iii
Anna Felicity Hobbiss
Research work coordinated by:
Oeiras, 12
thJuly, 2016
Structural scaling and threshold
modulation of dendritic spines driven
by homeostatic plasticity
Dissertation presented to obtain the Ph.D degree in Biology | Neuroscience
Instituto de Tecnologia Química e Biológica António Xavier | Universidade Nova de Lisboav
The work presented in this dissertation was carried out through the International Neuroscience Doctoral Programme (INDP) at the Champalimaud Neuroscience Programme, Champalimaud Centre for the Unknown, under the supervision of Dr Inbal Israely, and the thesis committee supervision of Dr Megan Carey and Dr Marta Moita. Financial support was given by a doctoral fellowship from Fundação para a Ciênica e Tecnologia (SFRH / BD / 51265 / 2010) and Champalimaud Foundation.
vi
Acknowledgements
As with true love, the course of a PhD never did run smooth. The journey along my particular labyrinthine path was made possible only by being joined on it by many people who at one time or another stepped onto the trail to walk with me. Some were there just for a few steps; some trod the whole way next to me. Some appeared with a map (or maybe new GPS co-ordinates) when every vestige of a route seemed to have been lost. Others dropped by to keep me company when I paused to rest. To all, your support, laughter, ideas, inspiration and kindness was what kept me moving when my legs and mind were weary. Or, as a much better writer than I put it, to “Force your heart and nerve and sinew, To serve your turn long after they are gone, And so hold on when there is nothing in you, Except the Will which says to them: ‘Hold on!’”.
Thank you to the Neuronal Structure and Function lab: Inbal Israely, Yazmín Cortez, Ali Özgür Argunşah, Inês Vaz, Maria Royo, Ana Vaz, Catia Feliciano and Daniela Pereira. I feel very lucky to have worked in a group of not just colleagues but close friends. Through travails with the two-photon microscope to heated biological and statistical discussions to over-eating at every lab dinner and a hundred small moments shared in the open lab, it’s been an immense pleasure to be part of this group. Although we had more than our share of technical problems and experimental frustrations, the willingness of everyone to give up their time, expertise, samples, experimental days and chocolate biscuits to help each other as a team was truly inspiring. Inbal, thank you for your guidance, your concern for us, your knowledge and advice on the intimidatingly immense world of synaptic plasticity, and always making sure we had the best birthday cakes of anyone in the CCU. Yazmínita Cortex, for taking us meninos horrorosos under your wing and helping us to become better scientists and better people. Özgürzinho, for being the one always travelling on the path with me, imparting your Matlab expertise and keeping my spirit up with Turkish humour or beer.
vii
Inês, for keeping me company in the microscope room countless times, giving me Portuguese help whenever I needed it and sharing our mutual enthusiasms for cellular biology, home-made sangria, piñatas and mole. Maria, for being a voice of experience and wise advice about science and the PhD, and helping to keep me calm in the face of adversity. Ana, Catia and Daniela, for all the help and support, and making our group such a nice place to work. To my thesis committee, Megan Carey and Marta Moita, for not only being caring and wise advisors, but also transcending that role to become friends.
One of the pleasures of these last 7 years has been spending them with such a warm, diverse and crazy community. The CNP is a unique and special group, which it was my good fortune to be part of from early in its ontogeny to its current mature form. Thanks in particular to the Ar team for helping to create something bigger and more successful than we ever imagined at the beginning and which I take immense pride in; to INDP 09 (definitely the best year group); and to all those who’ve shared conversations (scientific or otherwise), beers, costumes, thanksgiving dinners, jam sessions and much more. There isn’t enough space to name-drop everyone that’s been significant to me but I’ll make to thank you in person. One special shout-out: Libbi, everything was at least twice as fun because of you as it would have been otherwise.
Lastly, to my family – my parents who amongst everything else diligently proof-read this thesis, my siblings, my grandparents, and other family and friends in England and around the world, for the unwavering love, support and confidence; for reminding me there was a world outside my PhD; and for putting up with any long monologues I may have delivered to you on synaptic plasticity and cognition whenever you made the mistake of mentioning memory or the brain.
ix
Table of contents
1 General introduction ... 1
1.1 Learning and plasticity ... 2
1.2 Synaptic plasticity phenomena ... 3
1.3 Homeostatic plasticity theory ... 6
1.4 Homeostasis at the synapse: from theory to experiments ... 12
1.5 Mechanisms of Homeostatic Synaptic Plasticity... 16
1.6 Structural changes accompanying plasticity ... 26
1.7 Interplay between Hebbian and Homeostatic plasticity ... 29
1.8 Outline of work in the thesis ... 31
1.9 Bibliography ... 33
2 Structural correlates of Homeostatic Synaptic Plasticity .... 47
2.1 Abstract ... 49
2.2 Introduction ... 50
2.3 Materials and Methods ... 53
2.4 Results ... 59
2.5 Discussion ... 79
2.6 Bibliography ... 84
3 Threshold modulation of LTP induction by Homeostatic
Synaptic Plasticity ... 89
3.1 Abstract ... 91
3.2 Introduction ... 92
3.3 Materials and Methods ... 95
3.4 Results ... 97 3.5 Discussion ... 118 3.6 Supplementary figures... 124 3.7 Bibliography ... 126
4 General Discussion ... 129
4.1 Abstract ... 1304.2 Overview of work in this thesis... 131
4.3 Beyond synaptic scaling – silent synapses and network function ... 132
4.4 Heterosynaptic plasticity and synaptic clustering ... 134
4.5 Homeostatic Synaptic Plasticity in health and disease ... 137
4.6 The role of sleep in homeostasis ... 140
4.7 Conclusion ... 141
x
Table of figures
Figure 1.1 Homeostatic feedback regulation in the nervous system ... 5
Figure 1.2 Synaptic scaling following chronic activity perturbations ... 8
Figure 2.1 Hippocampal organotypic slice cultures as a study system for Homeostatic Synaptic Plasticity ... 54
Figure 2.2 Acute activity blockade does not cause structural changes ... 60
Figure 2.3 Activity block induces functional Homeostatic Synaptic Plasticity ... 62
Figure 2.4 Structural correlates of Homeostatic Synaptic Plasticity ... 64
Figure 2.5 Distributions of spine morphologies are not affected by HSP at 48 hours ... 66
Figure 2.6 Dendrite thickness and spine density are not affected by activity blockade ... 68
Figure 2.7 Firing rates return to control levels after removal of activity block ... 70
Figure 2.8 Removal of activity block reverses HSP-dependent spine volume increase ... 73
Figure 2.9 Loss of Tsc1 does not impair structural HSP ... 76
Figure 2.10 Chronic GABAA blockade does not induce downscaling through HSP ... 78
Figure 3.1 Protocol for uncaging experiments... 98
Figure 3.2 LTP magnitude and longevity following HSP ... 100
Figure 3.3 Spine growth dynamics during stimulation ... 102
Figure 3.4 LTP induction had increased efficacy following HSP ... 105
Figure 3.5 Small spines show greater LTP after HSP ... 106
Figure 3.6 Behaviour of different sized spines during LTP ... 109
Figure 3.7 Neighbouring spines respond to stimulation after HSP ... 112
Figure 3.8 Reduced threshold for LTP after HSP ... 115
Figure 3.9 Capacity for LTP is maintained after 72 hours of activity block ... 117
Figure 3.10 Post-stimulation growth for real stimulations and sham stimulations ... 124
Figure 3.11 LTP magnitude grouped by spine volume ... 124
Figure 3.12 Neighbouring spine parameters ... 125
xi
Glossary of abbreviations and chemicals
Name/Acronym Complete name Function
APV 2R-amino-5-phosphonovaleric acid NMDA receptor antagonist
ASD Autism Spectrum Disorder
Bicuculline GABAA antagonist
CNQX AMPA receptor antagonist
FMRP Fragile X Mental Retardation Protein
GluA1 Glutamate AMPA receptor 1 Receptor subunit of the AMPA
receptor
GluA2 Glutamate AMPA receptor 2 Receptor subunit of the AMPA
receptor
HSP Homeostatic Synaptic Plasticity
LTD Long Term Depression Synaptic weakening following
activity
LTP Long Term Potentiation Synaptic strengthening following
activity
mEPSC mini Excitatory Post-Synaptic Current Excitatory current caused by
spontaneous pre-synaptic
glutamate release
mTOR Mammalian Target of Rapamycin Signalling molecule fundamental in
the mTOR signalling pathway
NO Nitric oxide Retrograde signal important for
plasticity
PTP Post Tetanic Potential The large increase in potentiation
immediately following a stimulation
PTX Picrotoxin GABAA antagonist
RA all-trans Retinoic Acid
TEA Tetraethylammonium chloride K+ channel blocker
TNFα Tumour Necrosis Factor α
TSC1 Tuberous Sclerosis Complex 1 A repressor of the mTOR pathway
xiii
“Now, here, you see, it takes all the running you can do, to keep in the same place. If you want to get somewhere else, you must run at least twice as fast as that!”
The Red Queen ‘Through the Looking-Glass and What Alice Found There’ by Lewis Carroll
xiv
Resumo
Os circuitos neuronais empregam mecanismos de plasticidade sináptica homeostática (em inglês homeostatic synaptic plasticity, HSP) para manter a atividade neuronal dentro de um intervalo óptimo, contrariando a tendência de desvios na plasticidade do tipo Hebbiano, tanto LTP (long-term potentiation) como LTD (long-term depression). Este mecanismo manifesta-se por uma adaptação dos botões sinápticos, tanto aumentando como diminuindo a atividade neuronal geral. Nesta tese de doutoramento investigamos as estruturas que se correlacionam com a indução de HSP e determinamos como é que esta forma de modulação sináptica tem um impacto na capacidade de troca de informação
durante a plasticidade Hebbiana.Neste estudo demonstramos que o bloqueio
prolongado da atividade de culturas organotípicas de hipocampo causa o crescimento das espinhas dendríticas, complementando o incremento fisiológico da capacidade sináptica. Este crescimento é reversível, voltando aos níveis normais uma vez removido o bloqueio. Utilizando microscopia de dois fotões e uncaging de glutamato, investigamos a fundo como é que sinapses que sofreram adaptação provocada por HSP respondem à plasticidade Hebbiana. Neste estudo descobrimos que 48 horas após o bloqueio da atividade neuronal o LTP é mantido, demonstrando que a capacidade de transmissão de informação no circuito é mantida mesmo após a adaptação. Além disso, verificamos que há um aumento da longevidade da potenciação de espinhas dendríticas individuais, quando comparado com neurónios nos quais a atividade nunca foi restringida, e que o threshold para a indução de plasticidade é menor. Curiosamente, descobrimos que a expressão de LTP é diferencialmente modulada dependendo do tamanho inicial das espinhas dendríticas. Em espinhas pequenas a indução de plasticidade é mais eficaz após HSP, enquanto as espinhas de grandes dimensões são menos influenciadas por este tipo de plasticidade. Estes achados revelam que o mecanismo de modulação induzido durante HSP resulta numa alteração da
xv
distribuição do tamanho das espinhas dendríticas, ocorrendo esta modificação preferencialmente em espinhas de menores dimensões. Estes resultados demonstram que a capacidade do circuito de codificar informação é mantida após a HSP. No entanto, após manipulação da atividade neuronal global, espinhas individuais podem ser moduladas, permitindo que a rede neuronal regule a troca de informação dentro de uma amplitude ótima.
xvi
Abstract
Neural networks employ homeostatic synaptic plasticity (HSP) to maintain activity within an optimal range, countering the tendencies of unchecked Hebbian LTP or LTD to saturate the network. It is manifested by synaptic scaling of all the inputs on a neuron, either upwards to increase global activity or downwards to decrease it. In this thesis we investigate the structural correlates that accompany the induction of homeostatic plasticity and then determine how this form of synaptic modulation impacts the ability of inputs to undergo Hebbian plasticity. We show that prolonged activity blockade in organotypic hippocampal slices causes structural growth of individual dendritic spines, complementing the physiological increase in synaptic strength. This volume increase is reversible, returning to control levels after the activity block is lifted. Using glutamate uncaging and 2-photon imaging, we further investigate how synapses which have undergone synaptic scaling respond to Hebbian plasticity. We find that after 48 hrs of activity blockade, LTP is maintained, showing that the ability for the network to encode information is retained. Furthermore the longevity of potentiation of single spines is increased when compared to neurons in which activity has not been restricted, and the threshold for plasticity induction is decreased. Interestingly, we find that the expression of LTP is differentially modulated depending on the initial size of the spines. In small spines, the induction of plasticity is more efficacious after HSP, whereas larger spines are less impacted by this form of plasticity. After HSP neighbouring spines express short term structural plasticity. Our findings demonstrate that the gain modulation that is induced during HSP results in a shift in the size distribution of dendritic spines, but that such tuning is expressed preferentially at smaller inputs. These results illustrate that after HSP the information-encoding capacity of a network is conserved, but that synapses are individually tuned, allowing the network to regulate the strength of its inputs to function within an optimal range.
1
Chapter 1
1 General introduction
General Introduction:
Homeostatic Synaptic
Plasticity and Nervous system
functioning
2
1.1 Learning and plasticity
Living organisms are defined by two fundamental traits: their genetic make-up, modulated by their environmental experience. One of the essential features of organisms is their ability to sense their environment and respond accordingly, whether it be by locomoting to a chemical stimulus in the case of bacteria, changing patterns of cellular growth to orient towards a light source in flowering plants, or altering gene expression patterns to metabolise a new substrate in yeast. In the animal clade the evolution of the nervous system has made its members uniquely accomplished at responding to stimuli presented by the external world. Animal neurophysiology and the resultant behaviour are effected by a combination of genetic and experience-dependent components, which allow an animal to adapt its behaviour rapidly to better fit the environment. These behavioural changes, known as learning, occur when some aspects of the nervous system are modified, leading it to produce a different output – the learned output – to the same input pattern of activation. The strength of learning is correlated to the valence of the experience. Strongly affective stimuli, either positive such as food consumption, or negative such as pain, will lead to bigger consequent changes in the nervous system. These in turn will make the behaviour which causes the experience more likely or less likely respectively. Nervous systems are extremely well suited to quick and profound changes because their complex multi-layered organisation allows easy modification of their topography and functioning. This permits animals the fast environmental adaption that has made them such a supremely diverse and successful phylum. The ability of nervous systems to change their firing pattern and topography as a response to activity is called neural plasticity. This broad term encompasses a very large range of different mechanisms which can alter the activity of a neural network. The mechanisms vary in timescales (from milliseconds to the lifetime of an animal), in the process of induction and in the mode of implementation in
3
the biological system. This thesis will focus on one of these types of plasticity: the change of strength of synapses (the connections between neurons) known as synaptic plasticity. This mechanism has received a great deal of theoretical and experimental attention since it is considered one of the main biological substrates for memory storage, due to its ability to change circuit functioning and endure throughout an animal’s lifetime. This intense field of study has yielded a huge literature on types of synaptic plasticity and modes of induction, expression and maintenance. In the remainder of the thesis I will discuss two forms of synaptic plasticity which perform fundamentally different roles within the nervous system: Hebbian plasticity and Homeostatic plasticity.
1.2 Synaptic plasticity phenomena
1.2.1 Hebbian plasticity
In 1949, Donald Hebb proposed that if synapses could respond to a particular input pattern by becoming stronger, this could instantiate learning within the network (Hebb, 1949). Alterations of behaviour paralleling synaptic changes were demonstrated in the mid 1960s in Aplysia (Kandel and Tauc, 1965). In 1973 the first experimental evidence was discovered of synaptic strengthening caused by strong activity, and was termed Long Term Potentiation or LTP (Bliss and Lømo, 1973). A few years later, the discovery of Long Term Depression (LTD) as a counterpart to LTP showed that induced changes can be bi-directional (Lynch et al., 1977). LTP and LTD came to be known collectively as Hebbian plasticity, in accordance with Hebb’s postulate. By changing the connection strengths in a network, Hebbian plasticity allows it to encode information, and thus it is regarded as a major mechanism for learning and memory.
4
storage, it requires two important features. Firstly, it must be input specific – that is, the changes to synaptic strength must occur at the same place that the input signal was received, to ensure faithful encoding of the stimulus. Secondly, it must be long-lasting, to be a realistic substrate for memories which can last for a lifetime. These two aims are achieved through precisely calibrated signalling mechanisms, ultimately leading to stable molecular and structural changes at the level of the synapse, dendritic branch and whole cell.
1.2.2 Homeostatic plasticity
Although it is an extremely effective memory storage system, the implementation of Hebbian plasticity also entails a problem, in that it is intrinsically unstable. An increase in synaptic strengths, such as in LTP, would increase the excitatory drive on to a post-synaptic cell, making it more likely to fire. This in turn would increase the likelihood of more LTP; thus positive feedback would quickly saturate the system, resulting in a hyperactive state with saturated synaptic inputs. Conversely, excessive LTD would also proliferate, resulting in a silent state with inputs fully depressed. Modelling studies of Hebbian learning revealed this problem, leading to the need to introduce a stabilising mechanism to keep the activity of the network within a dynamic range (Malsburg, 1973; Miller and MacKay, 1994). Mechanisms which enact this type of compensatory changes would come to be called Homeostatic Plasticity, in accordance with other well established homeostatic mechanisms to control physiological parameters such as body temperature and blood glucose (Cannon, 1932). All of these mechanisms use a negative feedback system, such that if the parameter exceeds the limits of the optimal range, compensatory pathways are activated to push it back to within the correct boundaries. In the specific case of nervous system activity, two distinct but related parameters need to be kept within functioning ranges (Figure 1.1). One is network firing rates, which if left unchecked could reach pathologically high (epileptic) or low (silent) levels. The
5
other is synaptic strengths, because it is a variable range of synaptic weights which gives a network such an immense capacity for information storage. If all synapses become saturated or conversely fully depressed, the information stored in the cell and network will be lost. Conveniently, the regulation of synaptic strengths will also have the benefit of maintaining the network activity levels in check, so both of these parameters can be adjusted by controlling the range of synaptic weights of a neuron.
Figure 1.1 Homeostatic feedback regulation in the nervous system
Activity in the nervous system has an optimal level of activity, whether measured through action potential firing or synaptic strength. If, through changes in input or Hebbian plasticity processes, the level of activity is pushed out of the ideal bounds, the processing of the network and its storage capacity will be compromised. Homeostatic processes use negative feedback mechanisms to restore the system parameters to their ideal intermediate state.
6
1.3 Homeostatic plasticity theory
The first studies to show that loss of function could induce compensatory changes in the nervous system were at the neuromuscular junction. Since the middle of the 20th century, muscles deprived of innervation from motor neurons were known to show increased sensitivity to neurotransmitters such as acetylcholine and to applied current, a phenomenon known as ‘denervation sensitivity’ (Axelsson and Thesleff, 1959). This was described in multiple systems, from the frog skeletal muscle to cat gastrocnemius and iris muscles (Brown, 1937; Nicholls, 1956; Axelsson and Thesleff, 1959). In central mammalian neurons, bi-directional shifts in firing rates of cultured cortical neurons were demonstrated in the early 1990s (Ramakers et al., 1990; Corner and Ramakers, 1991). A long-term blockade of activity caused hyper-excitability once the blockade was released, whilst chronically increased activity lead to decreased firing rates. No mechanism that could account for these changes was proposed however, and the findings were not linked to homeostasis of the network. Instead they were attributed to an altered time course of development. Meanwhile, studies in invertebrate central pattern generators revealed that if rhythmic activity was disturbed by an experimental perturbation, over time compensatory changes could restore the rhythmicity of the circuit (Turrigiano et al., 1994; Thoby-Brisson and Simmers, 1998). In 1994, Miller and Mackay theorised that network stabilisation could be achieved at a synaptic level, a mechanism which I will henceforth refer to as Homeostatic Synaptic Plasticity (HSP) (Miller and MacKay, 1994). The detailed mechanism they proposed was a “non-specific decay of all synaptic strengths, provided the rate of this decay is set for the cell as a whole to cancel the total increase due to specific, Hebbian plasticity”. In essence, rather than any one particular synapse or set of synapses changing weight to compensate for activity changes, the whole synaptic population is affected equally, such that whilst the overall sum synaptic strengths and therefore activity of a neuron is changed, the relative weight
7
differences between synapses is preserved. The advantage of such a system, as opposed to other proposed methods such as a ‘competition rule’ between inputs (Malsburg, 1973), is that differences in weights between synapses are still preserved, so the computational and storage capacity of the network is not compromised and HSP will not erase the information set by Hebbian plasticity.
1.3.1 Synaptic scaling
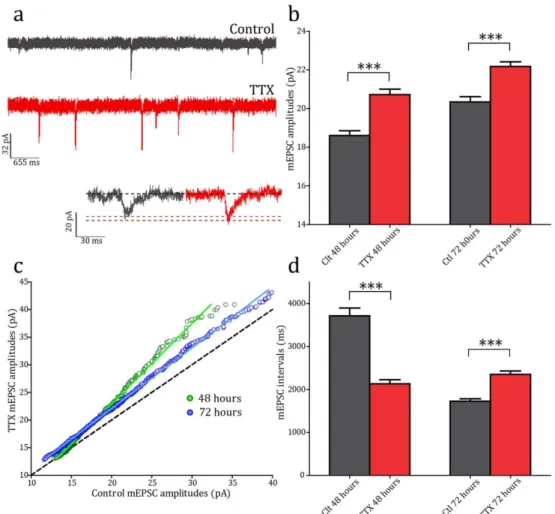
The process outlined by Miller and Mackay can be mathematically expressed as a multiplicative scaling of synaptic strengths (Figure 1.2). The first experimental evidence for such synaptic scaling came from experiments in primary cultures of rat cortical neurons (Turrigiano et al., 1998). Activity was chronically perturbed by application of pharmacological agents for 48 hours, using either Bicuculline (a GABAA receptor antagonist) to increase firing rates by preventing inhibitory transmission, or Tetrodotoxin (TTX) (a voltage-gated Na+ channel antagonist) to block all cell firing. Synaptic strengths were assayed by measuring miniature excitatory post-synaptic currents (mEPSCs) of treated cells. These occur through spontaneous (i.e. action potential-independent) pre-synaptic neurotransmitter release. The size of the post-synaptic current (as measured by whole-cell patch clamp at the soma) is proportional to the strength of the synapse; thus a recording of mEPSCs can be used to sample the synaptic strengths of the recorded neuron. Bicuculline application for 48 hrs led to initially increased activity, and compensatorily decreased mEPSC amplitudes. Conversely, TTX application causing activity blockade led to increased mEPSC amplitudes. In keeping with the mechanism proposed by Miller and MacKay, perturbations of activity caused a linear multiplicative scaling of mEPSC amplitudes, implying that all the synapses had been altered proportionally by the same scaling factor. This and subsequent work supporting work established HSP as a novel form of neural plasticity by which the output of a system can be modulated whilst the
8
information content is preserved (O’Brien et al., 1998; Turrigiano et al., 1998; Desai et al., 1999). Since it requires the integration of network activity, homeostatic plasticity acts at long timescales (on the order of hours to days), and in a global, cell-to-network-wide fashion, as opposed to the fast, single synapse resolution of Hebbian plasticity.
1.3.2 Other types of Homeostatic Plasticity
Since the initial discovery, it has become clear that homeostatic plasticity is a broad class of phenomena acting at different levels and through different mechanisms, a combination of which may be co-opted for use in any one situation (Davis, 2006, 2013; Karmarkar and Buonomano, 2006; Maffei and Turrigiano, 2008; O’Leary and Wyllie, 2011; Turrigiano, 2011). As well as the aforementioned post-synaptic scaling, network stabilisation can be achieved in
Figure 1.2 Synaptic scaling following chronic activity perturbations
Different sizes synapses have a different AMPA receptor content which is set by Hebbian plasticity processes. Chronic changes in activity cause the population of synapses to scale the number of AMPA receptors in the post-synaptic density either down or up, to counteract the shift. The relative numbers of AMPA receptors, and thus the strengths of the synapses, is preserved after the scaling has taken place.
9
numerous other ways (mirroring the abundant mechanisms for instantiating Hebbian plasticity (Nelson and Turrigiano, 2008)). The most prominent of these alternative means of establishing homeostasis are briefly described below. 1.3.2.1 Pre-synaptic homeostatic plasticity
Pre-synaptic changes in LTP and LTD are well established, and have more recently been confirmed as a mechanism of homeostatic plasticity, in systems as diverse as the neuromuscular junction in Drosophila and Xenopus to central nervous system synapses (Davis and Müller, 2014). One interesting question is whether the pre-synaptic changes are triggered by aberrant levels of activity in a cell autonomous way, or whether trans-synaptic signalling from the post-synaptic partner is the requisite signal of activity levels. In fact it is likely that both mechanisms are at play (Jakawich et al., 2010), although there is increasing evidence to suggest that a retrograde signal from the post to the pre-synaptic side is the major determinant controlling aspects of pre-synaptic strength (Petersen et al., 1997; Davis et al., 1998; Nick and Ribera, 2000; Frank et al., 2006; Branco et al., 2008; Lindskog et al., 2010). The identity of the signal is yet to be discovered, although in Drosophila TOR signalling is crucial (Penney et al., 2012) and endostatin has been identified as a possible candidate (Wang et al., 2014). Various different modifications of pre-synaptic function have been observed, including quantal size (i.e. how much neurotransmitter each pre-synaptic vesicle holds) (Davis et al., 1998), speed of vesicle uptake (Thiagarajan et al., 2005), number of docked vesicles (Murthy et al., 2001) and release probability (Sandrock et al., 1997; Davis et al., 1998; Frank et al., 2006; Lindskog et al., 2010; Zhao et al., 2011; Mitra et al., 2012; Wang et al., 2016). The latter mechanism may be caused by changes in calcium influx into the pre-synaptic bouton which promotes vesicle fusion (Frank et al., 2009; Zhao et al., 2011), mediated by plasticity in calcium channel expression levels in pre-synaptic terminals (Jensen et al., 2009).
10 1.3.2.2 Intrinsic threshold of excitability
An alternative to changing the strength of synaptic signalling is to shift the intrinsic excitability of the cell, so that for the same amount of synaptic input the output activity is either increased or decreased (Zhang and Linden, 2003; Turrigiano, 2011). This can be achieved by altering the expression of ion channels in the cell membrane, thus changing the threshold at which a cell will fire. This type of plasticity was observed in early studies on the lobster somatogastric ganglion circuit, where neurons isolated from their normal inputs regain their in vivo bursting patterns after 3 – 4 days in a calcium-dependent manner (Turrigiano et al., 1994). Changes in intrinsic excitability have been observed both in vitro following activity blockade and in vivo after sensory deprivation (Desai et al., 1999; Breton and Stuart, 2009). Various channels have been implicated in changing the membrane conductance. Very clearly important are potassium channels, which are down-regulated in response to chronic activity decrease (Desai et al., 1999; Cudmore et al., 2010; Lee and Chung, 2014; Lee et al., 2015). In addition, the HCN channel, which reduces excitability by increasing the leak current of the membrane, is down-regulated in dendrites in response to sensory deprivation (Breton and Stuart, 2009). Meanwhile, sodium channel conductance is up-regulated after activity blockade (Desai et al., 1999). These changes to intrinsic excitability appear to require calcium signalling (especially for down-shifts after excessive activity), with both L-type calcium channels and NMDA channels being implicated (O’Leary et al., 2010; Lee and Chung, 2014). Various generalised models have shown that if multiple different channels can adjust their levels based on activity, the circuit activity is robust to perturbations (Liu et al., 1998; O’Leary et al., 2014). One interesting thing to note about plasticity of intrinsic excitability is that, although it may take hours to days to develop (in keeping with other types of homeostatic plasticity), the actual switch between different patterns of firing can take less than 1 hr (Turrigiano et al., 1994). This occurs because small changes to conductances can flip neurons
11
between two bistable activity states, one with low levels of firing and one with bursting behaviour. A system can therefore potentially change between firing behaviours very quickly by employing this type of plasticity, allowing for fast adaption to different levels of activity.
1.3.2.3 Excitation-inhibition balance
Network activity can be altered by changing the balance between excitation and inhibition (E/I ratio) in the neural circuit. The E/I ratio has been especially studied in the cortex, where it affects many functions such as gain and feature selectivity (Barth et al., 2004). Its role in homeostatic plasticity was confirmed in
vivo after rats were dark reared for 2 days, resulting in an increased excitatory
and decreased inhibitory drive (Maffei et al., 2004). Successive studies have also demonstrated how changes to the E/I ratio can balance network activity whilst still maintaining cortical function (Atallah and Scanziani, 2009; Pouille et al., 2009; Xue et al., 2014). For example, a neuron receiving inputs with an increased strength has an increased threshold to fire due to correspondingly enhanced feed-forward inhibition (Pouille et al., 2009). This has the effect of stabilising the system, whilst simultaneously making the cortex sensitive to a wide range of input strengths. In the hippocampus, balanced levels of excitation and inhibition produced gamma oscillations which are critical for its function (Atallah and Scanziani, 2009). Indeed, individual cells themselves receive balanced amounts of excitation and inhibition, stabilising output in a cell-autonomous manner (Xue et al., 2014). One candidate for modulating the relative strengths of excitation and inhibition is nitric oxide (NO), which is known to play fundamental roles in Hebbian pre-synaptic plasticity, and also in homeostasis for both excitatory and inhibitory cells (reviewed in (Hardingham et al., 2013)); as well as other signalling molecules such as Npas4 (Spiegel et al., 2014).
In addition to changes to the E/I balance, homeostatic changes to inhibitory activity alone have become the focus of increasing interest (Wenner, 2011).
12
Classic studies on visual deprivation documented that the density of GABAergic cells was reduced following the loss of visual input or activity deprivation (Hendry and Jones, 1986; Benevento et al., 1995; Rutherford et al., 1997, 1998). Detailed studies into mechanisms have revealed that inhibitory synapses show many of the same homeostatic mechanisms when exposed to activity perturbations as excitatory synapses, including bidirectional shifts in activity (Karmarkar and Buonomano, 2006), synaptic scaling (Kilman et al., 2002; Holopainen and Lauren, 2003; Chang et al., 2010) and pre-synaptic changes (Hartman et al., 2006). This being said the relationship between activity shifts and inhibition is not a simple one, since visual deprivation during the critical period can also cause potentiation of inhibition rather than the expected depression (Maffei et al., 2006; Nahmani and Turrigiano, 2014). These results suggest that the timing of the activity shift is crucial for determining how the system will respond.
1.4 Homeostasis at the synapse: from theory to experiments
1.4.1 Computational modelling of Homeostatic Synaptic Plasticity
The homeostatic synaptic plasticity field has a rich history of combining theoretical predictions and experimental findings. Indeed modelling studies were quick to reveal the intrinsic instability of Hebbian processes, leading to the first proposals of homeostatic mechanisms. Initial papers exploring the computational dynamics of LTP and LTD had to compensate for the positive feedback inherent in Hebbian theory, for instance by providing constraints to the expressed plasticity (Oja, 1982), by setting an overall constant sum synaptic strength thus forcing competition for plasticity between inputs (Malsburg, 1973), or by using a sliding threshold of plasticity to balance LTP and LTD as seen in the seminal BCM rule of Hebbian plasticity (Bienenstock et al., 1982).
13
Initially these stabilising mechanisms were built into the Hebbian models as a fundamental aspect of the plasticity. Opposing this, a train of thought proposing that in fact Hebbian and homeostatic plasticity were separable processes began to arise. As was referred to earlier in this chapter, the first explored mechanism cell-wide synaptic strength changes proposed that a multiplicative decay (as opposed to a linear decay) would preserve information content in the network whilst constraining the output activity (Miller and MacKay, 1994).
Since these beginnings, the computational theory of homeostatic plasticity has expanded hugely, with the development of models which reflect realistic physiological parameters for synaptic strength changes. HSP has been shown influence Hebbian plasticity (Litwin-Kumar and Doiron, 2014), STDP (Clopath et al., 2010), connectivity (Tetzlaff et al., 2011) and working memory (Renart et al., 2003). Interestingly, some discrepancies between theory and experimental data have arisen. In particular, the timescale over which homeostasis occurs is still under debate. A recent study formulated a model where homeostasis needs to have a fast rate detector (on the order of seconds to minutes) to stabilise Hebbian plasticity (Zenke et al., 2013). Experimentally however, HSP is induced over hours to days (see parts 1.5.1.1 and 1.5.1.2 for more details). Very recently a fast form of pre-synaptic homeostasis was reported which could fulfil the conditions required (Wang et al., 2016). Future work theoretical and experimental is needed to resolve how the different timescales interact in dynamic systems.
1.4.2 The spatial expression and scale of HSP
Homeostatic synaptic plasticity has been shown to occur widely across the nervous system. The majority of studies have been conducted in cultured cells, from the cortex (Turrigiano et al., 1998), hippocampus (Lissin et al., 1998), peripheral neurons such as spinal cord (O’Brien et al., 1998), and somatogastric ganglion cells (Turrigiano et al., 1994). In vivo homeostatic synaptic plasticity in
14
various systems has also been demonstrated, although fewer studies exist due to the increased difficulty of the experimental protocol. The majority of studies are from sensory cortices, where activity reduction can be achieved through sensory deprivation protocols such as ocular deprivation (Desai et al., 2002; Maffei et al., 2004; Goel and Lee, 2007; Maffei and Turrigiano, 2008; Keck et al., 2013) or whisker trimming (Greenhill et al., 2015). Other sub-cortical areas such as the hippocampus (Pawlak et al., 2005; Echegoyen et al., 2007) and superior colliculus (Chandrasekaran et al., 2007), and the spinal cord (Knogler et al., 2010; Garcia-Bereguiain et al., 2013) have also been used to demonstrate homeostatic synaptic responses. The details of in vivo homeostasis and its implications for nervous system functioning are discussed later in this chapter in Section 1.5.2.
A complicating factor for the study of homeostasis is that although the basic processes employed seem to be universal, different synapses show different responses to shifts in activity. Even within single structures such as the hippocampus, different synaptic sites (dentate to CA3 and CA3 to CA1) show differing or even opposing responses (Kim and Tsien, 2008; Lee et al., 2013). Compounding the somewhat bewildering array of modes of synaptic homeostasis is the question of the spatial scale on which the homeostatic signal acts. Processes such as regulation of the excitation/inhibition balance are by definition network-level phenomena, but synaptic processes can be the result of much smaller-scale integrations of activity. Synaptic homeostasis is normally described and modelled as a cell-wide process, whereby neuron activity is integrated over long time-periods and all spines are then scaled in concert. Theoretically, this type of cell-wide scaling differentiates HSP from Hebbian plasticity and so the information encoded in synaptic weights is not lost (Toyoizumi et al., 2014). Although there is a weight of experimental evidence to support wide-scale scaling, most of the induction protocols involve a global up or down-regulation of activity, so it is not possible to differentiate between global
15
or local plasticity processes. In fact, studies looking at smaller neuronal segments, from individual dendrites (Sutton et al., 2006; Branco et al., 2008) down to single spines (Hou et al., 2008, 2011; Béïque et al., 2011), have indeed seen evidence for homeostasis occurring at these levels, in both the upward and downward directions. This spine-specific scaling was induced by manipulating the activity levels of upstream cells, and then examining the post-synaptic sites they connected to. Its expression involved changes in AMPAR content in the spines, similarly to global induction protocols (Hou et al., 2008, 2011). Neighbouring spines receiving normal levels of input remained unchanged, suggesting that homeostatic regulation was local to the individual synapse. In the visual cortex in vivo however, selective synapse silencing produced a potentiation of all synapses including those not affected by the deprivation protocol, suggesting that a more global homeostatic effect occurs (Greenhill et al., 2015). The potential for synapse-specific homeostasis presents an interesting conundrum for how HSP and Hebbian plasticity can co-exist, since the induction of one type would seem to be antagonistic to the other. In Hebbian plasticity, reduced signalling (i.e. low levels of calcium in the spine) leads to depression of the spine, but in homeostasis reduced activity would lead to potentiation. It will be intriguing in the future to see whether more evidence for synapse-specific HSP emerges.
Interestingly, there is also disagreement in the literature about whether post-synaptic homeostasis is induced by changes in pre-post-synaptic activity (i.e. the synapse’s own glutamatergic drive), or post-synaptic activity (implying some integration of the activity of the post-synaptic cell). Two studies saw a decoupling of post-synaptic spiking activity with synaptic scaling, leading to the conclusion that neurotransmission is the crucial induction signal for scaling (Hou et al., 2008; Fong et al., 2015). Other studies (Burrone et al., 2002; Ibata et al., 2008; Goold and Nicoll, 2010) have demonstrated that indeed activity changes in only the post-synaptic cell do change the input synaptic strength. It is likely
16
therefore that both pre- and post-synaptic changes have the capacity to induce homeostasis.
1.5 Mechanisms of Homeostatic Synaptic Plasticity
1.5.1 Molecular pathways of HSP
Following the discovery of HSP an intense area of research has been to uncover the molecular pathways involved. Unsurprisingly given the diversity of homeostatic phenomena, many different molecules have been seen to play a role in HSP. Various overlapping pathways have been identified, which depend on the protocol used to induce HSP. Below I will discuss some of the different molecular signatures of these alternate HSP forms.
1.5.1.1 Up-scaling induced by reduction in excitatory drive
The original synaptic scaling discovery used TTX, a sodium channel blocker, to prevent spiking activity in cultured neurons (Turrigiano et al., 1998). This has the effect of both blocking evoked neurotransmission (i.e. reducing excitatory drive) and also post-synaptic spiking. Notably however, miniature transmission (i.e. spontaneous vesicle fusion) still occurs under these conditions, so in contrast to some of the protocols described further on in this chapter, there is not a total absence of AMPA and NMDA activation. This difference is important mechanistically to determine the time-course and molecular identity of the scaling (Sutton et al., 2006). Sensory deprivation in vivo, which again has the result of reducing excitatory drive without pharmacologically blocking any of the receptors, should also induce this same type of HSP.
The synaptic strength changes of all different types of HSP appear to be mediated by changes in the AMPA receptor content at the synapse. Activity reduction protocols result in an increased AMPAR content causing functional up-scaling,
17
whilst increased activity leads to down-scaling due to less synaptic AMPAR. The AMPAR subunit required for the expression of the scaling differs depending on the induction protocol. There are 4 AMPAR subunit types; GluA1, GluA2 and GluA3, and GluA4, which have different functional properties. The full AMPA receptor is tetrameric, normally consisting of dimers of dimers which commonly combine GluA2 with either GluA1, 3 or 4 (Isaac et al., 2007). GluA2-lacking AMPARs have an important functional role to play in synaptic plasticity since they are calcium permeable, so will enhance calcium-dependent signalling pathways in synapses. GluA2-lacking AMPARs are thus known to be involved in the early stages of LTP, whereas GluA2-containing receptors are inserted at later stages during what seems to be a consolidation process.
When excitatory drive is reduced using either TTX or sensory deprivation, both the GluA1 and GluA2 subunits are up-regulated at synapses in a correlated fashion (Wierenga et al., 2005). It is GluA2 however that appears be to the crucial component, since knock down of GluA2 but not GluA1 abolishes scaling (Gainey et al., 2009). GluA2 is also inserted on a surprisingly fast timescale, with changes being visible after only 4 hours of activity block (despite most protocols of scaling showing changes only at the 24 hour timepoint) (Ibata et al., 2008). GluA1 may also have a role to play in the functional expression of scaling however, since phosphorylation of this subunit through PKA can cause up-scaling (Diering et al., 2014). Similarly single-spine induction protocols also appear to up-regulate, and require, GluA2-lacking AMPA receptors (identifiable through pharmacological or electrophysiological methods) (Hou et al., 2008; Béïque et al., 2011), as do some developmental homeostasis processes in vivo (Garcia-Bereguiain et al., 2013). Further on in this chapter I will explore the important role that GluA2-lacking receptors play in a different type of synaptic homeostasis, a fast retinoic acid-dependent form (see section 1.5.1.2).
18
expression of scaling, a lot of work remains to understand the molecular mechanisms involved. An increasingly complex set of pathways is emerging to span the gap between the start point of sensing of activity changes, to the insertion or removal of the AMPARs at the other end. As with other types of plasticity, calcium is looking increasingly promising as a candidate for initiating the plastic changes (Liu et al., 1998). In particular, a drop in Ca2+ levels at the soma can induce synaptic scaling (Ibata et al., 2008); a mechanism which makes intuitive sense for the detection of cell-wide activity (as opposed to local dendritic calcium changes which initiate Hebbian plasticity). The change in Ca2+ concentration appears to be sensed by CaMKIV, one of the calcium/calmodulin-dependent protein kinase (CaMK) family (Ibata et al., 2008). CaMKs play many important signalling roles in neurons, especially in plasticity through their role in calcium sensing. Another member of the family, CaMKII, is an extremely important and abundant signalling molecule in Hebbian synaptic plasticity. Two of the subunits which compose the complete CaMKII molecule, CaMKIIα and CaMKIIβ, change in proportion in response to activity blockade with TTX (Thiagarajan et al., 2002), with β levels rising whilst α is down-regulated (this shift is reversed after chronic increase of activity with bicuculline). β subunits have a higher sensitivity to calcium so are activated by lower calcium levels. This shift will therefore act to sensitise the quietened system to any activity. CaMKII in turn regulates the recruitment of GKAP, a post-synaptic scaffolding protein which is necessary for synaptic scaling (Shin et al., 2012).
Biology is a frugal system, and in HSP, as in multiple other physiological processes, molecules have been co-opted from other purposes to form part of these signalling pathways. For example, Tumour Necrosis Factor α (TNFα), a pro-inflammatory cytokine, is crucial for the synaptic scaling process, both in
vitro (Stellwagen and Malenka, 2006) and in vivo (Kaneko et al., 2008; Steinmetz
and Turrigiano, 2010), where it acts to maintain the synapses in an up-scaled state (Steinmetz and Turrigiano, 2010). Importantly, it is not required for LTP or
19
LTD (Stellwagen and Malenka, 2006; Kaneko et al., 2008; Knogler et al., 2010) – a crucial clue that at the molecular level these different types of plasticity can be differentiated. Adding another component to the HSP signalling circuit, TNFα is derived from glia surrounding the neurons undergoing scaling (Stellwagen and Malenka, 2006), through their sensitivity to glutamate signalling. It is possible that HSP can co-opt glial processes more easily that Hebbian plasticity since its induction usually involves a wide-scale activity change (rather than activation at single inputs). Indeed, glial and cell adhesion molecules are emerging as important players in the HSP process (Thalhammer and Cingolani, 2014). Towards the end of the scaling pathway, stargazin, an AMPA auxiliary subunit, has been shown to play a crucial role in AMPAR trafficking in the synapse upon induction of HSP (Louros et al., 2014). Stargazin is known to regulate the delivery of AMPARs to the synapse. It is up-regulated upon activity blockade, whereupon it is phosphorylated to promote scaling.
Alongside these major signalling players, numerous other molecules have been shown to influence scaling, including BDNF (Rutherford et al., 1998), MSK1 (Corrêa et al., 2012), Beta-3 integrins (Cingolani et al., 2008), STEP61 (Jang et al., 2015) and MHC1 (Goddard et al., 2007). It is clear that, as for Hebbian plasticity, HSP is an extremely complex process which requires many layers of control for its successful expression.
1.5.1.2 Retinoic acid dependent up-scaling
In 1998, just subsequent to the first description of HSP (Turrigiano et al., 1998), changes in the accumulation of AMPA receptors following chronic increases or decreases of synaptic signalling were described (O’Brien et al., 1998). When APV, an NMDA receptor antagonist, was applied in conjunction with an activity blocker for 72 hrs to eliminate synaptic signals, AMPA receptor levels increased in the synapses. Subsequent work using this protocol of NMDA receptor
20
blockade elucidated that this form of HSP is mechanistically different to the one described in the previous section (Ju et al., 2004; Sutton et al., 2004, 2006; Aoto et al., 2008; Sarti et al., 2012). The reason for this is that not only is activity-dependent glutamate release inhibited, but also the activation of post-synaptic NMDA receptors by spontaneous glutamate release (miniature synaptic transmission) is prevented. The loss of miniature signalling causes an up-regulation of translation in the dendrite, accompanied by up-scaling of synaptic strengths, which can be observed only an hour after application of the combined drugs (Sutton et al., 2004, 2006). In contrast, synaptic scaling caused by only action potential blockade can take 24 hours or more (Sutton et al., 2006). This fast form of HSP has been found to rely on a newly discovered function for
all-trans retinoic acid (RA), a signalling molecule previously known for its role as a
morphogen during development (Aoto et al., 2008; Chen et al., 2014). Two hours of RA application induces synaptic scaling, and scaling caused by TTX + APV is blocked by inhibitors of RA signalling (Aoto et al., 2008; Sarti et al., 2012). These two pathways – RA-mediated and non-RA mediated – remain mechanistically distinct even at the timescales where both are evident, i.e. at more than 24 hours of treatment, at which time TTX-mediated synaptic scaling is not affected by RA inhibition, but TTX + APV scaling is abolished (Soden and Chen, 2010; Wang et al., 2011).
Blockade of NMDA receptors inhibits Ca2+ entry thus lowering Ca2+ levels in the dendrite, which has been found to be a crucial step for the up-regulation of RA synthesis (Pawlak et al., 2005; Wang et al., 2011). This difference in Ca2+ signalling may explain why TTX + APV co-ops RA-mediated HSP, whilst TTX alone does not. As mentioned in the previous section, Ca2+ signalling in the soma, rather than the dendrites, was important for TTX-induced HSP (Ibata et al., 2008). Calcium concentration is sensed by calciuneurin, a Ca2+-dependent phosphotase, which then promotes RA synthesis (Arendt et al., 2015). In addition to calcium sensing, RA-synthesis and GluA1 synthesis also happen locally at
21
dendrites (Ju et al., 2004; Maghsoodi et al., 2008). This could give a vital insight into the different spatial scales at which HSP is seen in the literature. Non-RA mediated HSP, which is sensed at a cell-wide level, could co-opt scaling at all synapses simultaneously, whereas RA-mediated, which senses activity and initiates signalling mechanisms at a local level, could act on much more restricted spatial scales.
I have referred to this type of HSP as the ‘RA-dependent form’, and differentiated it from the previously described non-RA dependent form. It is probable that it is not only the RA component, but the whole signalling cascade which differs between these two types. Much less is known however about the other components. One better explored aspect involves protein synthesis. The scaling response requires both eEF2 (Sutton et al., 2007), and FMRP (Soden and Chen, 2010) which are both involved in the control of translation in dendrites. The requirement for protein translation and transcription will be explored more fully in section 1.5.2 but the de novo synthesis of proteins is an important part of scaling expression.
As with the previously described forms of HSP, RA-dependent scaling is effected by AMPAR insertion into the synapse. Whereas GluA2 seems to be the major subunit player in non-RA dependent HSP, the RA-dependent form is instantiated by GluA2-lacking receptors, likely GluA1 homomers (Ju et al., 2004; Thiagarajan et al., 2005; Shepherd et al., 2006; Sutton et al., 2006). These receptors are dendritically synthesised which may account for the fast dynamics of this type of scaling, since the receptors do not have to be newly transcribed and transported from the soma (Ju et al., 2004). Some recent work has shed light on how the synthesis of AMPAR subunits is controlled through interactions with micro-RNAs (miRNAs). These short lengths of RNA have received great attention in recent years for their regulatory roles in mRNA translation. Two different miRNAs have been identified which modulate AMPAR translation in opposing directions
22
during synaptic scaling – miR-92a which suppresses GluA1 synthesis and is down-regulated upon activity blockade (Letellier et al., 2014), and miR124 which suppresses GluA2 synthesis during activity blockade, leading to the formation of GluA2-lacking AMPARs (Hou et al., 2015).
1.5.1.3 Down-scaling
HSP would be of limited use if it could only implement scaling in one direction. Indeed, scaling downwards in response to over-activity is also known in the literature (O’Brien et al., 1998; Turrigiano et al., 1998; Goold and Nicoll, 2010; Hou et al., 2011). It is worth noting however that there are many fewer studies than for up-scaling, and the vast majority of these are in primary cultured neurons, instead of more the physiologically relevant systems of slices or in vivo. This may be partly due to the increased experimental challenges of chronically increasing activity (though see (Corrêa et al., 2012) for the use of environmental enrichment as an activity increase paradigm), but it could also represent that the system is not symmetric with regards to the methods used to implement up-scaling versus down-up-scaling, and so down-up-scaling is harder to observe.
That down-scaling is a qualitatively different process to up-scaling is shown by the different pathways involved. For instance, CaMKII, fundamental for the expression of up-scaling, is not required for down-scaling (Goold and Nicoll, 2010; Hou et al., 2011). A change in calcium level is still the most likely means by which activity levels are sensed, as shown by the necessity of various neuronal components which control intra-cellular calcium, including NMDA receptors and voltage-gated calcium channels (Goold and Nicoll, 2010; Hou et al., 2011). Other calcium-binding molecules must therefore take the calcium-sensing role that CaMKII played in up-scaling. Indeed, different members of the CaMK signalling cascade, most prominently CaMKK (which activates CaMKIV and CaMKI), are necessary for the expression of down-scaling (Goold and Nicoll, 2010).
23
Once over-activity has been sensed, a process to decrease synaptic strength must be enacted. Here the immediate early gene Arc plays an important role (Bateup et al., 2013a; Korb et al., 2013). As well as acting as a marker for recent activity, Arc is known to play a role in LTD by stimulating the endocytosis of glutamate receptors (Bramham et al., 2008). It is unsurprising that it also proves essential for the weakening of synapses in response to chronic over-activity, since AMPA removal is responsible for the functional post-synaptic change after downwards HSP (O’Brien et al., 1998). Arc is up-regulated after increased activity (Bateup et al., 2013b), and also acts to down-regulate transcription of GluA1 (Korb et al., 2013). Arc’s well-documented fast induction after synaptic activity (for which it has become known as an immediate-early gene) may have another consequence; that down-scaling is induced at shorter timescales than up-scaling. Using optogenetic methods to increase activity of single cells, the removal of GluA1 receptors in spines was seen to take place as early as 30 minutes into the light treatment (Hou et al., 2011). Other studies report strong effects after only a few hours of treatment (Bateup et al., 2013a, 2013b). Surprisingly however, Arc KO neurons are still capable of synaptic down-scaling, possibly implying that a number of parallel redundant pathways can achieve this result (Shepherd et al., 2006).
Finally, a host of other molecular players have also been shown to be necessary for synaptic down-scaling, including PP1 I-2 (Siddoway et al., 2013), MeCP2 (Qiu et al., 2012; Zhong et al., 2012) and components of the Plk pathway and CDK (Seeburg and Sheng, 2008; Seeburg et al., 2008) (for a full review of molecular pathways see (Siddoway et al., 2014)).
1.5.2 The involvement of proteins in HSP
Proteins are fundamental building blocks of cellular processes, and plasticity is no exception. As with Hebbian plasticity, de novo protein synthesis seems to be necessary to maintain HSP for extended periods of time. Synthesis of GluA1
24
occurs locally in dendrites in response to RA-mediated HSP (Ju et al., 2004; Sutton et al., 2004, 2006). FMRP, an important player in controlling levels of protein synthesis, is required for this process (Soden and Chen, 2010). Non-RA mediated HSP however requires not just translation but also transcription – reinforcing the theory already expounded in this introduction that the non-RA form is a global cell wide process, whereas the RA-form is more local (Ibata et al., 2008). Similarly, global downscaling required not just translation but also transcription (Goold and Nicoll, 2010). These findings reinforce the fundamental role new proteins play in the expression and maintenance of HSP.
1.5.3 HSP in vivo
The majority of work on HSP has been conducted in culture systems, either in primary neuron cultures (Turrigiano et al., 1998) or in organotypic slices (Karmarkar and Buonomano, 2006). This is due to the increased tractability of the in vitro system, where chemical manipulations, and their molecular and physiological effects, are much more easily controlled and assessed than in the live animal. In vitro systems have allowed huge strides to be made in understanding how HSP works using these systems. An increasing number of studies however have now addressed the question of how HSP is expressed in intact nervous systems.
Many of the in vitro studies on the mechanisms and expression of HSP have been borne out through in vivo studies. Multiplicative synaptic scaling has been seen in various sensory cortices after deprivation, including visual (Desai et al., 2002; Goel and Lee, 2007; Keck et al., 2013) and somatosensory (Greenhill et al., 2015), as well as in other regions such as the spinal cord (Knogler et al., 2010; Garcia-Bereguiain et al., 2013). As a counter example, synaptic scaling was absent after
25
One potentially confounding factor for drawing a universal conclusion about HSP
in vivo is that the type and extent expressed is highly influenced by
developmental age (Desai et al., 2002; Knogler et al., 2010). In sensory cortices, as well as other parts of the brain, the neurons progress though periods with differing capacities of plasticity expression. The peak, normally during infancy or childhood, is known as the critical period, and is the time when much of the framework for the sensory processing is established. Without activity during this period, the neurons can never recover their optimal functioning into adulthood (Wiesel and Hubel, 1963). This inability to recover from deprivation suggests that homeostasis is suppressed in some areas during the critical period – possibly because compensatory mechanisms could hinder the development of acuity. Indeed at the retinocollicular synapse, mice without normal retinal waves throughout development (due to deletion of a critical protein) have larger receptive fields, compensating for the lower peak responses (Chandrasekaran et al., 2007). In the visual cortex, studies from the Turrigiano lab found that homeostasis in layer 4 visual cortical neurons was prominent in early life (Desai et al., 2002; Maffei et al., 2004) but was switched off by P21 (Maffei et al., 2006), indicating that by the beginning of the visual critical period homeostasis is suppressed. In upper layers of the cortex however, plasticity remains prominent through the critical period (Maffei and Turrigiano, 2008) and into adulthood (Goel and Lee, 2007). An interesting study which examined the length of time needed to induce homeostatic responses at different developmental ages in the hippocampus found that in early life only 15 hours of activity shift was needed, but later in development more than 40 hours was required for HSP induction (Huupponen et al., 2007). These differences may reflect that fact that throughout development, the subunit composition of receptors is known to change, altering the functional properties of the circuit. One example is the NMDA receptor, a cation channel which acts as a coincidence detector for pre- and post- synaptic activity and thus is vital for circuit refinement and Hebbian plasticity. This
26
receptor has a developmental switch, from being enriched in the subunit GluN2B, which is highly calcium permeable and has a prolonged channel opening, to GluN2A, which with its decreased calcium permeability reduces the capacity for plasticity (Paoletti et al., 2013). These differences in synaptic receptors may make different types of homeostasis more or less likely, depending on the developmental stage they manifest. What is clear is that HSP, and indeed other complementary mechanisms of homeostasis, exist in a complex interplay throughout different periods of the lifetime of an animal.
1.6 Structural changes accompanying plasticity
As is detailed above, much has been elucidated regarding the molecular pathways and functional expression of HSP. An area which is still to be explored is the structural consequences of HSP for the dendritic spines and the neuron. The structure of a neuron – its physical size and shape – is strongly influential with regards to its function. The high degree of specialisation seen in different neuron subtypes (consider the extreme differences between pyramidal cells, Purkinje cells and motor neurons), shows how precisely adapted different neuron types are for their roles. Although there is undoubtedly a large genetic component, structural parameters such as dendrite branching, spine number or axon diameter are not fixed but can undergo activity and time-dependent changes throughout development and into adulthood.
In this thesis I will focus on structural changes to the dendritic arbour, since there is a wider scope for plasticity in the dendrites than the axon, and the dendrite and spines provide a visual read-out to predict downstream responses to activity. There is some evidence for structural changes to the axon upon homeostatic plasticity induction but this is outside the scope of this work (Yamahachi et al., 2009). Dendritic structural changes strongly influence the
27
function of the cell by changing the electrical and biochemical properties of its inputs, and thus its firing pattern. A simplistic example is that if a cell develops more dendritic spines it has increased input and thus will be more likely to fire (although this can of course be counterbalanced by other type of modifications such as threshold changes). A more subtle example occurs at the level of a single spine, where by increasing the spine volume there is more room for both receptors (such as AMPARs) and also organelles such as endoplasmic reticulum, which could enhance signalling at the synapse and so strengthen it. Hebbian plasticity is known to cause structural changes, for both LTP (Matsuzaki et al., 2004) and LTD (Ramiro-Cortés and Israely, 2013). Much less is known about structural correlates with regards to homeostatic plasticity. In particular, it remains to be discovered whether indeed structural changes are a hallmark of HSP, and if so whether they mirror the changes seen in Hebbian plasticity or whether they employ parallel mechanisms.
Much of the evidence for structural changes following HSP comes not from a mammalian system but from Drosophila, a classic model for homeostasis studies. Activity blockade throughout development causes large structural re-organisations, including increases in dendritic arbour size and complexity (Tripodi et al., 2008) and a larger synaptic glomerulus (Kremer et al., 2010).
Drosophila synapses are not located on dendritic spines however, so the
adaptations seen in may be different to those in a chordate system. In mammalian neurons, there is ample evidence that the number of synaptic connections changes after HSP. Here the developmental stage of activity deprivation determines the structural adaptations, with decreased synapses if deprivation occurs before synaptogenesis, but increased synapses if it occurs after (Stroemer et al., 1995; Burrone et al., 2002; Kirov et al., 2004; Zuo et al., 2005a; Arendt et al., 2013). Complementing this, synaptic elimination follows chronic increases in activity (Goold and Nicoll, 2010). However, see (Wallace and Bear, 2004; Thiagarajan et al., 2005) for experiments which result in unchanged
28
or decreased spine density following reduced activity. Interestingly, spine turnover can still occur even throughout chronic blockade of activity (Yasumatsu et al., 2008).
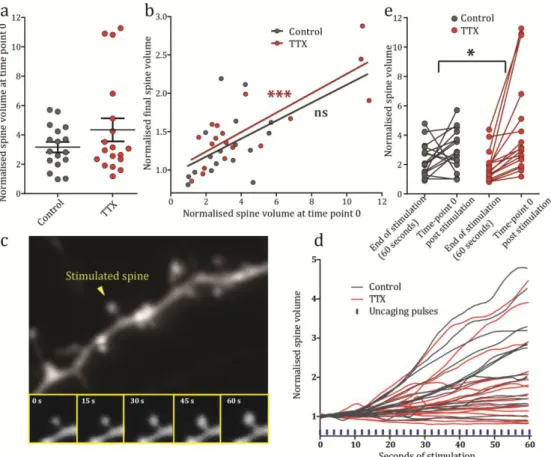
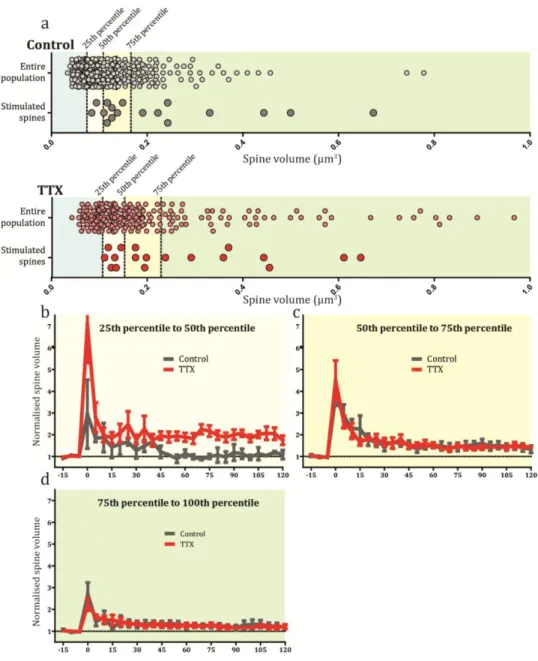
Not only spine number but also the structure of spines themselves can change with shifts in activity, although there is conflicting evidence over the dynamics of this process. Sensory deprivation in vivo and chemical blockade of activity have been reported to cause spine volume increases, accompanied by corresponding increases in the size of the pre-synaptic terminals (Murthy et al., 2001; Wallace and Bear, 2004; Keck et al., 2013). This affect was seen to be reversible after re-exposure to activity (Wallace and Bear, 2004). On the other hand, various studies have reported no change to spine volume following chemical manipulations or single synapses activity loss (Yasumatsu et al., 2008; Béïque et al., 2011). Presumably, these differences relate to the induction protocols used in the specific experiments.
The individual morphology of the spines also influences function. Spines are traditionally divided into 4 different classes; mushroom, stubby, thin and filopodia, with mushroom spines being the most mature and filopodia the most immature (Harris et al., 1992). In a normal system, a mixture of these different types is present. Throughout development the distribution of spine types evolves to the mature state. Certain neurodevelopmental disorders are known to skew this distribution. For example, loss of the Fragile X Mental Retardation protein 1 (FMRP1), which in humans causes the mental retardation disorder Fragile X Syndrome, leads to an over-proliferation of thin spines and filopodia (i.e. immature spines) in cortical and hippocampal regions (Irwin et al., 2000). Conversely, mutations in MeCP2, a transcriptional regulator, which in humans causes Rett syndrome, leads to increased spine volume (Xu et al., 2014). This inability to maintain the correct balance of spine types may well be causal for the cognitive impairments which accompany these disorders. It is currently