Temperatura média mínima (ºC) *Precipitação total estival (mm) Betula pendula
Bepe-Nord França Nord est et
Montagne 48°23'N 5°58'E 470 16,6 -0,4 362
Bepe-Kral Eslováquia Kralova 48°21'N 17°19'E 240 18,9 -2,3 381
Bepe-Unit Reino
Unido Wales 52°24'N 4° 1'W 15 14,9 4,2 1112
Quercus robur
Quro-Pago Espanha Litoral
Vasco-Navarro 43º12'N 2°25'E 90 20,2 8,9 395
Quro-Sud França Sud Ouest 44°9'N 0°32'W 120 19,7 5,1 378
Quercus suber
Qusu-Alca Portugal Alcácer do Sal 38º22'N 8º30'W 6 23,8 11,6 82
Qusu-Esp Espanha Haza de Lino 36º48'N 3º18'W 1190 22,5 5,2 104
Qusu-Pyre França Pyrénées
Orientales 42º37'N 2º27'E 120 22,4 6,5 305
5. Material e métodos
26 5.2. Parâmetros avaliados
5.2.1. Taxa de sobrevivência
A taxa de sobrevivência (Equação 1) foi avaliada em outubro de 2015 e teve como critério o número de plantas vivas por espécie e por proveniência desde a última observação, ou seja, desde a observação realizada no ano anterior (outubro de 2014). Foram consideradas todas as árvores plantadas originalmente, bem como as plantas retanchadas posteriormente.
Taxa de sobrevivência (%) = nobservado
nplantado × 100 (1)
Em que,
nobservado: número total de plantas vivas registadas desde a última observação
nplantado: número total de plantas instaladas
5.2.2. Crescimento
A avaliação do crescimento teve início em novembro de 2015, isto é, durante o período de repouso vegetativo, onde se registou a altura total (cm) e o diâmetro (mm) de todos os indivíduos (originais e retanchados). As medições em altura foram realizadas com auxílio de dois instrumentos. Nomeadamente, até uma altura de 200 cm as medições foram feitas através de uma vara graduada (Figura 10a), com 1 mm de precisão. Quando a altura ultrapassou os 200 cm utilizou-se o hipsómetro vértex. As medições em altura efetuaram-se desde a base da planta até à folha mais alta. O diâmetro foi medido a 2 cm do solo com recurso a um paquímetro digital (Figura 10b), com 0,01 mm de precisão. Não se efetuaram medições dos diâmetros à altura do peito (130 cm) devido à maioria das árvores não satisfazer esta condição.
27
(a) (b)
Figura 10 - Medições efetuadas em Q. suber: (a) em altura (cm) com recurso a uma vara graduada (b) em
diâmetro (mm) com auxílio de um paquímetro digital.
Foram removidas da análise, árvores cujo crescimento foi visivelmente prejudicado pelos tubos protetores ou por fatores bióticos (pragas e/ou doenças). Deste modo, os crescimentos referentes às variáveis altura (Equação 2) e diâmetro (Equação 3) foram calculados com base nos acréscimos médios anuais (Anexo B).
𝑨. 𝒎. 𝒂𝒉 = ℎ𝑓− ℎ𝑖
𝑎ℎ (2)
Em que:
A.m.ah: acréscimo médio anual para a variável altura, em cm/ano
hi: altura inicial, em cm
hf: altura final, em cm
ah: número de anos decorridos entre a altura inicial e a altura final
𝑨. 𝒎. 𝒂𝒅 =𝑑𝑓− 𝑑𝑖
𝑎𝑑 (3)
Em que:
A.m.ad: acréscimo médio anual para a variável diâmetro, em mm/ano
di: diâmetro inicial, em mm
df: diâmetro final, em mm
5. Material e métodos
28 5.2.3. Fenologia foliar
Em termos de comportamento fenológico, monitorizou-se semanalmente o desenvolvimento dos gomos foliares desde março a meados de junho de 2016. Para esta avaliação foi escolhido e marcado, com uma fita vermelha (Figura 11), o ramo com maior número de gomos foliares em seis das doze árvores por proveniência. Posteriormente os gomos foram observados e fotografados, com uma máquina SONY DSC-HX9V, a fim de registar o seu estádio de desenvolvimento. Para os registos fenológicos, os parceiros do projeto
REINFFORCE consideraram diferentes estádios (Anexo A) representativos da sua evolução,
desde os gomos em dormência até à maturação foliar. Nesta análise, a variável em estudo é o dia do ano em que o gomo selecionado atinge o estádio 19 (maturação foliar). De referir que para a proveniência eslovaca (Kral) da B. pendula, só foram classificados os estádios 09 e 19, uma vez que quando se iniciaram as observações, todos os indivíduos desta proveniência já se encontravam num estádio de desenvolvimento avançado.
Figura 11 - Ramo da Q. suber marcado para avaliação da fenologia.
5.2.4. Análises bioquímicas
5.2.4.1. Recolha e conservação do material vegetal
Para a caracterização da composição bioquímica dos indivíduos foram efetuadas as quantificações de pigmentos fotossintéticos, açúcares solúveis, amido, proteínas solúveis e compostos fenólicos. A colheita do material vegetal realizou-se em maio e de forma aleatória em seis indivíduos de cada proveniência por espécie, onde se recolheram, pela manhã, quatro folhas jovens, desprovidas de ataques de fungos e/ou insetos, tendo o cuidado de utilizar sempre o mesmo critério de recolha, ou seja, na parte superior da planta.


![Figura 3 - Exemplar da Q. robur [1].](https://thumb-eu.123doks.com/thumbv2/123dok_br/15867313.1087574/31.892.213.725.126.567/figura-exemplar-da-q-robur.webp)

![Figura 5 - Exemplar da Q. suber no estado adulto [2].](https://thumb-eu.123doks.com/thumbv2/123dok_br/15867313.1087574/34.892.207.731.714.1069/figura-exemplar-da-q-suber-no-estado-adulto.webp)