1 Corresponding author:
Received: May 15, 2017.
Accepted : October 09, 2017.
Copyright: All the contents of this
journal, except where otherwise noted, is licensed under a Creative Commons Attribution License.
Promoting fruit seedling growth
by encapsulated microorganisms
Carlos Henrique Barbosa Santos
1, Antonio Baldo Geraldo Martins
2,
Everlon Cid Rigobelo
2, Gustavo Henrique de Almeida Teixeira
2Abstract– The use of microorganisms capable of promoting plant growth has been accepted as an alternative to reducing the use of chemical fertilizers. The aim of this study was to evaluate the inoculation of plant growth promoting microorganisms in seedlings of fruit species, verifying the interaction of the inoculums with encapsulating agents such as clay and alginate. Microbial
inoculums contained the following species: Azospirillum brasilense, Burkolderia cepacia, Bacillus
thuringienses, Bacillus megaterium, Bacillus cereus, Bacillus subtilis, Tricoderma spp. and isolate
411. The fruit species evaluated were: Myrciaria cauliflora (DC.) O. Berg; Myrciaria glazioviana
(Kiaersk.) G. Barros & Sobral; Myrciaria dubia (Kunth) Mc Vaugh; Eugenia brasiliensis Lam.;
Diospyroskaki L.; Garcinia brasiliensis Mart.; Annona muricata L.; Duguetia lanceolata A.
St.-Hil.; Chrysophyllum cainito L.; Anacardium occidentale L.; Eriobotrya japonica (Thunb.) Lindl.
and Litchi chinensis Sonn. The experimental design was completely randomized, in a factorial scheme 3 (control, sodium alginate and clay) x 2 (presence and absence of microbial inoculum) with
five replicates (one seedling per replicate). Seedlings were maintained in 50% of illumination at
an average temperature of 22.5 °C for ninety days, and plant height, diameter, root and shoot dry mass were evaluated. Plant growth promoting microorganisms, regardless of encapsulation used,
promote higher development of C.cainito and L. chinensis seedlings.
Index terms: Fruit plants, development, encapsulation, plant-growth promoting agents.
Promoção do crescimento de mudas frutíferas
por meio de microrganismos encapsulados
Resumo – A utilização de microrganismos capazes de promover o crescimento vegetal tem sido aceita como alternativa à redução do uso de adubos químicos. O objetivo deste trabalho foi avaliar a inoculação de microrganismos promotores do crescimento de plantas em substrato
de mudas de espécies frutíferas, verificando a interação do inóculo com agentes encapsulantes: argila e alginato. O inóculo microbiano continha as seguintes espécies: Azospirillum brasilense, Burkolderia cepacia, Bacillus thuringienses, Bacillus megaterium, Bacillus cereus, Bacillus subtilis, Tricoderma spp. e isolado 411. As espécies frutíferas avaliadas foram: Myrciaria
cauliflora (DC.) O. Berg (jabuticabeira); Myrciaria glazioviana (Kiaersk.) G. Barros & Sobral
(cabeludinha); Myrciaria dubia (Kunth) Mc Vaugh (camu-camu); Eugenia brasiliensis Lam.
(grumixama); Diospyros kaki L. (caqui); Garcinia brasiliensis Mart. (bacupari); Annona
muricata L. (graviola); Duguetia lanceolata A. St. – Hil. (pindaíba); Chrysophyllum cainito
L. (caimito); Anacardium occidentale L. (caju); Eriobotrya japonica (Thunb.) Lindl. (nêspera)
e Litchi chinensis Sonn. (lichia). O delineamento experimental foi o inteiramente casualizado
(DIC), em esquema fatorial 3 (controle, alginato de sódio e argila) x 2 (presença e ausência de inóculo microbiano), com cinco repetições (uma muda por repetição). As mudas foram mantidas em 50% de iluminação, à temperatura média de 22,5 °C, durante noventa dias, sendo avaliados
a altura e o diâmetro do colo das plantas, e a massa seca da parte aérea e das raízes. Micro-organismos promotores de crescimento de plantas, independentemente do encapsulamento utilizado, promovem desenvolvimento superior de mudas de caimito e de lichia.
Termos para indexação: Plantas frutíferas, desenvolvimento, encapsulamento, promotores de crescimento vegetal.
1Agronomist. PhD. in Agronomy (Plant Production). São Paulo State University (UNESP), School of Agricultural and Veterinarian Sciences, Campus of Jaboticabal. Email: [email protected]
2
C. H. B. Santos et al.
Rev. Bras. Frutic., Jaboticabal, 2018, v. 40, n. 3: (e-179)
Introduction
The production of fruit seedlings is influenced
by abiotic factors, such as light, temperature, gas exchange, water, moisture, substrate and nutrition, and biotic factors, such as pathogens, pests and rhizosphere microorganisms (HARTMANN et al., 2010).
In relation to the substrate, it must be dense and
firm, highly decomposed and stable, sufficiently porous
to drain excess water, free of pests, diseases and weeds, present low salinity, pasteurized or chemically treated, have high cation exchange capacity (CEC), nutrient retention and easy availability (HARTMANN et al., 2010). In addition, biotic factors, such as rhizosphere microorganisms (mycorrhizal bacteria and fungi), can
influence the development of seedlings, and plant growth promoting bacteria (PGPB) exert beneficial effects by stimulating, for example, nitrogen fixation (BEUTLER
et al., 2016), nutrient solubilization (MAJEED et al., 2015), production of plant regulators (FAHAD et al., 2015) and increase of root absorption capacity (PII et al., 2006).
Indirectly, PGPB acts as biological control agents against pathogens present in the substrate, possibly through the production of hydrocyanic acid, bactericides
and antibiotics, by space competition, uptake of Fe+3
and other nutrients, parasitism, resistance induction and cross protection (MOTA et al., 2017).
In relation to PGPB species, the genus
Azospirillum has several species that grow widely in the rhizosphere of grasses and, after being isolated and used in plant production, presented positive responses
on productivity and on biological nitrogen fixation
(BNF) (HUNGRIA et al., 2015; MÜLLER et al., 2016;
PEREG et al., 2016). Burkholderia cepacia is another
important species of bacteria in the BNF process (GÓES
et al., 2015). Bacillus isolates have been shown to have
satisfactory effects when used as biocontrol agents and
to promote plant growth, highlighting Bacillus subtilis
and Bacillus cereus isolates (SOUZA et al., 2017),
Bacillus thuringienses (CONSTANSKI et al., 2015) and Bacillus megaterium (ZHOU et al., 2016). Among
fungi, Trichoderma spp. stands out as a biocontrol agent
(CONTRERAS-CORNEJO et al., 2016) and also in plant growth promotion ((NIETO-JACOBO et al., 2017).
One way of incorporating PGPB into substrates is to encapsulate solid, liquid and gaseous materials into small capsules that release their contents under controlled conditions. Encapsulation is important for maintaining the stability of microorganisms and for increasing the resistance of bacteria in the substrate. Among the many types of encapsulating materials, calcium and sodium alginate, as well as whey proteins and gums have been the most widely used (SCHOEBITZ et al., 2013; VEMMER; PATEL, 2013). Clay is also a good encapsulating agent
with properties different from alginate, but maintaining the desired properties of an encapsulating agent. The use of clay for the preservation of conidia has demonstrated satisfactory effect in storage, guaranteeing the viability of this for a long period (SHI, 1988).
The isolated or combined inoculation of microorganisms may constitute a biotechnological tool of interest in the production of fruit plants. In this context, the aim of this study was to evaluate the effect of inoculation of microorganisms (PGPB) on substrate used for seedlings of fruit species, verifying the interaction of the inoculum with encapsulating agents clay and sodium alginate.
Material And Methods
The experiment was conducted between March
16 and June 16, 2015 (90 days) in the Fruit field of the
School of Agricultural and Veterinarian Sciences, São Paulo State University (UNESP), Campus of Jaboticabal, located in the following geographical coordinates (lat 21º14’05’’S, long 48º17’09’’W, 590 m above sea level). The climate of the region, according to the Köppen
classification is Cwa, with average annual temperature
of 22 ºC.
The inoculum used contained the following species
of microorganisms: Azospirillum brasilense, Burkolderia
cepacia, Bacillus thuringienses, Bacillus megaterium, Bacillus cereus, Bacillus subtilis, Tricoderma spp.
and isolate identified as 411. All microorganisms are
preserved in the collection of the Laboratory of Soil Microbiology of the Department of Plant Production - UNESP, Campus of Jaboticabal.
Microorganisms were separately cultured in flasks
containing nutritive broth for seven days and incubated in B.O.D. oven (Biochemical Oxygen Demand) (Eletrolab, model 347 F, Brazil), at 25 ºC. After the incubation period, microorganisms were separately centrifuged at 10,000 rpm for 10 min at 28°C (Novatecnica, model MLW K24, Brazil). The inoculum concentration was standardized as recommended by Barry and Thornsberry
(1991) and Sahm and Washington II (1991) in 1 x 107
CFU mL-1, using a spectrophotometer (Micronal, model
B382, Brazil) at absorbance at 695 nm.
To each microorganism solution, sodium alginate
solution (NaC6H7O6) at concentration of 1% (w/v) was
added. The microorganism solutions added of sodium alginate were dripped in 0.1M calcium chloride solution
(CaCl2), giving rise to beads with the microbial inoculum
(SHEU; MARSHALL, 1993; SULTANA et al., 2000). Beads were then washed with distilled water using a sieve
and submitted to drying in a laminar flow chamber for a
applied in each pot, each seedling received 96 microbial beads, which were incorporated into the substrate in a hole at approximately 10 cm in depth in order to guarantee a greater proximity of the material deposited in the substrate with the roots of seedlings. Treatment containing alginate received only sodium alginate beads without the presence of the microbial inoculum, and 96 beads per pot were added.
In order to inoculate microorganisms with clay, suspension was prepared containing 1 mL of each microorganism was prepared for 1 mL of clay in the
dilution of 1: 3 (w/v) distilled water (adapted from
CARNEIRO, GOMES, 1997). Each pot received a total of 8 mL of microbial inoculum added to 8 mL of clay, which were incorporated into the substrate in hole of approximately 10 cm in depth, totaling 16 mL per pot. For treatment containing only clay, the same procedure was performed, excluding only the addition of the microbial inoculum.
All seedlings of fruit species studied were originated from seeds obtained from mother plants belonging to the Germplasm Active Bank of UNESP, Campus de Jaboticabal, conducted in one plant per pot. Microorganisms were inoculated on substrates where seedlings of the following fruit species were
being conducted: Myrciaria cauliflora (DC.) O. Berg
(jabuticaba); Myrciaria glazioviana (Kiaersk.) G. Barros &
Sobral (cabeludinha); Myrciaria dubia (Kunth) Mc Vaugh
(camu-camu); Eugenia brasiliensis Lam. (grumixama);
Diospyros kaki L. (Persimmon); Garcinia brasiliensis
Mart. (bacupari); Annona muricata L. (graviola); Duguetia
lanceolata A. St.-Hil. (pindaíba); Chrysophyllum cainito
L. (caimito); Anacardium occidentale L. (cashew);
Eriobotrya japonica (Thunb.) Lindl. (medlar) and Litchi chinensis Sonn. (lychee).
The substrate used was composed of a 3: 1:
1 (v/v/v) mixture of soil, sand and bovine manure,
disposed in polyethylene bags (22 x 11 cm). Irrigation was manual, daily, according to substrate moisture
monitoring. Seedlings were kept under a 50% luminosity
cover for a period of 90 days. During the experiment, the minimum temperature was 17 °C and the maximum temperature was 28 °C.
The seedlings selected for this experiment were standardized according to stem height and diameter before inoculation. At the end of 90 days, they were evaluated as to plant height (cm), measuring from the plant base to the apex; stem diameter (mm), in the neck region with the aid of a digital caliper; shoot dry mass (g) and roots dry mass (g) (65 °C, 72 h), as described by Araújo Neto et al. (2015).
Due to the different growth patterns of each fruit species, statistical analysis comparing the results among species was not performed. Thus, for each fruit species, completely randomized design was used in a
factorial scheme 3 (control, sodium alginate and clay) x 2 (presence and absence of microbial inoculum) with
five replications (one repetition), totaling 30 plants.
Data were submitted to analysis of variance (ANOVA)
and means compared by the Tukey test (p≤0.05), using
AgroEstat Software (BARBOSA; MALDONADO JÚNIOR, 2009).
Results and Discussion
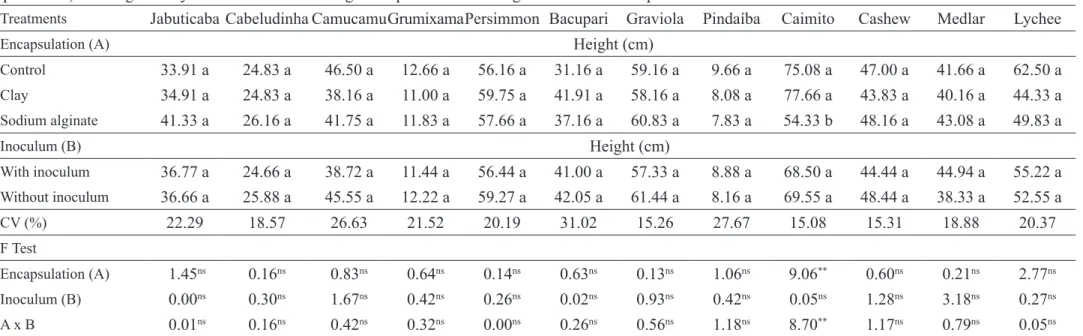
Significant effect (P<0.01) of the growth
promoter microorganisms was observed only for C.
cainito L. (caimito) plant height, both for encapsulation in sodium alginate and for microorganisms used in the
inocula (Table 1). For the other species, no significant
differences were observed for plant height, indicating
that the microorganisms did not influence the growth of
seedlings, regardless of type of encapsulation (clay or sodium alginate).
It was reported by Spassin et al. (2016) that
bacterial isolates may have beneficial or no beneficial
effects in relation to the survival and rooting of Eucalyptus
dunnii (Myrtaceae), which raises the hypothesis that the establishment of growth-promoting microorganisms and their association with the plant species may vary according to species, management and, possibly, the place of study. However, Salla et al. (2014) concluded
that the use of PGPB (Streptomyces PM9 rhizobacteria)
resulted in higher yield of indoleacetic acid and a better potential for root induction, as well as in the levels of
phenolic compounds and flavonoids involved in the
defense mechanism of Eucalyptus grandis and E.
globulus plants. The authors also reported alteration in secondary metabolism, suggesting an induced systemic response, indicating their potential use as a biological control in forestry activities.
When analyzing the height variable for the caimito species, it was observed that the encapsulation with clay yielded better results compared to sodium alginate encapsulation, since it did not differ statistically from the control (Table 2). Lower growth of untreated (control) seedlings when compared to growth in the other treatments was also observed (Table 2), suggesting that the use of microorganisms promoted the development of caimito seedlings. Thus, it could be inferred that the associations of bacterial communities associated with the rhizosphere of fruit species are hosts dependent on the taxon, that is, on the species itself, as concluded by Lambais et al. (2014), who studied the rhizosphere of arboreal species of the Atlantic Forest and showed that the bacterial communities associated to trees were
specific to the hosts of plants.
4
C. H. B. Santos et al.
Rev. Bras. Frutic., Jaboticabal, 2018, v. 40, n. 3: (e-179)
Vaugh (camu-camu) seedlings. For this species, the stem diameter was higher when growth promoting microorganisms were not used, either encapsulated or not (Table 3). This can occur when the microorganisms used present some incompatibility with the native microbiota associated with the plant rhizosphere, which will generate competition between them, altering the positive effects of the target microorganism and, consequently, an unsuitable development of seedlings. Another possible result is due to variable root colonization due to inoculum survival problems or to unfavorable conditions for microorganisms, since the rhizosphere colonization depends on a set of abiotic and biotic factors, including plant genotypes and microorganisms, environmental conditions and the dynamic network of plant interactions
(HARDOIM et al., 2015). However, for C. cainito
(caimito) and L. chinensis (lychee) species, the opposite
occurred, that is, larger stem diameter was observed in both species when seedlings were inoculated with
microorganisms (Table 3). This result confirms the result
observed for plant height variable, in which changes in plant growth may be related to the interaction between microorganisms and rhizosphere.
One possible explanation for this is that soil microorganisms found in the root zone affect plant growth and development, but the potential to take
advantage of these benefits depends on the abundance and diversity of individuals influencing the beneficial
effects on the plant (PANKE-BUISSE et al., 2015). In addition, the same plant species can stimulate diverse microbial communities from one site to another, which provides different responses to the inoculation of a single isolated growth promoter (XU et al., 2016). Additionally, genetic variability in seed propagated seedlings may be considered a factor of plant growth differentiation (HARTMANN et al., 2010).
Lychee seedlings presented significant increases (P<0.05) in the dry mass (DM) content of the plant
shoots with the presence of microorganisms, regardless of whether they were encapsulated or not (Table 4). This result suggests that there may have been an association with the rhizosphere of lychee seedlings, which promoted better establishment of microorganisms and, consequently, provided larger stem diameter, possibly as a function of the higher DM content.
Root dry mass was significantly different only
in caimito seedlings treated with growth promoting microorganisms (Table 5). In this species, root DM accumulation was higher when the substrate was inoculated with microorganisms, regardless of type of encapsulant used.
When evaluating eleven isolates of the genus
Acremonium (GFR6 and GRR1), Colletotrichum
(GFR4 and PFR4), Phomopsis (PFR3 and GCR4),
Cylindrocladium (GRR4), Chaetomium (GRR7) and
Fusarium (GRR5, PRR1 and PRR6), Silva et al. (2006) reported effects of these microorganisms on growth
the parameters of Annona squamosa L. and Annona
muricata L. seedlings with an increase in the shoot
dry matter content of Annona squamosa L. seedlings
between 23.2 and 32.7%, but no isolate affected the root
dry mass content, similar to that observed in this study. Silva et al. (2006) also reported that 20 isolates had a negative effect on the root dry matter content of
Annona squamosa L. seedlings, and observed that in
apparently healthy Annona squamosa L. and Annona
muricata L. tissues, some fungi that could promote the growth of shoots were found, but others could reduce root growth and others that would not affect the growth of seedlings. Another interesting observation is that isolates that promoted the growth of sugar apple seedlings came from graviola, evidencing that isolates from one host can easily colonize hosts of different species, even with greater intensity (Silva et al., 2006). These characteristics
may be associated to the efficiency of root colonization
5
Promoting fruit seedling growth by encapsulated microor
ganisms
Rev
.
Bras.
Frutic.,
Jaboticabal,
2018,
v.
40,
n.
3:
(e-179)
persimmon (Diospyros kaki) (Ebenaceae); bacupari (Garcinia brasiliensis) (Clusiaceae); graviola (Annona muricata), pindaiba (Duguetia lanceolata) (Annonaceae); caimito (Chrysophyllum cainito) (Sapotaceae); cashew (Anacardium occidentale) (Anacardiaceae); medlar (Eriobotrya japonica) (Rosaceae) and lychee (Litchi chinensis) (Sapindaceae) seedlings 90 days after inoculation with growth promoter microorganisms in two encapsulation methods.
Treatments Jabuticaba Cabeludinha CamucamuGrumixamaPersimmon Bacupari Graviola Pindaíba Caimito Cashew Medlar Lychee
Encapsulation (A) Height (cm)
Control 33.91 a 24.83 a 46.50 a 12.66 a 56.16 a 31.16 a 59.16 a 9.66 a 75.08 a 47.00 a 41.66 a 62.50 a
Clay 34.91 a 24.83 a 38.16 a 11.00 a 59.75 a 41.91 a 58.16 a 8.08 a 77.66 a 43.83 a 40.16 a 44.33 a
Sodium alginate 41.33 a 26.16 a 41.75 a 11.83 a 57.66 a 37.16 a 60.83 a 7.83 a 54.33 b 48.16 a 43.08 a 49.83 a
Inoculum (B) Height (cm)
With inoculum 36.77 a 24.66 a 38.72 a 11.44 a 56.44 a 41.00 a 57.33 a 8.88 a 68.50 a 44.44 a 44.94 a 55.22 a
Without inoculum 36.66 a 25.88 a 45.55 a 12.22 a 59.27 a 42.05 a 61.44 a 8.16 a 69.55 a 48.44 a 38.33 a 52.55 a
CV (%) 22.29 18.57 26.63 21.52 20.19 31.02 15.26 27.67 15.08 15.31 18.88 20.37
F Test
Encapsulation (A) 1.45ns 0.16ns 0.83ns 0.64ns 0.14ns 0.63ns 0.13ns 1.06ns 9.06** 0.60ns 0.21ns 2.77ns
Inoculum (B) 0.00ns 0.30ns 1.67ns 0.42ns 0.26ns 0.02ns 0.93ns 0.42ns 0.05ns 1.28ns 3.18ns 0.27ns
A x B 0.01ns 0.16ns 0.42ns 0.32ns 0.00ns 0.26ns 0.56ns 1.18ns 8.70** 1.17ns 0.79ns 0.05ns
(1)Means followed by the same letter in the column do not differ by the Tukey test, * significant at 5% probability, ** significant at 1% probability and ns not significant.
Table 2. Behavior of height variable of caimito (Chrysophyllum cainito) seedlings (cm) 90 days after inoculation with growth-promoting microorganisms as a function of encapsulation.
Encapsulation (A)
Inoculum (B)
Absence (B1) Presence (B2)
Plant height (cm)
Control (A1) 65.33 aB 84.83 aA
Clay (A2) 74.50 aA 80.83 aA
Sodium alginate (A3) 68.83 aA 39.83 bA
6
C. H. B. Santos et al.
Rev
. Bras. Frutic., Jaboticabal, 2018, v
. 40, n. 3: (e-179)
Table 3. Stem diameter (DIAM) of jabuticabra (Myrciaria cauliflora), cabeludinha (Myrciaria glazioviana), camu-camu (Myrciaria dubia), grumixama (Eugenia brasiliensis) (Myrtaceae); persimmon (Diospyros kaki) (Ebenaceae); bacupari (Garcinia brasiliensis) (Clusiaceae); graviola (Annona muricata), pindaíba (Duguetia lanceolata) (Annonaceae); caimito (Chrysophyllum cainito) (Sapotaceae); cashew (Anacardium occidentale) (Anacardiaceae); medlar (Eriobotrya japonica) (Rosaceae) and lychee (Litchi chinensis) (Sapindaceae), 90 days after inoculation with growth promoting microorganisms in two encapsulation methods.
Treatments Jabuticaba Cabeludinha Camucamu Grumixama Persimmon Bacupari Graviola Pindaíba Caimito Cashew Medlar Lychee
Encapsulation (A) DIAM (mm)
Control 7.12 a 2.90 a 3.83 a 2.13 a 6.85 a 6.65 a 9.61 a 2.58 a 9.21 a 7.76 a 4.22 a 9.37 a
Clay 7.00 a 2.93 a 3.60 a 2.02 a 5.93 a 6.24 a 8.69 a 2.18 a 9.07 a 6.98 a 3.54 a 8.68 a
Sodium alginate 7.22 a 2.50 a 3.50 a 1.96 a 7.43 a 7.07 a 8.51 a 2.31 a 7.31 a 7.77 a 4.05 a 8.31 a
Inoculum (B) DIAM (mm)
With inoculum 7.14 a 2.84 a 3.32 b 2.07 a 6.77 a 6.31 a 9.00 a 2.37 a 9.27 a 7.24 a 3.92 a 9.64 a
Without inoculum 7. 10 a 2.71 a 3.97 a 2.00 a 6.70 a 6.99 a 8.87 a 2.35 a 7.79 b 7.77 a 3.93 a 7.93 b
CV (%) 11.69 19.19 12.14 17.43 15.23 36.12 10.59 22.49 16.68 14.28 16.32 17.82
F Test
Encapsulation (A) 0.11ns 1.25ns 0.84ns 0.35ns 3.24ns 0.18ns 2.33ns 0.89ns 3.33ns 1.08ns 1.83ns 0.71ns
Inoculum (B) 0.01ns 0.28ns 9.55** 0.19ns 0.02ns 0.36ns 0.08ns 0.00ns 4.89* 1.10ns 0.00ns 5.38*
A x B 2.02ns 0.50ns 0.11ns 0.35ns 1.05ns 0.21ns 2.85ns 0.94ns 0.30ns 0.90ns 0.22ns 0.16ns
(1)Means followed by the same letter in the column do not differ by the Tukey test, being * significant at 5% probability, ** significant at 1% probability and ns not significant.
Table 4. Shoot dry matter (SDM) of jabuticaba (Myrciaria cauliflora), cabeludinha (Myrciaria glazioviana), camu-camu (Myrciaria dubia), grumixama (Eugenia brasiliensis) (Myrtaceae); persimmon (Diospyros kaki) (Ebenaceae); bacupari (Garcinia brasiliensis) (Clusiaceae); graviola (Annona muricata), pindaíba (Duguetia lanceolata) (Annonaceae); caimito (Chrysophyllum cainito) (Sapotaceae); cashew (Anacardium occidentale) (Anacardiaceae); medlar (Eriobotrya japonica) (Rosaceae) and lychee (Litchi chinensis) (Sapindaceae), 90 days after inoculation with growth promoting microorganisms in two encapsulation methods
Treatments Jabuticaba Cabeludinha Camucamu Grumixama Persimmon Bacupari Graviola Pindaíba Caimito Cashew Medlar Lychee
Encapsulation (A) SDM (g)
Control 9.79 a 4.83 a 6.34 a 4.40 a 12.04 a 21.18 a 11.92 a 4.89 a 33.52 a 12.04 a 15.42 a 10.86 a
Clay 9.26 a 5.14 a 4.61 a 4.62 a 11.49 a 18.16 a 12.19 a 4.62 a 34.60 a 11.49 a 12.05 a 11.59 a
Sodium alginate 12.35 a 6.66 a 4.59 a 4.89 a 10.47 a 22.21 a 14.23 a 4.89 a 23.95 a 10.12 a 13.36 a 11.69 a
Inoculum (B) SDM (g)
With Inoculum 10.35 a 5.14 a 4.60 a 4.67 a 11.70 a 16.97 a 11.53 a 4.71 a 35.51 a 11.46 a 14.85 a 12.22 a
Wthout Inoculum 10.58 a 5.95 a 5.76 a 4.60 a 10.97 a 24.97 a 14.04 a 4.89 a 25.88 a 10.97 a 12.38 a 10.54 b
CV (%) 32.50 29.17 31.91 9.46 15.81 75.66 28.17 10.25 40.53 16,17 30.25 13.80
F Test
Encapsulation (A) 1.41ns 0.21ns 2.21ns 1.86ns 1.18ns 0.11ns 0.74ns 0.61ns 1.33ns 1.79ns 1.02ns 0.50ns
Inoculum (B) 0.02ns 1.12ns 2.19ns 0.13ns 0.74ns 0.94ns 2.17ns 0.56ns 2.70ns 0.33ns 1.62ns 5.15*
A x B 0.30ns 0.74ns 0.32ns 0.67ns 0.17ns 0.50ns 3.37ns 1.06ns 0.70ns 0.46ns 1.64ns 0.88ns
T
able 5
.
Root dry matter (RDM) of jabuticaba
(
Myr
ciaria
cauliflora
), cabeludinha (
Myr
ciaria glazioviana
), camu-camu (
Myr
ciaria dubia
), grumixama (
Eugenia brasiliensis
)
(Myrtaceae); persimmon (
Diospyr
os kaki
) (Ebenaceae); bacupari (
Gar
cinia brasiliensis
) (Clusiaceae); graviola (
Annona muricata
), pindaíba (
Duguetia lanceolata ) (Annonaceae); caimito ( Chrysophyllum cainito ) (Sapotace ae); cashew ( Anacar dium occidentale ) (Anacardiaceae); medlar ( Eriobotrya japonica ) (Rosaceae) and lychee ( Litchi chinensis
) (Sapindaceae), 90 days after inoculation with growth promoting microor
ganisms in two encapsulation methods
T ratamentos Jabuticaba Cabeludinha Camucamu Grumixama Persimmon Bacupari Graviola Pindaíba Caimito Cashew Medlar L ychee Encapsulation (A) RDM (g) Control 6.03 a 1.26 a 2.97 a 1.31 a 5.85 a 7.02 a 8.18 a 1.18 a 10.54 a 1.97 a 3.17 a 3.33 a Clay 5.25 a 1.33 a 2.09 a 1.25 a 5.82 a 5.53 a 6.82 a 1.23 a 1 1.09 a 1.95 a 2.47 a 3.62 a Sodium alginate 7.28 a 1.86 a 1.74 a 1.08 a 5.77 a 6.57 a 5.95 a 1.01 a 6.85 a 2.02 a 2.49 a 3.16 a Inoculum (B) RDM (g) W ith Inoculum 5.97 a 1.44 a 2.01 a 1.25 a 5.91 a 6.07 a 6.98 a 1.1 1 a 1 1.76 a 1.92 a 2.79 a 3.74 a Wthout Inoculum 6.41 a 1.53 a 2.40 a 1.18 a 5.72 a 6.68 a 6.99 a 1.17 a 7.22 b 2.04 a 2.63 a 2.99 a CV (%) 23.26 39.65 32.86 16.24 7.98 68.71 49.33 15.15 43.31 19.80 54.63 26.43 F T est Encapsulation (A) 3.03 ns 1.86 ns 3.26 ns 2.03 ns 0.04 ns 0.18 ns 0.64 ns 2.69 ns 1.89 ns 0.05 ns 0.43 ns 0.40 ns Inoculum (B) 0.42 ns 0.1 1 ns 1.29 ns 0.58 ns 0.78 ns 0.09 ns 0.00 ns 0.64 ns 5.50 * 0.38 ns 0.05 ns 3.17 ns A x B 0.62 ns 0.85 ns 0.33 ns 1.22 ns 0.84 ns 0.21 ns 2.59 ns 0.57 ns 0.47 ns 0.63 ns 1.95 ns 1.01 ns
(1)Means followed by the same letter in the column do not dif
fer by the
Tukey test, being * significant
at 5% probability
, ** significant at 1% probability and ns not significant.
Conclusion
Plant growth promoting microorganisms, regardless of encapsulation used, promote higher development of caimito and lychee seedlings.
Acknowledgements
To the Coordination for the Improvement of Higher Education Personnel (CAPES), by granting a
scholarship to the first author; to Dr. José Carlos Barbosa
for his attention and help in the accomplishment of statistical analyses; to Luiz Carlos de Assis, for assistance in carrying out analyses; and Sidney Bedin, for their assistance in conduction of seedlings. To the São Paulo Research Foundation (FAPESP), for the
research funding number 2014/18313-8
References
ARAUJO NETO, S.E.; FREDNBERG, N.T.N.; MINOSSO, S.C.C., NOVELLI, D. da S.; ANDRADE NETO, R. de C. Substrate conditioners for prodution
of organic cupuasu seedling. Revista Brasileira de
Fruticultura, Jaboticabal, v.37, n.4, 1083-1088, 2015.
(1)
BARBOSA, J.C.; MALDONADO JÚNIOR, W.
Software AgroEstat: sistema de análises estatísticas de ensaios agronômicos. Jaboticabal: Universidade
Estadual Paulista, 2009. (2)
BARRY, A.L.; THORNSBERRY, C. Susceptibility tests: diffusion test procedures. In: BALOWS, A.; HAUSER, W.J.; HERMANN, K.L.; ISENBERG, H.D.;
SHAMODY, H.J. Manual of clinical microbiology.
5thed. Washington: American Society for Microbiology,
1991. p.1117-1125. (2)
BEUTLER, A.N.; BURG, G.M.; DEAK, E.A.;
SCHMIDT, M.R.; GALON, L. Effect of nitrogen-fixing bacteria on grain yield and development of flooded
irrigated rice. Revista Caatinga, Mossoró, v.29, n.1,
p.11-17, 2016. (1)
CARNEIRO, R.M.D.G.; GOMES, C.B. Encapsulação do
fungo Paecilomyces lilacinus em matrizes de
alginato-argila e avaliação da viabilidade dos conídios em duas
temperaturas. Nematologia Brasileira, Piracicaba,
8
C. H. B. Santos et al.
Rev. Bras. Frutic., Jaboticabal, 2018, v. 40, n. 3: (e-179)
CONSTANSKI, K.C.; ZORZETTI, J.; BOAS, G.T.V.; RICIETO, A.P.S.; FAZION, F.A.P.; BOAS, L.V.; MONNERAT, R.G.; NEVES, P.M.O.J. Seleção e
caracterização molecular de isolados de Bacillus
thuringiensis para o controle de Spodoptera spp. Pesquisa Agropecuária Brasileira, Brasília, DF, v.50, n.8,
p.730-773, 2015. (1)
C O N T R E R A S C O R N E J O , H . A . ; M A C Í A S -RODRÍGUEZ, L.; DEL-VAL, E.; LARSEN, J.Ecological
functions of Trichoderma spp.and their secondary
metabolites in the rhizosphere: interactions with plants.
FEMS Microbiology Ecology, Oxford, v.92, n.4, 2016. 1)
FAHAD, S.; HUSSAIN, S.; BANO, A.; SAUD, S.; HASSAN, S.; SHAN, D.; KHAN, F.; CHEN, Y.; WU, C.; TABASSUM, M.A.; CHUN, M.X.; AFZAL, M.; JAN, A.; JAN, M.T.; HUANG, J. Potential role of phytohormones and plant growth-promoting rhizobacteria in abiotic stresses: consequences for changing environment.
Environmental Science and Pollution Research,
Heidelberg, v.22, n.7, p.4907-4921, 2015. (1)
GÓES, G.S.; GROSS, E.; BRITO-ROCHA, E.; MIELKE,
M.S. Efeitos da inoculação com bactérias diazotróficas e
da adubação nitrogenada no crescimento e na qualidade de
mudas de Inga laurina (SW.) Willd.(Fabaceae). Revista
Árvore, Viçosa, MG, v.39, n.6, p.1031-1038, 2015. (1)
HARDOIM, P.R.; OVERBEEK, L.S.; BERG, G.; PIRTTILÄ, A.M.; COMPANT, S.; CAMPISANO, A.; DÖRIN, M.; SESSITSCH, A. The hidden world within plants: ecological and evolutionary considerations for defining functioning of microbial endophytes.
Microbiology and Molecular Biology Reviews,
Washington, v.79, n.3, p.293-320, 2015. (1)
HARTMANN, H.T.; KESTER, D.E.; DAVIES, F.T.;
GENEVE, R. Plant propagation:principles and practices.
8th ed. New Jersey: Prentice Hall, 2010. 928 p. (2)
HUNGRIA, M.; NOGUEIRA, M.A.; ARAUJO, R.S.
Soybean seed co-inoculation with Bradyrhizobium spp.
and Azospirillum brasilense: a new biotechnological tool
to improve yield and sustainability. American Journal
of Plant Sciences, Oxford, v.6, n.6, p.811-817, 2015. (1)
LAMBAIS, M.R.; LUCHETA, A.R.; CROWLEY, D.E. Bacterial community assemblages associated with the phyllosphere, dermosphere, andrhizosphere of tree species
of the Atlantic forest are host taxon dependent. Microbial
Ecology, New York, v.68, n.3, p.567–574, 2014. (1)
MAJEED, A; ABBASI, M.K.; HAMEED, S.; IMRAN, A.; RAHIM, N. Isolation and characterization of plant growth-promoting rhizobacteria from wheat rhizosphere
and their effect on plant growth promotion. Frontiers in
Microbiology, Lausanne, v.6, e-198, p.1-10, 2015. (1)
MOTA, M.S.; GOMES, C.B.; SOUZA JÚNIOR, I.T.; MOURA, A.B. Bacterial selection for biological control of plant disease: criterion determination and validation.
Brazilian Journal of Microbiology, São Paulo, v.48, n.1,
p.62-70, 2017. (1)
MÜLLER, T.M.; SANDINI, I.E.; RODRIGUES, J.D.; NOVAKOWISKI, J.H.; BASI, S.; KAMINSKI, T.H. Combination of inoculation methods of Azospirilum brasilense with broadcasting of nitrogen fertilizer
increases corn yield. Ciência Rural, Santa Maria, v.46,
n.2, p.210-215, 2016. (1)
NIETO-JACOBO, M.F.; STEYAERT, J.M.; SALAZAR-BADILLO, F.B.; NGUYEN, D.V.; ROSTÁS, M.; BRAITHWAITE, M.; SOUZA, J.T.; JIMENEZ-BREMONT, J.F.; OHKURA, M.; STEWART, A.; MENDOZA-MENDOZA, A. Environmental growth conditions of
Trichoderma spp.affects indol acetic acid derivatives, volatile organic compounds, and plant growth promotion.
Frontiers in Plant Science, Lausanne, v.8, e-102, p.1-18,
2017. (1)
PANKE-BUISSE, K.; POOLE, A.C.; GOODRICH, J.K.; LEY, R.E.; KAO-KNIFFIN, J. Selection on soil microbiomes reveals reproducible impacts on plant
function. The ISME Journal, London, v.9, n.4, p.980–
989, 2015. (1)
PEREG, L.; DE-BASHAN, L.E.; BASHAN, Y.Assessment
of affinity and specificity of Azospirillum for plants.Plant and soil, Dordrecht, v.399, n.1-2, p.389-414, 2016. (1)
PII, Y.; MIMMO, T.; TOMASI, N.; TERZANO, R.; CESCO, S.; CRECCHIO, C. Microbial interactions in
the rhizosphere: beneficial influences of plant
growth-promoting rhizobacteria on nutrient acquisition process.
A review. Biology and Fertility of Soils, New York, v.51,
n.4, p.403-415, 2015. (1)
SAHM, D.F., WASHINGTON II, J.A. Antibacterial susceptibility tests: dilution methods.In: BALOWS, A.; HAUSER, W.J.; HERMANN, K.L.; ISENBERG, H.D.;
SHAMODY, H.J. Manual of clinical microbiology.
5th ed.Washington: American Society for Microbiology,
SALLA, T.D.; SILVA, T.R.; ASTARITA, L.V.; SANTARÉM,
E.R. Streptomyces rhizobacteria modulate the secondary
metabolism of Eucalyptus plants. Plant Physiology and
Biochemistry, Paris, v.85, p.14-20, 2014. (1)
SCHOEBITZ, M.; LÓPEZ, M.; ROLDÁN, A. Bioencapsulation of microbial inoculants for better soil–
plant fertilization: a review. Agronomy for Sustainable
Development, Paris, v.33, n.4, p.751-765, 2013. (1)
SHEU, T.Y.; MARSHALL, R.T. Micro-encapsulation of
Lactobacilli in calcium alginate gels. Journal of Food
Science, Hoboken, v.54, n.3, p.557–561, 1993. (1)
SHI, Z.M. Technology for conidial preparation of
Beauveria bassiana. In: LI, Y.W.; LI, Z.Z.; LIANG,
Z.Q.; WU, J.W.; WU, Z.K.; XU, Q.F. (Ed.). Study and
application of entomogenous fungi in China. Beijing:
Academic Periodical Press, 1988. v.1, p.114–115. (2)
SILVA, R.L.de O.; LUZ, J.S.; SILVEIRA, E.B.da;
CAVALCANTE, U.M.T. Fungos endofíticos em Annona
spp: isolamento, caracterização enzimática e promoção do
crescimento em mudas de pinha (Annona squamosa L.).
Acta Botanica Brasilica, Belo Horizonte, v.20, n.3,
p.649-655, 2006. (1)
SOUZA, G.L.O.D.; SILVA, D.F.; NIETSCHE, S.; XAVIER, A.A.; PEREIRA, M.C.T. Endophytic bacteria used as bioinoculants in micropropagated banana seedlings.
Revista Brasileira de Fruticultura, Jaboticabal, v.39,
n.2, e-324, 2017. (1)
SOUZA, R. de; AMBROSINI, A.; PASSAGLIA, L.M.P. Plant growth-promoting bacteria as inoculants
in agricultural soils. Genetics and Molecular Biology,
Ribeirão Preto, v.38, n.4, p.401-419, 2015. (1)
SPASSIN, A.C.; VANDRESEN, P.B.; NEVES, D.; GARCIA, F.A.O.; UKAN, D.PERES, F.S.B. Promoção do
crescimento e enraizamento de miniestacas de Eucalyptus
dunnii por rizobactérias e bactérias do filoplano. Floresta,
Curitiba, v.46, n.3, p.387-395, 2016. (1)
SULTANA, K.; GODWARD, G.; REYNOLDS, N.; ARUMUGASWAMY, R.; PEIRIS, P.; KAILASAPATHY, K. Encapsulation of probiotic bacteria with alginate–starch and evaluation of survival in simulated gastrointestinal
conditions and in yoghurt. International Journal of Food
Microbiology, Amsterdam, v.62, n.1-2, p.47-55, 2000. (1)
VEMMER, M.; PATEL, A.V. Review of encapsulation methods suitable for microbial biological control agents.
Biological Control, San Diego, v.67, n.3, p.380-389,
2013. (1)
XU, S.J.; PARK, D.H.; KIM, J.-Y.; KIM, B.-S. Biological control of gray mold and growth promotion of tomato
using Bacillus spp. isolated from soil. Tropical Plant
Pathology, Brasília, DF, v.41, n.3, p.169-176, 2016. (1)
ZHOU, C.; MA, Z.; ZHU, L.; XIAO, X.; XIE, Y.; ZHU,
J.; WANG, J. Rhizobacterial strain Bacillus megaterium
BOFC15 induces cellular polyamine changes that improve
plant growth and drought resistance. International
Journal of Molecular Sciences, Basel, v.17, n.6, e-976,