NATHÁLIA WANDERLEY BUENO
VARIABILIDADE E DESEMPENHO FORENSE DE MICROSSATÉLITES UTILIZADOS NA VERIFICAÇÃO DE PARENTESCO DO PLANTEL BRASILEIRO DE CAVALO PURO SANGUE
INGLÊS (PSI)
Dissertação apresentada ao programa de Pós-Graduação Strictu Sensu em Ciências Genômicas e Biotecnologia da Universidade Católica de Brasília, como requisito parcial para a obtenção do Título de Mestre em Ciências Genômicas e Biotecnolgia.
Orientador: Doutor Dario Grattapaglia Co-orientador: Doutor Márcio Elias Ferreira
Ficha elaborada pela Coordenação de Processamento do Acervo do SIBI – UCB.
B928v Bueno, Nathália Wanderley.
Variabilidade e desempenho forense de microssatélites
utilizados na verificação de parentesco do plantel brasileiro de
cavalo puro sangue inglês (PSI) / Nathalia Wanderley Bueno. –
2008.
122 f.: il. ; 30 cm.
Dissertação (mestrado) – Universidade Católica de Brasília, 2008.
TERMO DE APROVAÇÃO
Dissertação de autoria de Nathália Wanderley Bueno, intitulada VARIABILIDADE E
DESEMPENHO FORENSE DE MICROSSATÉLITES UTILIZADOS NA VERIFICAÇÃO DE
PARENTESCO DO PLANTEL BRASILEIRO DE CAVALO PURO SANGUE INGLÊS (PSI), apresentada
como requisito parcial para a obtenção do grau de Mestre em Ciências Genômicas e Biotecnologia da
Universidade Católica de Brasília, em 24 de Agosto de 2007, defendida e aprovada pela banca
examinadora abaixo assinada:
_____________________________________________________
Prof. Dr. Dario Grattapaglia
Orientador
Professor do Programa de Pós-Graduação em Ciências Genômicas e Biotecnologia - UCB
_____________________________________________________
Prof. Dr. Márcio Elias Ferreira
Co-orientador
Professor do Programa de Pós-Graduação em Ciências Genômicas e Biotecnologia - UCB
_____________________________________________________
Dr. Samuel Rezende Paiva
Pesquisador da Embrapa Recursos Genéticos - Cenargen
_____________________________________________________
Prof. Dr. Alexandre Siqueira Guedes Coelho
Professor do Programa de Pós-Graduação em Genética e Melhoramento de Plantas - UFG
Brasília
Ao meu pai Eduardo (in memorian), à minha mãe Gilberta e irmão Vinícius pela educação e incentivo
AGRADECIMENTOS
Ao Prof. Dr. Dario Grattapaglia pela orientação técnica, disponibilidade e auxílio que me ajudaram na
realização deste trabalho.
Ao Prof. Dr. Márcio Elias Ferreira pela co-orientação e apoio.
Aos amigos da HERÉDITAS/GENOMAX, Mônica, Polys, Claudinha, Carol, Maria, Zilma, Saulo e
Ronaldo pelo auxílio na geração dos dados e segurar a barra em momentos de ausência. Ao André,
Milítize e Sandra pela organização dos dados cadastrais dos animais. Ao Josias, Tânia e Ellen pela
amizade.
Ao Adriano pela criação do DNAGRO e geração das simulações.
À ABCPCC pela coleta das amostras de sangue e organização de dados dos plantéis de PSI do Brasil.
Aos amigos do Cenargen, Eva, Marília e Isabela pela amizade e ajuda quando foi necessário.
Aos professores da UCB pelas instruções teóricas necessárias à realização deste trabalho.
Aos técnicos de laboratório da UCB André e Willian, pela ajuda na geração de dados.
Aos amigos da Pós-Graduação da UCB em especial Túlio, Breno, Érika e André.
Ao CNPq pela bolsa de estudos.
Aos meus familiares que me apoiaram nos meus projetos, em especial minha mãe Gilberta e meu
irmão Vinícius.
Ao meu pai Eduardo que com certeza estaria muito orgulhoso de mim.
Ao Alexandre pelo estímulo, amizade e amor.
Aos amigos, Gui, Pritt, Aline, André, Driano, Marcelo, Rê e as Jaques.
À Deus por estar sempre ao meu lado.
A todos que de forma direta ou indireta contribuíram de maneira relevante para a elaboração deste
RESUMO
A raça de cavalos Puro Sangue Inglês (PSI) é derivada de uma base genética consideravelmente restrita envolvendo poucos garanhões fundadores apresentando hoje uma diversidade alélica reduzida em comparação a outras raças de cavalos. Animais PSI demandam, entretanto, um rigor elevado no controle genealógico tendo em vista se tratar de uma raça internacional com ampla comercialização de animais, na qual o Brasil tem importante participação. Neste trabalho foi avaliado em detalhe o desempenho forense de uma bateria de 17 locos microssatélites amplamente utilizados, incluindo os nove marcadores recomendados pela International Society of Animal Genetics (ISAG), bem como a bateria de 13 locos TKY recentemente desenvolvida e incluída no exercício de comparação da ISAG de 2005-2006. Para a investigação do desempenho das baterias de marcadores, foram analisadas amostras de 597 éguas (PSI 597) e 142 garanhões (PSI 142) utilizados na geração do plantel brasileiro de cavalos PSI nascidos em 2004 e 2005. Ambas as baterias apresentaram múltiplos alelos e alta heterozigosidade e alcançaram um Poder de Exclusão (PE) acima de 99,95%. Para os locos TKY foi detectada uma pequena mas significativa redução de heterozigosidade devido a cruzamentos aparentados. O conjunto de nove locos internacionalmente recomendados pela ISAG alcançou um PE bem inferior àquele das baterias mais amplas seja em PSI bem como em outras raças de cavalos brasileiros. Em 23 trios analisados, nos quais duas inconsistências de transmissão alélica foram originalmente observadas, não foram observadas outras exclusões ao se analisar uma bateria adicional de 13 locos TKY o que corroborou as declarações originais de qualificação dos produtos. Estes resultados dão suporte experimental ao procedimento adotado no laboratório de declarar uma qualificação mesmo na presença de duas exclusões isoladas. Resultados de simulações indicam, entretanto, que este procedimento somente pode ser adotado com segurança ao se utilizar baterias mínimas de 15 a 16 locos ISAG e no caso de duos este procedimento é praticamente inviável a menos que baterias super expandidas de microssatélites sejam utilizadas. Recomendações de quais e quantos locos utilizar para a análise genética em raças eqüinas dependem de vários fatores. Simples comparações entre baterias de microssatélites devem ser vistas como procedimentos ad-hoc pois soluções alternativas podem sempre ser apresentadas variando quantos e principalmente quais locos utilizar. Os resultados deste trabalho sugerem que baterias de 13 locos TKY ou baterias de 14 locos selecionados envolvendo os nove locos ISAG obrigatórios, locos complementares e um loco TKY mais informativo, permitem alcançar um PE acima de 99,95%, limite este internacionalmente recomendado pelo ISBC (International Studbook Committee). As análises realizadas neste trabalho indicam por fim que os nove marcadores recomendados internacionalmente pela ISAG são insuficientes para uma adequada exclusão de paternidade em cavalos PSI. Além disso, a bateria de locos TKY constitui uma excelente ferramenta para a resolução de questões mais complexas de parentesco envolvendo supostos garanhões geneticamente relacionados ou quando exclusões isoladas são observadas nos painéis de microssatélites de rotina.
ABSTRACT
Thoroughbred horses (TB) is a breed derived from a considerably narrow genetic base involving a few founder stallions, displaying today a reduced allelic diversity when compared to other horse breeds. TB animals, however, demand a very high rigor in the genealogical control in view of its international reputation and ample worldwide trading in which Brazil plays an ever increasing role. In this study the forensic performance of a large set of microsatellite markers was evaluated including the nine ISAG recommended loci, 8 widely used complementary loci and the newly proposed battery of TKY markers validates in the 2005-2006 ISAG comparison trials. The forensic performance was evaluated in two samples of Brazilian PSI, one made up of 142 stallions (PSI 142) and the other of 597 mares (PSI 597) that took part in the generation of foals in 2004-2005. Markers in both sets showed ample allelic diversity and high heterozygosity and both loci sets reached a Power of Exclusion (PE) greater than 99,95%. For the TKY loci a very small but significant inbreeding was detected. The nine locus set recommended by ISAG as a minimum set reached a significantly lower PE not only in TB but also in other more diverse horse breeds raised in Brazil. In 23 paternity tests involving both parents and foal in which two isolated inconsistencies in allelic transmission were observed no further exclusions were detected after testing the 13 TKY locus set, therefore validating the original conclusion of qualification. These results provide experimental support to the procedure adopted in our lab to declare a foal as positively qualified as progeny of the tested stallion even when two isolated genotype inconsistencies are observed. Results of a simulation study indicate however, that such a procedure can only be adopted in complete trios when minimum sets of 15 ISAG markers are used and that this procedure is not viable when the dam is not tested, unless very large sets of microsatellites, over 30, are used. Recommendations about how many and which markers use in horse parentage testing depend on many factors. Simple comparisons among common sets of microsatellites should be seen as ad-hoc
approaches as alternative solutions can always be suggested varying how many and which loci are used. The results of this study also suggest that sets including 13 TKY loci or sets of 14 loci including the nine recommended by ISAG, complementary markers and a highly informative TKY locus can reach a PE above 99.95%, the ISBC (International Studbook Committee) recommended threshold. The analyses carried out also show that the nine ISAG markers are not sufficient for an adequate paternity exclusion in TB horse. Furthermore the TKY set of markers constitutes an excellent tool for the resolution of more complex situations involving genetically related stallions or when isolated exclusions are detected in routine parentage testing.
LISTA DE FIGURAS
Figura 1. Os três garanhões fundadores da raça Puro Sangue Inglês, Darley Arabian (à esq.), Goldolphin Arabian (meio), Byrley Turk (à dir.). ... 18
Figura2. Exemplar típico de Puro Sangue Inglês. ... 20
Figura 3. Cartão de coleta HEREDITAS/GENOMAX com identificação de código de barras. ... 34
Figura4. Conjunto de 15 locos TKY distribuídos em três multiplexes de acordo como o arranjo descrito por Tozaki et al. (2001a). Cada multiplex é formado por cinco locos, com diferentes faixas de tamanho de alelos, marcados com uma única fluorescência. Os três multiplexes podem ser carregados e
analisados simultaneamente no seqüenciador automático. ... 37
Figura5. Conjunto de 17 locos (ISAG + complementação) distribuídos em três multiplexes de acordo como o arranjo descrito por Ribeiro et al. (2005). ... 42
Figura 6. Eletroferogramas de DNA amplificado utilizando a bateria de 15 marcadores TKY, detectados em eletroforese no seqüenciador ABI377XL. Painel superior - Amostra controle do exercício ISAG
2005-2006 amplificado com os 15 marcadores TKY. Os locos em azul, verde a amarelo estão
marcados respectivamente com os fluorocromos 6-FAM, HEX e NED da Applied Biosystems. Painel inferior – Análise de transmissão alélica em trio. Somente os locos marcados em amarelo estão
apresentados. ... 46
Figura7. Eletroferogramas da análise de uma amostra de DNA com os três multiplexes envolvendo a bateria de nove marcadores microssatélites, recomendados pela ISAG, e oito marcadores adicionais,
no seqüenciador ABI377XL. Painel A - Multiplex A formado por oito locos (VHL20, HTG4, AHT4, HMS7, HTG6, HMS6, HTG7 e HMS3). Painel B - Multiplex A formado por oito locos (AHT39, UM011, AHT29, UM010, HMS6, UCDEQ425, HTG7 e UCDEQ405). Painel C - Multiplex C formado por quatro locos (AHT5, HMS6, HTG10 e ASB2). Os locos em azul, verde a amarelo estão marcados com os fluorocromos 6-FAM, HEX e NED da Applied Biosystems... 49
Figura8. Histogramas de distribuição de freqüências alélicas para 17 locos microssatélites utilizados em rotina nas raças eqüinas, Crioulo, Campolina e Puro sangue Inglês (PSI). Duas amostras
populacionais da raça PSI estão representadas. As cores dos histogramas correspondem às cores das
fluorescências utilizadas para detecção dos locos no seqüenciador automático. ... 67
Figura 9. Histogramas de distribuição de freqüências alélicas para os 9 locos microssatélites recomendados pela ISAG em planteis de Puro Sangue Inglês no Brasil (PSI 597 e PSI 142), Eslováquia
e Coréia. Não foram incluídos na análise os locos AHT4, AHT5 e HTG10 para a população de PSI da
correspondem às cores das fluorescências utilizadas para detecção dos locos no seqüenciador
automático. ... 75
Figura 10. Histogramas de distribuição de freqüências alélicas para os 13 locos microssatélites TKY em planteis de Puro Sangue Inglês no Brasil (PSI 142) e do Japão (PSI JP 250). As cores dos
histogramas correspondem às cores das fluorescências utilizadas para detecção dos locos no
seqüenciador automático. ... 82
Figura 11. Distribuição de freqüência de exames em trios completos nos quais o garanhão seria excluído com números crescentes de locos diagnósticos de exclusão ao se utilizar baterias de 11 até
16 marcadores microssatélites ISAG. ... 94
Figura12. Distribuição de freqüência de exames em duos (sem a participação da genitora) nos quais o garanhão seria excluído com números crescentes de locos diagnósticos de exclusão ao se utilizara
baterias de 11 até 16 marcadores microssatélites ISAG. ... 95
Figura13. Poder de Exclusão de paternidade estimado por simulações, considerando a exigência de observação de pelo menos 3 locos diagnósticos de exclusão ao se testar baterias crescentes de locos
no exame em animais PSI. São apresentadas as situações de exames de duo (ausência de um dos
LISTA DE TABELAS
Tabela1. Bateria de 15 marcadores microssatélites TKY. Os multiplexes cujos locos são detectados nas fluorescências azul, verde e amarela são compostos por 5 locos cujos primers diretos são
marcados no terminal 5’ com os fluorocromos 6-FAM, HEX e NED, respectivamente. ... 35
Tabela2. Bateria de 17 marcadores microssatélites, incluindo os nove marcadores recomendados pela ISAG indicados com *. (MPX = multiplex; n.d = não disponível) ... 40
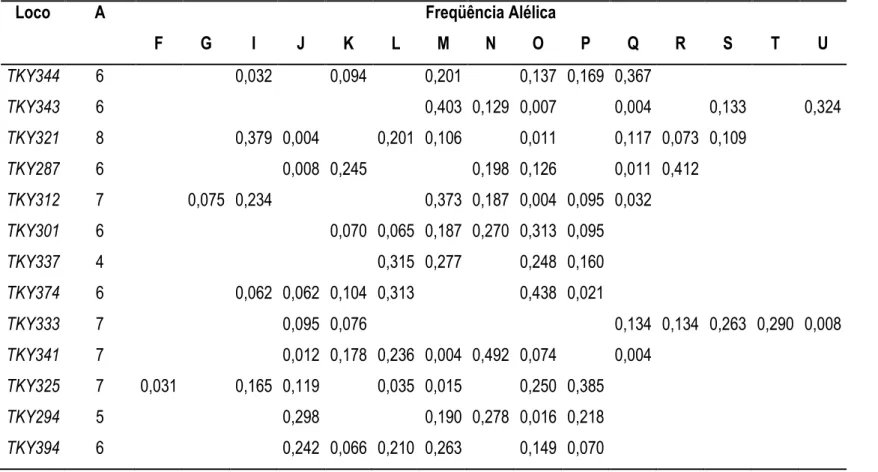
Tabela3. Número de alelos por loco (A) e distribuição de freqüências alélicas em 13 microssatélites TKY baseados em uma amostra de 142 garanhões do plantel brasileiro de PSI. Os alelos foram
identificados por letras alfabéticas de acordo com a nomenclatura recomendada pelo ISAG. ... 48
Tabela 4. Número de alelos por loco (A) e distribuição das freqüências alélicas em 17 microssatélites ISAG , baseados em duas amostras de animais PSI, 142 garanhões (PSI 142) e 597 éguas (PSI 597).
... 50
Tabela5. Parâmetros genéticos de desempenho dos 17 microssatélites estudados para a investigação de individualidade genética e de paternidade em duas populações de PSI; estimativas de coeficiente de
endocruzamento (Fis) e p valor do teste exato para Equilíbrio de Hardy Weinberg. (# Intervalo de
confiança a 95% sobre a estimativa de Fis total). ... 53
Tabela 6. Teste exato de Fisher para equilíbrio de ligação de locos dois a dois. Todos os pares de locos na tabela apresentaram valores significativos com a correção de Bonferroni (α = 0,00036). ... 54
Tabela 7. Parâmetros genéticos de desempenho forense dos 13 microssatélites TKY estimados na amostra de 142 garanhões PSI brasileiros e comparação com uma amostra populacional de 250 PSI
japonês; estimativas de coeficiente de endocruzamento e teste para Equilíbrio de Hardy Weinberg (#
Intervalo de confiança a 95% sobre a estimativa de Fis total). ... 57
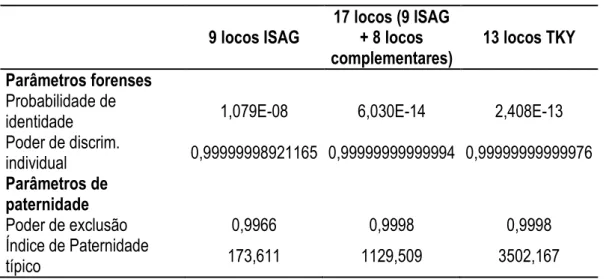
Tabela8. Parâmetros de desempenho forense e paternidade de três diferentes baterias de marcadores microssatélites na população PSI 142. ... 58
Tabela9. Parâmetros genéticos de desempenho dos 17 microssatélites (ISAG+Complementação) em três raças de eqüinos; estimativas de coeficiente de endocruzamento (Fis) e pValor do teste para
Equilíbrio de Hardy Weinberg (# Intervalo de confiança a 95% sobre a estimativa de Fis total). ... 69
UCDEQ425, UCDEQ405), em três raças de eqüinos criadas no Brasil (CRI Crioulo; CAM Campolina; PSI Puro Sangue Inglês). ... 70
Tabela11. Poder de exclusão de cada loco individualmente e dos locos combinados para os locos 17 ISAG na presença de parentesco entre garanhões testados, estimados com base nas amostras de
animais PSI 597 e PSI 142. ... 84
Tabela12. Poder de exclusão de cada loco individualmente e dos locos combinados para os locos TKY na presença de parentesco entre garanhões testados, para estimado com base na amostra de 142
garanhões PSI brasileiros. ... 85
Tabela13. Poder de exclusão de cada loco individualmente e dos locos combinados para a bateria de rotina envolvendo nove locos ISAG, quatro locos complementares e um loco TKY na presença de
parentesco entre garanhões testados, para estimado com base na amostra de 142 garanhões PSI
brasileiros. ... 85
Tabela14. Genótipos de 23 trios testados nos quais foram observadas duas inconsistências isoladas de transmissão alélica na bateria de 17 locos testados e em seguida complementados com 13 locos
TKY. Os locos nos quais foi detectada uma inconsistência estão destacados em cinza. ... 87
SUMÁRIO
1. INTRODUÇÃO ... 10
2. REVISÃO BIBLIOGRÁFICA... 17
2.1 História e origem da raça Puro Sangue Inglês ... 17
2.2 Registros genealógicos da raça Puro Sangue Inglês ... 20
2.3 Verificação de parentesco em eqüinos via tipagem sangüínea ... 22
2.4 Marcadores microssatélites em eqüinos ... 25
2.5 Marcadores microssatélites para identificação e parentesco em eqüinos ... 29
3. OBJETIVOS ... 32
4. MATERIAL E MÉTODOS... 33
4.1 Coleta de amostras e extração de DNA ... 33
4.2 Utilização de multiplexes de locos microssatélites TKY... 34
4.3 Banco de dados e desempenho forense dos locos TKY ... 38
4.4 Desempenho forense dos locos ISAG e verificação de parentesco em PSI brasileiro ... 40
4.5 Análise genética comparativa do PSI brasileiro com PSI de outros países e outras raças ... 43
4.6 Estimativa do poder de exclusão em situações de parentesco ... 43
4.7 Análise de locos complementares em casos de inconsistências (exclusões) isoladas ... 44
4.8 Estimativa de PE realizado e simulações de exames de exclusão ... 44
5. RESULTADOS ... 45
5.1 Banco de dados dos locos microssatélites TKY ... 45
5.2 Banco de dados de freqüências alélicas dos locos ISAG ... 49
5.3 Diversidade genética e desempenho forense dos locos ISAG e TKY e comparações ... 51
5.4 Comparações de PSI brasileiros com PSI de outros países e outras raças criadas no Brasil 58 5.5 Estimativa do poder de exclusão em situações de parentesco entre garanhões ... 83
5.6 Análise de locos complementares em casos de inconsistências (exclusões) isoladas ... 86
5.7 Estimativas de PE realizado e simulações de exames de exclusão ... 93
6. DISCUSSÃO ... 97
7. CONCLUSÕES ... 108
1. Introdução
A identificação individual e determinação de vínculo genético é rotina nos serviços de registro
genealógico de animais domésticos e está sendo aplicada de forma crescente nos programas de
melhoramento genético animal em diversos países do mundo. A utilização de marcadores genéticos
para a verificação de parentesco foi adotada nos serviços de registro de raças de cavalos nos anos 60
após os trabalhos pioneiros de Stormont e Suzuki (1964, 1965) os quais descreveram a herança e
alelismo de 16 fatores sangüíneos identificados em cavalos. Por aproximadamente três décadas, estes
sistemas envolvendo grupos sangüíneos e variantes protéicos vêm sendo utilizados com sucesso.
Esses testes, embora rápidos e baratos requerem o uso de sangue fresco, apresentam poder de
discriminação limitado e demandam a produção de reagentes biológicos in vivo, processo complexo e pouco accessível. Embora ainda estejam sendo utilizados, a transição para a utilização exclusiva de
marcadores de DNA é hoje considerada irreversível. Nos últimos 15 anos, metodologias de análise de
polimorfismos genéticos baseadas em variações de seqüências no DNA usando a tecnologia da PCR
(Polymerase Chain Reaction) forneceram alternativas à tipagem sangüínea em cavalos, particularmente a análise de polimorfismos de microssatélites ou SSR (Simple Sequence Repeats) (Bowling et al., 1997). A análise de microssatélites hipervariáveis fornece hoje a mais alta precisão na identificação genética em seres vivos, constituindo uma ferramenta indispensável de certificação de
qualidade genealógica de produtos nos programas de registro genealógico. Muitas são as vantagens
dos testes de DNA em relação à tipagem sangüínea, entre elas a possibilidade de se utilizar para
exame não somente amostras de sangue, mas também sêmen, bulbos de pêlos e outros tecidos,
ampliando as possibilidades de verificação de identidade ou parentesco para situações de investigação
forense animal.
A genotipagem do DNA de animais se baseia na determinação do genótipo multilocal para um
resolução, tipicamente em seqüenciadores automáticos. Com base nestes genótipos é definido um
perfil de DNA único, altamente discriminativo, comumente chamado de ―impressão digital de DNA‖.
Este perfil genético pode ser utilizado, entre outros, para: (a) Verificação de paternidade e maternidade
para registro genealógico de animais; (b) Registro de animais importados cujos pais somente possuem
tipagem de DNA; (c) Controle de qualidade de programas de inseminação artificial e transferência de
embrião; (d) Determinação de paternidade em sistemas de cruzamento com múltiplos reprodutores; (e)
Estimativa de distância genética entre animais para introgressão de nova variabilidade ou planejamento
de cruzamentos; (f) Suporte ao melhoramento para estimativas de valor de melhoramento de animais
elite; (g) Certificação e controle de qualidade de sêmen.
Várias centenas de marcadores microssatélites foram publicados para eqüinos nos últimos 15
anos (ex. Ellegren et al., 1992; Binns et al., 1995; Tozaki et al., 2004). A Sociedade Internacional de Genética Animal (ISAG - International Society of Animal Genetics), no sentido de facilitar o intercâmbio e comparação de resultados de genotipagem entre laboratórios vem recomendando um conjunto
básico de nove marcadores microssatélites para raças eqüinas. A definição destes nove marcadores
ocorreu de forma ad hoc com base em alguns poucos relatos da literatura (ex. Bowling et al., 1997) e experiência de alguns poucos laboratórios (Veterinary Genetics Laboratory – US Davis), procurando atender a esta demanda específica e sem planejamento prévio ou avaliações mais aprofundadas sobre
a robustez analítica ou conteúdo informativo dos nove marcadores. No Brasil, o Ministério da
Agricultura, Pecuária e Abastecimento (MAPA), ciente da necessidade de evolução e adequação dos
Serviços de Registros Genealógicos das diversas associações brasileiras de criadores aos padrões
internacionais, deliberou acerca da regulamentação das normas a serem adotadas nos serviços de
análise de DNA. O MAPA, através da Instrução Normativa nº 74 de 20 de outubro de 2004
(http://www.in.gov.br/imprensa/in), que estabelece as normas para credenciamento de laboratórios de
determinação de vínculo genético em eqüinos no Brasil. Estes marcadores são os mesmos
recomendados pela ISAG para verificação de paternidade e maternidade em eqüinos
(http://www.isag.org.uk). Alguns marcadores adicionais tais como ASB17, ASB23, LEX33, HMS2, HTG6 e HTG7, vêm também sendo utilizados por diversos laboratórios participantes da ISAG para a complementação de genotipagem em casos especiais. Nos últimos anos, a robustez analítica dos nove
marcadores adotados pela ISAG vem sido avaliada em testes comparativos envolvendo dezenas de
laboratórios no mundo com um grau considerável de concordância entre os laboratórios.
Embora a bateria de nove marcadores forneça um poder de exclusão mais elevado do que a
bateria de marcadores de grupos sangüíneos e polimorfismos protéicos tradicionalmente utilizados,
ainda assim em raças eqüinas que se caracterizam por elevada consangüinidade, marcadores
adicionais freqüentemente se fazem necessários para atingir um poder de exclusão de paternidade (ou
maternidade) (PE) efetivo maior do que 99,99%. No caso específico da raça Puro Sangue Inglês (PSI),
derivada de uma base genética consideravelmente restrita, um forte efeito fundador de apenas três
garanhões no século XVII (veja histórico a seguir), e um sistema de acasalamentos consangüíneos
deliberado, a reduzida variabilidade alélica mesmo a locos microssatélites tem sido documentados em
algumas publicações recentes (Cunningham et al., 2001; Tozaki et al., 2001a; Lee e Cho, 2006). Conjuntos de marcadores microssatélites relativamente bem estabelecidos vêm sendo
utilizados em diversos laboratórios do mundo. Entretanto quais e quantos microssatélites devem ser
utilizados ainda é uma questão debatida no âmbito do grupo de trabalho de eqüinos da ISAG. As
principais considerações para a seleção de microssatélites são a sua robustez para análise em rotina, o
seu poder de exclusão e a estrutura genética e histórico evolutivo da raça em questão. Os resultados
disponíveis na literatura sugerem que o número adequado de marcadores deverá ser maior do que os
nove recomendados pela ISAG, principalmente para raças com maior nível de consangüinidade. Quais
os marcadores a serem utilizados torna-se uma questão mais complexa, pois passa por uma avaliação
que é geneticamente mais informativo para uma raça pode não o ser para outra. Outra consideração
importante do ponto de vista prático é a possibilidade de se combinar os diferentes locos de forma
eficiente em uma mesma reação de PCR com o objetivo de maximizar a geração de dados por unidade
de tempo reduzindo o número e reações de PCR e análises eletroforéticas necessárias para concluir a
tipagem de um animal. Finalmente, a definição de quais locos utilizar deve ainda respeitar uma
recomendação internacional de forma a permitir a comparação de dados entre diferentes laboratórios e
a utilização de tipagens feitas em diferentes países para fins de verificação de parentesco.
Uma questão relevante ao se realizar exames de paternidade e maternidade em populações
subestruturadas ou com forte histórico de cruzamento aparentados é o risco de declaração de falsas
inclusões, ou seja, qualificar um determinado produto como sendo filho de um suposto genitor ou
genitora quando na verdade o genitor não é o pai ou mãe biológico(a) do produto. Este risco, embora
pequeno, se torna maior em situações nas quais o verdadeiro pai biológico é aparentado do suposto
pai testado.
O poder estatístico de um loco para análise de paternidade é geralmente avaliado pelo poder
de exclusão (PE), ou seja, a probabilidade do loco excluir da paternidade um indivíduo tomado ao
acaso da população que foi erroneamente apontado como pai (Weir, 1996). A expressão que calcula
PE assume que o macho tido como suposto pai foi tomado ao acaso da população e de que outros
machos que eventualmente estejam competindo pela paternidade sejam também uma amostra ao
acaso de uma população panmítica. No caso de eqüinos de raças consangüíneas, uma fonte
importante de subestruturação de população que leva à violação da premissa de panmixia ocorre
justamente quando dois ou mais garanhões geneticamente relacionados, irmãos, pai-filho ou mesmo
primos, são testados como supostos pais em uma verificação de paternidade. Nestes casos a
estimativa de PE para um determinado loco será superestimada, pois nestas circunstâncias o poder de
exclusão será consideravelmente menor do que o esperado pela premissa de panmixia (Jeffreys e
exceção do pai verdadeiro hoje existente na literatura para eqüinos devem ser consideradas, em geral,
superestimadas. Além disso, a real probabilidade de exclusão é afetada não apenas pela presença de
um segundo macho geneticamente relacionado, mas também pelo número total de machos
relacionados existentes que competem pela paternidade. Quanto maior o número de machos
competindo pela paternidade, maior a probabilidade de um deles ser erroneamente declarado como
sendo o verdadeiro pai (Double et al., 1997; Sherman et al., 2004).
Um segundo aspecto freqüentemente levantado em exames de parentesco com
microssatélites é a ocorrência de exclusões isoladas, isto é, exclusões em apenas um loco do conjunto
total de locos analisados. A recomendação internacional nestes casos é a declaração de uma inclusão
e a informação da inconsistência alélica encontrada, tendo em vista que eventos desta natureza podem
ser explicados por mutações ou ocorrência de alelos nulos devido a mutações que impedem a
amplificação de alelos ao loco sob análise (Chakraborty e Stivers, 1996). Ocorrências de
inconsistências isoladas foram relatadas em eqüinos, e estimativas de taxa de mutação da ordem de
0,01 a 0,02% por geração foram relatadas para os locos AHT4 e HTG10 (Bowling et al., 1997). Em seres humanos, onde um grande número de exames de paternidade vem sendo realizado no mundo
em diversos laboratórios, já foram relatados casos de duas incompatibilidades isoladas em exames
nos quais de fato o suposto pai era o pai biológico e as mutações confirmadas (Gunn et al., 1997; Grattapaglia com. pess.). Um estudo em humanos baseado na bateria internacionalmente
recomendada de 13 microssatélites e as respectivas taxas de mutação relatadas na literatura, relatou
resultados interessantes a este respeito (Calafell, 2000). Em casos de falsa paternidade, exclusões são
declaradas, em média, 7,65 locos excludentes dos 13 analisados. A freqüência de casos de falsa
paternidade com a observação de apenas uma exclusão foi estimada em 0,0108% ou seja um caso a
cada 9.259 e com duas exclusões em 0,14% ou seja 1 caso a cada 704. Considerando, por outro lado,
casos de paternidade verdadeira e a ocorrência de mutações, em 4,85% dos casos foi observada uma
exclusões isoladas oriundas de mutações. Em outras palavras, a probabilidade de ocorrência de duas
mutações na análise de 13 marcadores é relativamente alta principalmente se um grande número de
exames é realizado. Tomando agora situações nas quais o suposto pai testado era irmão do verdadeiro
pai biológico, o número médio de locos excludentes caiu de 7,65 para apenas 3,89 e em 0,98% ou
seja, em cerca de 1 caso a cada 100, nenhuma exclusão foi observada (Calafell, 2000).
Em eqüinos ainda não foram relatados casos de mais de uma inconsistência alélica isolada em
exames de paternidade, embora tendo em vista o grande número de exames realizados atualmente,
isso seja possível. A questão que fica em populações endocruzadas de cavalos PSI é como proceder
em exames nos quais são observadas apenas duas exclusões isoladas. Tendo em vista o alto nível de
endocruzamento em PSI, estimado em 0,13 em uma amostra de 211 cavalos (Cunningham et al., 2001), espera-se que em casos de falsa paternidade poucos locos excludentes sejam observados
gerando, portanto, uma situação de dúvida: trata-se de fato de uma real exclusão ou de uma
ocorrência de mutações? Bowling et al. (1997) estudando cavalos Quarto de Milha declararam exclusão mesmo observando apenas duas exclusões isoladas. Embora esta em geral tenha sido a
prática em laboratórios que realizam exames de paternidade, a ISAG tem recomendado que nestes
casos seja analisada uma bateria complementar de locos visando identificar exclusões adicionais e
com isso aumentar a confiabilidade da declaração da exclusão (Binns et al., 1995). Mutações em locos comumente utilizados em exames de paternidade de eqüinos têm sido descritos, mas ainda de forma
muito incipiente. Eggleston-Stott et al. (1997a) descreveram perda ou baixa eficiência no anelamento do primer HMS7 devido a transversão de C para A, resultando em alelo nulo. Neste caso, o alelo não é detectável no produto da PCR e um aparente homozigoto é observado o qual pode ser erroneamente
interpretado como uma transmissão não mendeliana (Callen et al., 1993). Guérin et al. (1994) observaram anteriormente, uma aparente transmissão não mendeliana no loco HMS7 em cavalos quarto de milha, na qual uma égua foi excluída como genitora de dois produtos, baseado no fato de
alelo em comum. Como a égua foi qualificada genitora em outros 20 locos, foi levantada a suspeita de
baixo rendimento da detecção do produto da PCR. Achmann et al. (2001) revelou que incorretas exclusões foram interpretadas devido a identificação errônea de alelos em genótipos particulares. Estes
erros ocorreram porque os alelos M do marcador HMS3 e L do marcador ASB2 foram fracamente amplificados e assim não pode ser distinguido do stutter do alelo N (HMS3) e M (ASB2), respectivamente. Nestes casos, exclusões baseadas em observações de genótipos homozigotos para
diferentes alelos entre produto de suposto genitor devem ser considerados com cautela.
Este trabalho teve por objetivo avaliar em detalhe o desempenho de uma ampla bateria de
marcadores microssatélites para verificação de parentesco em cavalos Puro Sangue Inglês criados no
Brasil. Esta raça possui uma base genética estreita devido a um importante efeito fundador mas ao
mesmo tempo demanda um rigor elevado no controle genealógico dos seus animais tendo em vista se
tratar de uma raça internacional com ampla comercialização de animais. O estudo visou avaliar o
conjunto de locos recomendado pela ISAG, locos complementares de freqüente utilização internacional
bem como a bateria de locos TKY recentemente incluída no exercício de comparação da ISAG de
2005-2006 e que deverá ser cada vez mais utilizada em análises de rotina no mundo. O estudo visou
especificamente avaliar o desempenho forense destes marcadores em populações de animais
nascidos no Brasil ano de 2004 bem como no plantel de garanhões mais amplamente utilizados no país
procurando avaliar o grau de ancestralidade comum entre os mesmos. Foram realizadas simulações
com base no conjunto de animais genotipados visando avaliar o número médio de locos excludentes
em casos de falsas paternidades bem como em casos de paternidades verdadeiras. Finalmente foi
estimado ainda o poder de exclusão de cada loco e de combinações de locos para situações nas quais
o suposto pai testado é parente de primeira ordem do verdadeiro pai biológico do produto, fornecendo
assim uma visão mais realista do real poder de exclusão da análise genética com microssatélites na
2. Revisão bibliográfica
2.1 História e origem da raça Puro Sangue Inglês (PSI)
Por volta de 1500, as corridas de cavalos se constituíam em um divertimento dos senhores das
terras aos quais, as atividades de suas fazendas não lhes permitiam grandes perspectivas de
passarem os domingos e feriados de maneira mais alegre. Assim, faziam correr seus animais pesados
e carentes de velocidade. Foi este pequeno grupo de criadores que nos séculos XVI e XVII começaram
com o cruzamento de cavalos nativos em éguas importadas da Espanha, Turquia e Itália.
É possível determinar quando a raça de cavalo Puro Sangue Inglês - PSI (em inglês
Thoroughbred - TB) desenvolveu-se. Relatos históricos indicam que por volta de 1600 a 1750, cerca de 174 reprodutores orientais, árabes, berberes e turcos, foram importados, com o propósito de
incrementar a velocidade dos cavalos utilizados para o popular esporte de corridas. Destes, três
garanhões fundadores tornaram-se famosos pela sua contribuição na formação da raça cuja
ancestralidade remonta essencialmente a Darley Arabian, Godolphin Arabian e Byerly Turk (Figura 1),
batizados por seus respectivos proprietários Thomas Darley, Lord Godolphin e Captain Robert Byerly.
Estes três garanhões foram cruzados com os cavalos nativos ingleses mais fortes, porém menos
precoces. O resultado foi um animal que podia carregar peso com velocidade constante por grandes
distâncias, qualidades estas que trouxeram uma nova perspectiva para o esporte aristocrata e em
franco crescimento na época que era o da corrida de cavalos. A incorporação destes garanhões na
reprodução em diferentes épocas, mediante intensa consangüinidade, produziu, em última instância, o
moderno cavalo Puro Sangue Inglês, de forma que cada um dos vários milhões de animais atuais de
Puro Sangue Inglês, ao redor do mundo remonta a este restrito grupo de fundadores
Figura 1. Os três garanhões fundadores da raça Puro Sangue Inglês, Darley Arabian (à esq.), Goldolphin Arabian (meio), Byerley Turk (à dir.).
O primeiro reprodutor a chegar, foi o turco, Byerley nascido em 1680. Seu proprietário era um
coronel que o havia arrebatado aos turcos durante a batalha de Buda (Hungria). Ao longo de vários
anos se serviu daquele animal como seu cavalo de guerra, e mais tarde, quando se retirou da vida
militar em 1690, o destinou à reprodução. Os potros nascidos de Byerley alcançaram pouco renome,
mas seu bisneto, Herod, embora não tivesse se destacado como um grande corredor, projetou-se
como um dos principais progenitores da raça PSI e gerou outros ganhadores de 1042 corridas.
Darley Arabian nasceu em 1700 na Síria e foi importado para Inglaterra, onde teve a primeira
geração de descendentes, dentre eles descatou-se o Flying Childers, o primeiro grande cavalo de
corridas. Através de seu outro filho, Bartlett's Childers, tornou-se tataravô de Eclipse, possivelmente o
cavalo de corrida mais famoso de todos os tempos. Eclipse gerou 344 ganhadores de dele resultaram
linhagens de grande importância no desenvolvimento da raça PSI.
A origem do terceiro garanhão fundador da raça PSI, o árabe/berbere Godolphin Arabian é
ainda incerta. Godolphin nasceu em 1724 e integrava um grupo de oito cavalos presenteados ao rei da
França Luís XIV em 1731. Sua história possui três versões, uma conta que aos cinco anos foi levado
da França para Inglaterra, por Edward Coke, que o encontrou puxando uma carroça de água nas ruas
de Paris. Outra se refere que foi comprado por três libras de um carroceiro em Paris e presenteado ao
Lord Godolphin passando a servir em seu haras. A última história relata que o cavalo teria sido roubado
notáveis cavalos, entre eles o Lath e o famoso Cade (1734) pai de Matchem, que como Eclipse e
Herod, havia de fundar uma destacada estirpe do PSI.
A história da criação de cavalo de corrida no Brasil teve início em 1874, ano em que ocorreu o
nascimento do potro Brasil na Província de Minas Gerais, que viria a ser o primeiro garanhão nacional
Puro Sangue. O genitor e genitora de Brasil são os cavalos ingleses Zephyro e Hierogliph,
respectivamente, descendentes do conceituado alazão Eclipse (Brotto, 1979).
A aparência dos cavalos PSI revela uma ancestralidade árabe. Eles caracterizam-se por
apresentarem muita finura, beleza e grande classe. Apresentam uma altura média de 1,60 m, linda
cabeça, perfil reto ou levemente ondulado, fronte ampla, olhos grandes, narinas elípticas e dilatadas,
orelhas médias, pele fina, cernelha destacada e musculosa, dorso reto comprido e lombo curto, garupa
inclinada, peito estreito e tórax profundo. Possuem espáduas inclinadas, membros fortes, joelhos
baixos e canelas curtas. A pelagem em geral é uniforme e pode variar em cor sendo as principais
categorias castanho, alazão, preto e tordilho. Marcas brancas na face e pernas são freqüentemente
observadas e são importantes descritores morfológicos na resenha do animal (Figura 2) (Brotto, 1979).
O cavalo PSI é um dos cavalos mais brilhantes e versáteis criados no mundo hoje. Sua principal
característica se refere à sua capacidade de carregar peso por longas distâncias, ou seja, a
combinação entre velocidade e resistência. Em função disso o cavalo PSI é popular não apenas para
corridas, mas também em outras disciplinas tais como caça, saltos e pólo. O cavalo PSI tem tido ainda
papel de destaque na criação de novas raças de cavalos e para o melhoramento genético de raças
Figura 2. Exemplar típico de Puro Sangue Inglês.
2.2 Registros genealógicos da raça Puro Sangue Inglês
Desde o século XVII, com a criação da raça PSI, começou um processo de seleção direcional
que vem acontecendo por mais de 300 anos sempre intercruzando os melhores garanhões com as
melhores éguas, sendo que a prova de superioridade e excelência é estabelecida de forma objetiva na
pista de corrida. Um aspecto chave para o sucesso deste programa contínuo de melhoramento
genético baseado em forte consangüinidade tem sido a integridade dos registros genealógicos desta
raça. Antigamente, os registros de pedigrees eram desconexos e freqüentemente incompletos mesmo
porque era costume batizar cavalos somente após o mesmo comprovar uma capacidade de destaque
nas pistas de corrida.
Foi somente com o Sir James Weatherby, com seus recursos próprios e pesquisa pessoal que
foi possível a consolidação de um grande número de registros genealógicos privados que resultou na
publicação do primeiro livro de registros o General Stud Book. Ele completou isso em 1791, listando os pedigrees de 387 éguas, todas elas que podiam ser relacionadas por ancestria à Eclipse, um cavalo
Byerly Turk. Este livro geral de registros, o General Stud Book ainda é publicado na Inglaterra pela Weatherby e Sons, secretários do Jockey Club (www.thejockeyclub.co.uk).
Muitos anos depois, com a proliferação de corridas de cavalo no continente norte americano, a
necessidade de um serviço de registros genealógicos de cavalos PSI nascidos nos EUA, nos moldes
do General Stud Book, se tornou aparente. O Coronel Sanders D. Bruce, morador de Kentucky publicou o primeiro volume do American Studbook em 1873 e até 1896 produziu mais seis volumes do
registro quando então o projeto foi assumido pelo Jockey Club dos EUA
(http://www.jockeyclub.com/registry.asp). Este mesmo procedimento foi adotado por outros países nos
últimos 50 a 100 anos. No Brasil o Serviço de Registro Genealógico do Cavalo de Corrida, denominado
"Stud Book Brasileiro", tem por finalidade a manutenção do controle genealógico do cavalo da raça Puro Sangue de corrida e seus mestiços. Este serviço funciona na dependência da Associação
Brasileira de Criadores e Proprietários do Cavalo de Corrida (ABCPCC) e é por esta administrado em
todo o território nacional, por delegação expressa do órgão competente do Ministério da Agricultura
Pecuária e Abastecimento (MAPA), com fundamento na Lei nº 4.716 de 29 de junho de 1965.
O objetivo essencial do Stud Book Brasileiro é a manutenção dos registros genealógicos e de identidade dos cavalos Puro Sangue de Corrida, zelando pelo aprimoramento e pureza da raça. Este
serviço mantém estreito e permanente relacionamento com Stud Books de outros países, exercendo o controle e fiscalização da procriação, gestação, nascimento, identificação e filiação, inscrição de
animais importados, outorga de certificados de exportação, de identidade, de propriedade e qualquer
outra documentação relacionada com tais atividades (www.studbook.com.br).
A integridade de um serviço de Stud Book é a fundação sobre a qual todo o sistema de criação, comercialização, melhoramento e seleção de cavalos de corrida se baseia hoje. Além dos
aspectos de registro de dados cadastrais dos animais, é peça chave neste sistema a integridade e
certeza dos parentescos declarados entre os animais. Um passo importante na certificação da
programa de verificação de parentesco de animais pelo sistema de tipagem sangüínea desenvolvido no
início da década de 60 (Stormont e Suzuki, 1964; 1965). Nos EUA desde o final da década de 70 até
2000 todo animal PSI nascido e registrado no American Stud Book bem como ambos os seus pais foram tipados para um conjunto de grupos sangüíneos para verificação do parentesco declarado.
Recentemente, o Comitê Internacional do Stud Book (ISBC –International Stud Book Comittee) tem requerido um poder de exclusão mais elevado que o alcançado por tipagem sangüínea.
Conseqüentemente, o Jockey Club americano substituiu o sistema de tipagem sangüínea pela análise de marcadores microssatélites no DNA aumentando a precisão de 97% (Bowling et al,. 1985) para 99,9% (Bowling et al., 1997), respectivamente, começando com os produtos nascidos em 2001. No Brasil, a verificação de paternidade por tipagem sangüínea passou a ser requerida para cavalos PSI
para todas as crias nascidas após 1° de janeiro de 1992. Em 2005, o Stud Book Brasileiro com o intuito de cumprir as exigências nacionais e internacionais e manter a criação nacional rigorosamente dentro
dos padrões exigidos mundialmente, iniciou o programa de verificação de parentesco com base na
análise de DNA. A colheita de amostras para realização do exame teve início em 17/10/2005 visando à
verificação dos produtos nascidos no ano de 2004. Para a realização das verificações foram também
colhidas amostras do plantel de garanhões e éguas (www.studbook.com.br).
2.3 Verificação de parentesco em eqüinos via tipagem sanguínea
Pelos últimos 20 a 30 anos, os programas de verificação de parentesco de cavalos PSI bem
como de diversas outras raças se basearam na tipagem sangüínea, i.e. grupos sangüíneos e
polimorfismos protéicos. Esta tecnologia se baseia em uma série de rotinas confiáveis, tecnicamente
simples e de baixo custo ainda hoje oferecidas por vários laboratórios no mundo, embora sendo
rapidamente substituídas pela análise de DNA. A tipagem sangüínea consiste no reconhecimento de
fatores sangüíneos (antígenos) presentes na superfície das células vermelhas do animal, com os
interpretadas reações de hemólise e/ou aglutinação das hemácias do animal, quando submetidas à
presença de anticorpos específicos para os diversos fatores sangüíneos. Os anticorpos podem ser
ocasionalmente coletados de éguas imunizadas pelo seu próprio sangue. Esses anticorpos naturais
produzem uma fraca reação hemolítica significativa in vivo dificultando a interpretação dos resultados. O descobrimento de variabilidade em fatores sangüíneos em eqüinos foi relatado originalmente
em uma série de estudos por Podliachouk (1958) na França, o qual descreveu a existência de dez
fatores sangüíneos peculiares a cavalos e mulas. Logo em seguida, nos Estados Unidos, Franks
(1962) descreveu onze fatores sangüíneos. Estes estudos, entretanto, não haviam estabelecido
relações de alelismo entre estes fatores e apenas citavam associações entre fatores atribuindo a
ligação gênica. A ausência de uma interpretação mendeliana correta da herança destes fatores de
certa forma limitava a devida interpretação para estudos de parentesco. Foi somente com os estudos
de Stormont e Suzuki (1964) que estabeleceram a relação de alelismo e demonstraram a herança
destes fatores sangüíneos que se iniciou a utilização sistemática de grupos sangüíneos para
verificação de parentesco em eqüinos. Neste trabalho pioneiro foram descritos 16 fatores sangüíneos,
demonstrada a sua herança mendeliana em 639 cruzamentos envolvendo 103 garanhões de diferentes
raças e estimadas as suas freqüências alélicas em duas raças com 391 animais da raça de Ponies
Shetland e 276 Puro Sangue Inglês (Thoroughbred). Este trabalho definiu de forma clara a existência de oito locos responsáveis pela produção dos 16 fatores sangüíneos e conseqüentemente pelo controle
dos grupos sangüíneos em cavalos.
Do ponto de vista prático, pouco mudou desde estes estudos pioneiros e embora nenhum novo
grupo sangüíneo foi descoberto, o poder de resolução da tecnologia foi ampliado pela introdução de
variantes protéicos. Testes bioquímicos analisam as principais variantes protéicas presentes nas
hemácias ou no plasma dos animais através de técnicas de eletroforese em gel de amido. Os oito
sistemas usualmente detectados em testes com hemácias e plasma são: albumina, erastase, vitamina
glicoproteína. Hoje, são regularmente testados 7 sistemas de grupos sangüíneos, compreendendo 34
fatores sangüíneos diferentes e complementados por 6 variantes protéicas distintas. Bowling e Clark
(1985) estimaram as freqüências alélicas para 20 grupos sangüíneos e locos protéicos polimórficos (A,
C, D, K, P, Q, U, Al, Tf, Pi, Xk, Es, Gc, PGD, CA, Cat, PGM, AP, Hb e PHI) em sete raças de cavalos
nos Estados Unidos (Puro Sangue Inglês, Árabe, Standardbred, Morgan, Quarto de Milha, Paso Fino e
Peruvian Paso). Os dados demonstraram que esta bateria de marcadores apresentava uma
probabilidade de exclusão de falsa paternidade de pelo menos 96% em Puro Sangue Inglês chegando
a 99% em Quarto de Milha. Bernoco et al. (1997), em sua revisão, cita estudos não publicados nos quais, ao se utilizar 5 sistemas de grupos sangüíneos e 4 sistemas de variantes protéicas, foi estimada
uma PE (Probabilidade de Exclusão de Paternidade) para as raças Apaloosa, Árabe, Quarto de Milha e
Puro Sangue Inglês, de 97,4%, 94,4%, 98,2% e 91,5 % respectivamente.
Vinocur et al. (2003) analisaram, a partir de sete sistemas de grupos sangüíneos e oito sistemas protéicos, a variabilidade genética de seis populações de cavalos Crioulos criados no sul do
Brasil. A heterozigosidade média observada dos 15 locos analisados juntamente foi de 0,4631 e foram
observados no total 87 alelos. A probabilidade de exclusão de um indivíduo indicado nos 15 locos
variou de 95,5% a 98,3%. Bowling e colaboradores (1997), considerando sete sistemas de grupos
sangüíneos e mais oito sistemas de variantes protéicas, calcularam a PE de 97,3% para a raça Quarto
de Milha. Embora estes testes têm sido eficientes para solucionar questões de paternidade e
maternidade resolvendo dúvidas de troca de produtos entre éguas ou paternidade quando mais de um
garanhão poderia ter gerado o produto, algumas limitações ocasionalmente geraram frustrações para
criadores e serviços de registro. A primeira é o fato dos exames dependerem da disponibilidade de
sangue fresco e devidamente conservado, excluindo assim a possibilidade de utilizar outros tipos de
amostras inclusive amostras forenses de animais falecidos. Em segundo lugar a coleta de sangue
requer muitas vezes a assistência de um veterinário e condições refrigeradas de manipulação e envio
quais dois garanhões geneticamente relacionados cobriram a mesma égua, a tipagem sangüínea
freqüentemente não permite a exclusão conclusiva de paternidade de um deles em função do limitado
poder de resolução do sistema. No caso específico de Puro Sangue Inglês a situação fica mais
complexa ainda uma vez que as estimativas de PE são sistematicamente as menores em comparação
às outras raças provavelmente como resultado da reduzida diversidade alélica oriunda do forte efeito
fundador que ocorreu na formação da raça.
Além disso, em função da natureza do material biológico analisado na tipagem sangüínea, há
casos em que esse metodologia não é suficiente, como no relato feito por Bowling et al. (1993b), em que uma égua portadora de quimerismo sangüíneo era excluída como mãe de um potro pelo exame de
tipagem sangüínea. Após testar o DNA extraído do sangue desta fêmea com cinco microssatélites
persistiu a exclusão. No entanto, ao testar-se o DNA extraído de bulbo capilar nenhuma exclusão foi
observada. Assim, verificou-se que a égua era uma quimera, possuindo duas linhagens celulares
distintas, sendo uma proveniente de seu irmão gêmeo (mostrada em seu sangue) e outra própria
(evidenciada nos bulbos capilares).
2.4 Marcadores microssatélites em eqüinos
Marcadores microssatélites são baseados na amplificação de seqüências de DNA compostas
por um número variável de nucleotídeos, normalmente de 1 a 7 pares de bases, repetidos várias vezes
e encontradas no genoma de eucariotos (Hamada et al., 1982; Tautz e Renz, 1984; Litt e Luty, 1989; Weber e May, 1989). Embora o polimorfismo presente nestas regiões tenha sido explorado inicialmente
Os microssatélites são abundantes e encontram-se regularmente distribuídos no genoma de
mamíferos (Tautz e Renz, 1984; Lagercrantz et al., 1993). Além disso, apresentam multialelismo e alto nível de polimorfismo resultante da alta incidência de escorregamento na replicação. Todas estas
características, aliadas ao padrão de segregação co-dominante, têm feito dos microssatélites o
marcador mais apropriado para o desenvolvimento de fingerprints genéticos únicos de espécies de plantas e animais.
Marcadores microssatélites são detectados através da técnica de reação de polimerase em
cadeia (PCR) com primers correspondentes a DNA de seqüência única que flanqueia as repetições em
tandem. Regiões contendo seqüências simples repetidas são amplificadas individualmente através de PCR utilizando-se um par de iniciadores específicos (primers) (de 20 a 30 bases) complementares às seqüências únicas que flanqueiam o microssatélite. Segmentos amplificados a partir destes sítios
quase que invariavelmente apresentam um polimorfismo extensivo resultante da presença de
diferentes números de elementos simples repetidos. Assim, cada microssatélite, independente do
elemento repetido, constitui um loco genético altamente variável, multialélico e de grande conteúdo
informativo. Cada segmento amplificado de tamanho diferente (geralmente de várias dezenas até
algumas centenas de pares de bases) representa um alelo diferente do mesmo loco.
A detecção de seqüências microssatélites via PCR é feita em gel de eletroforese utilizando-se
poliacrilamida ou agarose especial de alta resolução, uma vez que é necessário um gel adequado para
a separação de produtos de PCR que diferem por poucos pares de base, dependendo do número de
nucleotídeos do elemento repetido no microssatélite (Ferreira e Grattapaglia, 1998). A eletroforese para
a detecção destes marcadores moleculares deve ser realizada em padrões tais que permitam distinguir
bandas que diferem em apenas uma base e, por este motivo, são utilizadas condições de eletroforese
de alta resolução usadas para seqüenciamento de DNA.
Cada loco de microssatélite é analisado individualmente ao se utilizar o par de primers
analisado de cada vez quando os alelos de cada loco têm tamanhos suficientemente diferentes e
migram para zonas separadas no gel. Nestes métodos de genotipagem denominados "multiplex", mais
do que um par de primers específicos é utilizado simultaneamente na mesma reação de PCR (Ferreira e Grattapaglia, 1998).
Por possuírem distribuição abundante dentro do genoma, e terem um elevado grau de
polimorfismo, os marcadores microssatélites se tornaram uma ferramenta muito valiosa na genética de
eqüinos seja no mapeamento de genomas, estudos populacionais e na identificação individual e
estudos de parentesco. Em comparação com as populações humanas, entretanto, cavalos com
pedigree apresentam níveis mais elevados de homogeneidade genética. Isto é resultado principalmente
do forte efeito fundador principalmente em raças como o PSI bem como de acasalamentos
consangüíneos utilizados para manter o tipo e as características individuais das diferentes raças, bem
como a baixa freqüência ou mesmo ausência de fluxo gênico entre raças devido às barreiras impostas
pelos sistemas de pedigree. Mesmo nestas condições, devido à ampla hipervariabilidade, marcadores
microssatélites têm sido úteis para o mapeamento genético de eqüinos (Shiue et al., 1999; Guérin et al., 2003), verificação da extensão do desequilíbrio de ligação (DL) (Tozaki et al., 2005), estudos de evolução, caracterização e diferenciação entre populações e raças (Kruger et al., 2003; Aberle et al., 2004; Achmann et al., 2004; Bjørnstad e Røed, 2001; Zabek et al., 2005), e aplicações na verificação de parentesco (Binns et al., 1995; Bowling et al., 1997; Tozaki et al., 2001a; Lee e Cho, 2006).
Nos últimos 12 anos, com o avanço da automação e reagentes para análise molecular, o uso
de marcadores moleculares baseados em microssatélites para fins de genotipagem individual evoluiu
para o desenvolvimento de sistemas de análise multiloco semi-automatizada (Levitt et al., 1994; Kimpton et al., 1993). Estes sistemas foram desenvolvidos inicialmente para estudos de genética humana (Fregeau et al., 1993; Gill et al., 1995). Eles baseiam-se na amplificação via PCR de um ou mais locos microssatélites, simultaneamente, utilizando iniciadores marcados com fluorocromos, e
Recentemente, sistemas semelhantes foram desenvolvidos para análise de espécies animais
de importância econômica. O objetivo tem sido estabelecer sistemas universais e altamente robustos
de identificação genética para fins de exames de vínculo genético, estudos de variabilidade em
populações e estimativas de distância genética. Entre as vantagens destes sistemas estão: (1) rapidez
e acurácia na geração de dados; (2) avaliação simultânea de vários locos microssatélites em uma única
pista do gel, reduzindo o número de análises necessárias em relação às técnicas convencionais; (3)
aquisição e armazenamento dos dados (computadorizados) em tempo real, reduzindo a manipulação
manual da informação; (4) possibilidade de exportação direta dos dados para programas de análise
genética e (5) transferibilidade imediata do sistema para outros laboratórios, permitindo a padronização
dos locos do genoma amostrados e a comparação e compartilhamento de resultados.
No caso de eqüinos, várias centenas de microssatélites foram desenvolvidos por diversos
grupos de pesquisa no mundo e publicados ao longo dos últimos 12 anos (ex. Ellegren et al., 1992; Marklund et al., 1994; van Haeringen et al., 1994; 1998; Guérin et al., 1994; Binns et al., 1995; Mickelson et al., 2003; Tozaki et al., 2000a, 2000b, 2001b, 2004; Wagner et al., 2004). A primeira geração de mapas de ligação de eqüinos foi publicada em 2000 (Swinburne et al., 2000a) com 31 grupos de ligação utilizando uma família de irmãos completos. Desde então, outros mapas de ligação
de eqüinos usando famílias de meios irmãos(Guérin et al., 2003) e híbridos de radiação (Chowdhary et al., 2003) vem sendo publicados.Os mapas de ligação têm sido usados para identificar a base genética de características hereditárias associadas com o desempenho e saúde do animal. Tozaki e
colaboradores (2005) utilizaram a característica da cor da cobertura de cavalos como um modelo para
verificar a extensão do desequilíbrio de ligação, chegando a uma janela de 7cM. As informações sobre
microssatélites mapeados, as seqüências dos seus primers e as respectivas características dos locos
foram organizadas em um banco de dados mantido pelo Roslin Institute no endereço
http://www.thearkdb.org/browser?species=horse. Atualmente este banco conta com 1470 locos dos
grande maioria microssatélites mapeados utilizando mapeamento em uma coleção de linhagens
celulares de híbridos de radiação. Recentemente, consolidando dados de quatro fontes de segregação
de marcadores foi publicado um mapa mais atualizado com 766 marcadores distribuídos pelos 31
cromossomos cobrindo 3.740 cM com uma distância média entre marcadores de 6,3 cM (Penedo et al. 2005).
2.5 Marcadores microssatélites para identificação e parentesco em eqüinos
Com a evolução das tecnologias do DNA recombinante, o descobrimento de seqüência simples
repetitivas no genoma de eucariotos e o desenvolvimento da PCR, a partir de meados da década de 90
foi iniciada a transição de tecnologia de verificação de parentesco em eqüinos, passando de tipagem
sangüínea para a tipagem de marcadores moleculares baseados na variação observada em
seqüências de DNA. O trabalho de Bowling et al. (1993a) é o pioneiro descrevendo, durante um congresso mundial de identificação humana, a introdução da tipagem de DNA para a verificação de
parentesco em bovinos e eqüinos. Em seguida Binns et al. (1995) utilizaram uma bateria de 8 marcadores microssatélites em cavalos PSI. Eles analisaram oito casos de dupla cobertura, ou seja,
com dois supostos garanhões, que não puderam ser resolvidos pela tipagem sangüínea e em cinco dos
oito casos eles identificaram exclusões claras de paternidade. Eles concluem o trabalho afirmando que
a análise de DNA é um adjunto útil à tipagem sangüínea e que, provavelmente no futuro, iria
substituí-la. Hoje, 2007, a transição de tipagem sangüínea para tipagem de microssatélites ainda está de certa
forma ocorrendo uma vez que muitos laboratórios e associações possuem um grande número de
animais genotipados via tipagem sangüínea e ficam, portanto, resistentes a mudar de tecnologia e com
isso ter que re-genotipar todos estes animais. Entretanto esta mudança tecnológica é irreversível
principalmente devido à pressão do comércio mundial de animais com destaque para animais Puro
Várias são as vantagens da análise de marcadores microssatélites. Com a análise de DNA,
não são necessárias amostras de sangue fresco, bastando sangue imobilizado sobre papel filtro e sem
necessidade de refrigeração. Alternativamente podem ainda ser usados bulbos capilares de pêlos de
crina ou mesmo tecidos e ossos nos casos de animais falecidos. Além destas vantagens práticas,
marcadores de DNA fornecem um poder mais elevado de discriminação individual e de exclusão de
paternidade e maternidade mesmo em casos de forte consangüinidade. A análise de DNA permite
ainda a utilização de um grande número de marcadores adicionais para solucionar casos complexos de
verificação de parentesco nos casos das baterias padrão de marcadores não conseguirem fornecer um
resultado conclusivo.
Baterias de marcadores microssatélites para eqüinos têm sido propostas e otimizadas em
alguns sistemas multiplex nos últimos anos visando atender situações de dificuldade de discriminação
com base em tipagem sangüínea principalmente em situações nas quais garanhões aparentados são
testados frente a um mesmo produto na qualidade de supostos pais. Bowling et al. (1997) validaram uma bateria de 11 microssatélites que inclui os nove recomendados pela ISAG. Para avaliar esta
bateria de marcadores, eles compararam o desempenho forense destes marcadores com 15 locos de
grupos sangüíneos e polimorfismos protéicos em 4803 cavalos da raça Quarto de Milha que, embora
endogâmica, apresenta um histórico de base genética mais ampla do que cavalos PSI. Eles relataram
que os 26 locos juntamente apresentaram uma efetividade teórica de 99,999% na detecção de
paternidade incorreta. Apesar de relativamente poucos locos, a tipagem de DNA foi mais efetiva que a
tipagem sangüínea alcançando um PE de 99,9%.
Alguns trabalhos otimizaram baterias de marcadores microssatélites para a genotipagem de
eqüinos em sistemas multiplex nos últimos anos. Marklund et al. (1994) otimizaram uma bateria de 8 marcadores microssatélites (HTG3, 4, 6, 8, 10, 12, 14 e 15) para a genotipagem de quatro raças de eqüinos em sistemas multiplex. O PE combinado estimado para esse conjunto de marcadores foi de
Tozaki et al. (2001a) propuseram uma nova bateria de microssatélites desenvolvida no Japão a partir de um amplo trabalho de desenvolvimento de novos microssatélites para eqüinos, batizados com
o código TKY. Uma bateria de 15 marcadores TKY aplicada em 250 eqüinos não relacionados da raça
PSI, forneceu um poder combinado de exclusão de 99,998%, significativamente superior à bateria hoje
em utilização recomendada pela ISAG. Lee e Cho (2006) utilizaram uma bateria de 14 microssatélites
que inclui os nove marcadores recomendados pela ISAG e mais 5 adicionais comumente utilizados
internacionalmente (ASB17, ASB23, CA425, HMS1, LEX33), atingindo uma PE de 99,98% com base
em uma amostra de 1285 cavalos PSI. Jakabová et al. (2002) avaliaram o poder individual e
combinado de seis microssatélites (ASB2, HMS3, HMS6, HMS7, HTG4, VHL20) em uma amostra de
352 cavalos PSI na República Eslovaca e estimaram um poder combinado de exclusão de 98,88%. Foi
observado que com apenas cinco marcadores microssatélites com maior PE individual, foi alcançada
uma probabilidade de exclusão combinada de 98,45%. Luis et al. (2002) utilizaram seis microssatélites (ASB2, HMS3, HMS7, HTG4, HTG10, e VHL20) em três raças portuguesas autóctones de cavalos alcançando um poder de exclusão combinado de 99,6%, 99,5% e 88,5% para as raças Lusitano,
Garrano e Sorraia, respectivamente. Esta última raça é caracterizada por um histórico de forte
endocruzamento.
Dimsoski (2003) otimizou uma bateria de 17 microssatélites que inclui os nove recomendados
pela ISAG (VHL20, HTG4, AHT4, HMS7, HMS6, HMS3, AHT5, ASB2 e HTG10) e mais oito
marcadores também freqüentemente usados (ASB17, LEX3, HMS1, CA425, HTG7, HTG6, HMS2 e ASB23), constituindo um kit comercializado pela Applied Biosystems. Embora este trabalho não especifique o poder de exclusão atingido pela bateria de marcadores, certamente deverá estar acima