Universidade do Minho
Escola de Engenharia
Luís Filipe da Silva Fernandes
14
Luís F
ilipe da Silva F
ernandes
Envelhecimento Cognitivo – Um estudo com
Eletroencefalografia e Ressonância Magnética
Funcional
Envelhecimento Cognitivo – Um es tudo com Ele troencefalografia e R essonância Magnética FuncionalDissertação de Mestrado
Mestrado em Engenharia Biomédica
Ramo de Eletrónica Médica
Trabalho efetuado sob a orientação do
Professor Doutor Carlos Manuel Gregório Santos Lima
e do
Professor Doutor Nuno Sérgio Mendes Dias
Universidade do Minho
Escola de Engenharia
Luís Filipe da Silva Fernandes
Envelhecimento Cognitivo – Um estudo com
Eletroencefalografia e Ressonância Magnética
Funcional
DECLARAÇÃO Nome: Luís Filipe da Silva Fernandes
Endereço eletrónico: luis.filipe.sf@gmail.com
Cartão do Cidadão: 13932468
Título da dissertação: Envelhecimento Cognitivo – Um estudo com Eletroencefalografia e Ressonância Magnética Funcional
Orientadores:
Professor Doutor Carlos Manuel Gregório Santos Lima Professor Doutor Nuno Sérgio Mendes Dias
Ano de conclusão: 2014
Mestrado em Engenharia Biomédica Ramo de Eletrónica Médica
É AUTORIZADA A REPRODUÇÃO INTEGRAL DESTA DISSERTAÇÃO APENAS PARA EFEITOS DE INVESTIGAÇÃO, MEDIANTE DECLARAÇÃO ESCRITA DO INTERESSADO, QUE A TAL SE COMPROMETE.
Universidade do Minho, _____/_____/_________ Assinatura:
A
GRADECIMENTOS
Um agradecimento muito especial ao Professor Doutor Nuno Dias pela orientação, apoio, confiança e sobretudo dedicação que sempre demonstrou ao longo deste ano.
Um obrigado especial ao Professor Pedro Almeida e à Joana Festa pela ajuda, disponibilidade e transmissão de novos conhecimentos, que foram tão importantes para a realização desta dissertação.
Ao Professor Doutor Carlos Lima pelo interesse, ajuda e colaboração.
Ao Nuno Afonso, João Ferreira, João Meireles, Joana Reis, Joana Festa, Cristiana Merendeiro, Ana Maria, Daniela Ferreira e Mariana Pereira, meus colegas de trabalho que rapidamente se tornaram amigos. Um profundo agradecimento a este grupo e a estas pessoas fantásticas pelo apoio, ajuda e sobretudo pela amizade e pelos bons momentos que passámos.
Ao Paulo Marques, Pedro Moreira e Ricardo Magalhães, os “meninos” da ressonância magnética, pela disponibilidade e empenho durante as aquisições funcionais, bem como pela ajuda durante a escrita da dissertação.
Às várias instituições que tornaram esta dissertação possível, ICVS, CCA e Hospital de Braga, bem como a todos os participantes pelo interesse e disponibilidade.
Aos meus amigos e colegas de curso pela amizade e incentivo ao longo deste ano.
Finalmente, um agradecimento muito especial aos meus pais, irmão e padrinhos pela preocupação e confiança demonstradas. Sem o seu apoio, esta dissertação não seria possível.
R
ESUMO
O cérebro, tal como qualquer outro tecido do corpo humano, sofre um processo natural de envelhecimento através de alterações neuroquímicas, estruturais e funcionais, o que leva a uma perda progressiva de plasticidade cerebral. Este declínio é refletido principalmente em perda de memória episódica, diminuição da capacidade da memória de trabalho e de tomada de decisão, perda de flexibilidade cognitiva e atenção seletiva, entre outras funções cognitivas. O declínio cognitivo induzido pelo envelhecimento afeta um importante segmento da população e é o principal fator de risco da doença de Alzheimer, entre outras doenças neurodegenerativas. Desta forma, a deteção e avaliação do declínio cognitivo induzido pelo envelhecimento é de extrema importância para a sociedade atual, uma sociedade cada vez mais envelhecida.
O objetivo inicial focou-se na implementação e estabilização das técnicas combinadas de EEG e fMRI no Hospital de Braga. Na segunda fase do trabalho, utilizou-se o WCST como um instrumento neuropsicológico sensível a várias funções cognitivas como a tomada de decisão, memória de trabalho e flexibilidade cognitiva, com o objetivo de avaliar a componente P300 do potencial evocado como um marcador de défice cognitivo associado ao envelhecimento. Nesta fase, as características extraídas da componente, bem como a análise dos ensaios iniciais e finais, permitiu avaliar a relação entre P300 e idade, testar a mudança atencional aquando de uma nova categoria, bem como a teoria da atualização de um modelo de estímulo (nesta caso, a categoria) em memória durante o desempenho do WCST. Numa terceira fase, o mesmo teste foi realizado enquanto se adquiria simultaneamente sinais com as técnicas de EEG e fMRI, avaliando assim o efeito do envelhecimento cognitivo na perspetiva destas duas técnicas simultâneas.
Inicialmente, a técnica de aquisição combinada de EEG e fMRI foi implementada com sucesso no Hospital de Braga. Os estudos realizados permitiram concluir que a latência e amplitude de pico da componente P300, bem como a sua largura, parecem discriminar sujeitos por idade e desempenho como resultado da aplicação do WCST. As diferenças observadas entre os ensaios finais e iniciais do WCST sugerem a atualização gradual de uma representação da memória de trabalho para a categoria do estímulo em vigor. Os resultados provenientes da análise das imagens de ressonância magnética funcional, em paralelo com aqueles obtidos através dos dados de EEG, parecem sugerir que os sujeitos jovens ativam mais as regiões parietais em comparação com os sujeitos idosos, durante o desempenho
do teste de cartas de Wisconsin, e que esta diferença poderá dever-se a défices no recrutamento de regiões cerebrais com relevância nos mecanismos de memória de trabalho e atenção seletiva.
A
BSTRACT
The brain, like every other human tissue, suffers a natural process of aging where it suffers several neurochemical, structural and functional changes, which leads to brain plasticity loss. These deficits are primarily reflected on episodic memory, cognitive flexibility and selective attention loss, decrease of working memory and decision making capacity and other cognitive impairments. The cognitive decline prompted by aging affects a major segment of the population and it is the primary risk factor for Alzheimer’s disease and other prevalent neurodegenerative disorders. Thus, the cognitive decline detection and evaluation are very important for our aged society.
The initial goal focused on the implementation and stabilization of the EEG and fMRI combined techniques at Hospital de Braga. In the second phase of this work, it was used the WCST as a neuropsychological instrument sensitive to several cognitive functions such as decision making, working memory and cognitive flexibility, with the purpose of evaluating the P300 component of the evoked potential as a marker for cognitive deficits associated with aging. At this stage, the features extracted from the component, as well as early-late trial analysis, enabled the assessment of the relationship between P300 and age, and tested the attentional set shifting when category shifts and working memory updating hypothesis for the stimulus category during WCST performance. In a third phase, the same test was conducted while EEG and fMRI signals were acquired simultaneously, assessing the effect of cognitive aging from the perspective of these two combined techniques.
At first, the combined techniques EEG and fMRI was implemented successfully at Hospital de Braga. From the conducted studies we concluded that P300 component peak latency and amplitude, as well as its peak width, seem to discriminate subjects by age and performance as a result of WCST testing. The observed differences between early and late trials of the WCST suggest the gradual updating of a working memory representation for the stimulus category. The results from the functional magnetic resonance imaging analysis, in parallel with those obtained from the EEG data analysis, seem to suggest that younger subjects activate more parietal regions than elderly subjects while WCST testing, and that this difference may be due to deficits on the recruitment of important brain regions associated with working memory and selective attention mechanisms.
Í
NDICE Agradecimentos ... iii Resumo... v Abstract... vii Lista de Figuras ... xi Lista de Tabelas ... xvLista de Abreviaturas, Siglas e Acrónimos ... xvii
1. Introdução ... 1 1.1 Motivação ... 1 1.2 Objetivos ... 2 1.3 Contribuições ... 2 1.4 Estrutura da Dissertação ... 3 2. Conceitos teóricos ... 5 2.1 Neurónio ... 5 2.2 Cérebro ... 8 2.3 Eletroencefalografia ... 12
2.4 Ressonância Magnética Funcional ... 21
2.5 Teste de cartas de Wisconsin... 30
3. Estado da arte ... 33
3.1 Envelhecimento cognitivo ... 33
3.2 Do envelhecimento cognitivo saudável à demência ... 34
3.3 Eletrofisiologia do envelhecimento cognitivo ... 35
3.4 Imagiologia do envelhecimento cognitivo... 37
3.5 Técnica EEG-fMRI e o envelhecimento cognitivo ... 39
3.6 Estudos com o teste de cartas de Wisconsin ... 40
4. Análise da relação entre P300 e idade durante o desempenho do wcst ... 45
4.1 Metodologia ... 45
4.1.1 Sujeitos ... 45
4.1.2 Aquisição do Eletroencefalograma ... 46
4.1.4 Procedimento Experimental ... 49
4.1.5 Processamento de Sinal do Eletroencefalograma ... 49
4.1.6 Tratamento Estatístico ... 55
4.2 Resultados e Discussão ... 56
5. Efeito do envelhecimento cognitivo na perspetiva da aquisição simultânea de EEG e fMRI ... 67
5.1 Metodologia ... 67
5.1.1 Sujeitos ... 67
5.1.2 Aquisição do Eletroencefalograma ... 68
5.1.3 Aquisições de Imagem por Ressonância Magnética Funcional ... 70
5.1.4 Teste de cartas de Wisconsin ... 71
5.1.5 Procedimento Experimental ... 73
5.1.6 Interação entre o Sinal do Eletroencefalograma e a Ressonância Magnética Funcional 74 5.1.7 Processamento de Sinal do Eletroencefalograma ... 77
5.1.8 Processamento das Imagens da Ressonância Magnética Funcional ... 82
5.1.9 Tratamento Estatístico ... 90 5.2 Resultados e Discussão ... 92 6. Conclusões ... 105 6.1 Conclusões ... 105 6.2 Perspetivas Futuras ... 106 Bibliografia ... 107
Anexo I – Conversor fibra ótica em TTL ... 123
Anexo II – Esquema do Sistema EEG+fMRI ... 125
L
ISTA DE
F
IGURAS
Figura 1. Morfologia de um neurónio e fluxo do impulso elétrico através deste.. ... 6
Figura 2. Potencial de ação (potencial na membrana do axónio) de uma lula gigante.. ... 7
Figura 3. a) Ilustração de uma vista sagital do cérebro humano. b) Vista sagital do cérebro humano através de uma ressonância magnética. ... 8
Figura 4. Divisões do sistema nervoso central. ... 9
Figura 5. Decomposição de um sinal de EEG nos quatro principais ritmos cerebrais (delta, teta, alfa e beta). ... 16
Figura 6. Diferenças numa componente do ERP entre sujeitos alcoólicos e não alcoólicos. ... 18
Figura 7. ERP médio de 40 sujeitos com fadiga mental e antes de possuírem esta condição. ... 18
Figura 8. Subcomponentes P3a e P3b da componente P300 do ERP. ... 19
Figura 9. Modelo da atualização contextual. ... 20
Figura 10. Precessão de uma partícula atómica giratória na presença do campo magnético primário 𝐵0. ... 23
Figura 11. Eixos ortogonais que definem os vários planos de corte orientados com o campo magnético primário 𝐵0. ... 23
Figura 12. a) Variação do relaxamento longitudinal ao longo do tempo desde a interrupção do pulso radiofrequência; b) Variação do relaxamento transversal ao longo do tempo desde a interrupção do pulso radiofrequência. ... 25
Figura 13. Decaimento de indução livre (FID). ... 26
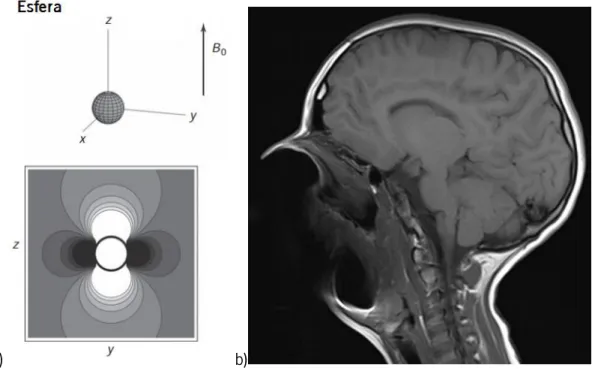
Figura 14. a) Distorção causada no campo magnético primário por uma esfera metálica. b) Distorção causada por um aparelho dentário completo numa imagem de ressonância magnética. ... 27
Figura 15. Comparação entre o tempo de decaimento T2 e T2*. ... 28
Figura 16. Imagem de uma hemorragia interna cerebral utilizando o contraste T2*. ... 29
Figura 17. Cartas padrão do teste de cartas de Wisconsin. ... 30
Figura 18. Disposição dos elétrodos no registo de EEG utilizado. ... 47
Figura 19. Procedimento utilizado no teste de cartas de Wisconsin (estudo Análise da relação entre P300 e idade durante o desempenho do WCST). ... 48
Figura 20. Mapas topográficos padrão das componentes independentes dos a) artefactos oculares piscar de olhos (mapa topográfico à esquerda) e b) sacadas oculares (mapa topográfico à direita). ... 52
Figura 21. Artefacto ocular sacada (curva azul escuro) a ser atenuado com a remoção da componente independente correspondente (curva vermelha). ... 52 Figura 22. Artefacto ocular piscar de olhos (curva azul escuro) a ser atenuado com a remoção da componente independente correspondente (curva vermelha). ... 53 Figura 23. Topografia centro-parietal padrão de uma componente P300. ... 54 Figura 24. Percentagem de sessões sem a componente P300 detetável para os três grupos etários. . 56 Figura 25. a) Valor médio e desvio padrão da latência de pico da componente P300 para cada grupo etário; b) Valor médio e desvio padrão da amplitude de pico da componente P300 para cada grupo etário. ... 57 Figura 26. Sincronização da componente P300 ao longo das várias sessões para cada grupo etário. O tempo é representado no eixo X. O eixo Y representa os sujeitos à medida que a idade aumenta. ... 58 Figura 27. a) Valor médio e desvio padrão da latência de pico da componente P300 para cada grupo de desempenho no WCST; b) Valor médio e desvio padrão da amplitude de pico da componente P300 para cada grupo de desempenho no WCST. ... 59 Figura 28. Sincronização da componente P300 ao longo das várias sessões para cada grupo de desempenho no WCST. O tempo é representado no eixo X. O eixo Y representa os sujeitos à medida que o desempenho diminui. ... 60 Figura 29. a) Valor médio e desvio padrão da latência de pico da componente P300 nos ensaios iniciais e finais considerando todos os sujeitos com componente P300 observável; b) Valor médio e desvio padrão da amplitude de pico da componente P300 nos ensaios iniciais e finais considerando todos os sujeitos com componente P300 observável. ... 62 Figura 30. a) Valor médio e desvio padrão da latência de pico da componente P300, nos ensaios iniciais, para cada grupo etário; b) Valor médio e desvio padrão da amplitude de pico da componente P300, nos ensaios iniciais, para cada grupo etário. ... 63 Figura 31. a) Valor médio e desvio padrão da latência de pico da componente P300, nos ensaios finais, para cada grupo etário; b) Valor médio e desvio padrão da amplitude de pico da componente P300, nos ensaios finais, para cada grupo etário. ... 63 Figura 32. Versão utilizada do sistema de localização internacional 10-20. ... 68 Figura 33. Touca de 64 elétrodos utilizada durante as aquisições no estudo “Efeito do envelhecimento cognitivo na perspetiva da aquisição simultânea de EEG e fMRI”. ... 69 Figura 34. a) Máquina de ressonância magnética Siemens Magnetom Avanto 1,5 T. b) Bobina RF utilizada para a cabeça. ... 70
Figura 35. Disposição do sistema ressonância magnética, bobine RF, amplificadores, touca e sujeito
durante as aquisições simultâneas de imagem e EEG. ... 71
Figura 36. Procedimento utilizado no teste de cartas de Wisconsin (estudo Efeito do envelhecimento cognitivo na perspetiva da aquisição simultânea de EEG e fMRI)... 72
Figura 37. Efeito do artefacto de gradiente nos sinais de EEG de um participante do estudo. ... 74
Figura 38. Correção do artefacto de gradiente nos sinais de EEG de um participante do estudo. ... 75
Figura 39. Efeito do artefacto cardiobalístico nos sinais de EEG de um participante do estudo. ... 76
Figura 40. Correção do artefacto cardiobalístico nos sinais de EEG de um participante do estudo. ... 76
Figura 41. Local de colocação do elétrodo de ECG. ... 78
Figura 42. Topografia centro-parietal padrão de uma componente P300. ... 81
Figura 43. a) Fatia aleatória de um volume; b) Intensidades da resposta BOLD antes do movimento lateral; c) Intensidades da resposta BOLD depois do movimento lateral. ... 83
Figura 44. Efeitos do movimento da cabeça ao longo do tempo em 4 imagens funcionais. ... 84
Figura 45. Seleção da primeira imagem como imagem de referência no processo de realinhamento.. 85
Figura 46. Ilustração do cálculo dos parâmetros necessários à transformação de corpo rígido. ... 85
Figura 47. Realinhamento de todas as imagens funcionais em relação à imagem de referência). ... 86
Figura 48. Transformações lineares necessárias ao passo da normalização... 87
Figura 49. Imagens funcionais dos vários cortes do cérebro antes da normalização; b) Imagens funcionais dos vários cortes do cérebro após normalização. ... 88
Figura 50. Representação de um filtro gaussiano bidimensional e efeito deste no voxel central de uma região cerebral. ... 88
Figura 51. Ilustração da medida FWHM em relação ao filtro gaussiano. ... 89
Figura 52. Imagens funcionais dos vários cortes do cérebro antes da filtragem espacial; b) Imagens funcionais dos vários cortes do cérebro após filtragem espacial. ... 90
Figura 53. a) Valor médio e desvio padrão da latência de pico da componente P300, medidos no elétrodo Pz, para cada grupo etário; b) Valor médio e desvio padrão da amplitude de pico da componente P300, medidos no elétrodo Pz, para cada grupo etário. ... 94
Figura 54. Resposta média da componente P300 para cada um dos grupos etários, no elétrodo Pz. O tempo é representado no eixo X. O eixo Y representa a amplitude do sinal. ... 94 Figura 55. a) Valor médio e desvio padrão da latência de pico da componente P300, medidos no elétrodo Fz, para cada grupo etário; b) Valor médio e desvio padrão da amplitude de pico da componente P300,
Figura 56. Resposta média da componente P300 para cada um dos grupos etários, no elétrodo Fz. O tempo é representado no eixo X. O eixo Y representa a amplitude do sinal. ... 96 Figura 57. Ativações cerebrais significativas durante a baseline do teste, tendo em conta todos os sujeitos. Utilizou-se a probabilidade individual do voxel de 0,01 e o limite inferior para o tamanho dos clusters obtidos de 157 voxels. ... 98 Figura 58. Ativações cerebrais significativas durante o evento “carta”do teste, tendo em conta todos os sujeitos. Utilizou-se a probabilidade individual do voxel de 0,01 e o limite inferior para o tamanho dos clusters obtidos de 107 voxels. ... 99 Figura 59. Ativações cerebrais significativas da relação das ativações do bloco de teste exceto as ativações do bloco de baseline (Teste - Baseline), tendo em conta todos os sujeitos. Utilizou-se a probabilidade individual do voxel de 0,01 e o limite inferior para o tamanho dos clusters obtidos de 119 voxels. ... 100 Figura 60. Diferenças significativas nas ativações neuronais entre os sujeitos jovens e idosos (Ativações dos jovens > Ativações dos idosos), considerando a relação das ativações do bloco de teste exceto as ativações do bloco de baseline (Teste - Baseline). Utilizou-se a probabilidade individual do voxel de 0,05 e o limite inferior para o tamanho dos clusters obtidos de 245 voxels. ... 102
L
ISTA DE
T
ABELAS
L
ISTA DE
A
BREVIATURAS
,
S
IGLAS E
A
CRÓNIMOS
EEG – Electroencefalograma ECG – Electrocardiograma
fMRI – Imagiologia por Ressonância Magnética Funcional TTL – Lógica Transístor-Transístor
BOLD – Sinal Dependente dos Níveis de Oxigénio no Sangue ICA – Análise de Componentes Independentes
RF – Radiofrequência FOC – Cabo de Fibra Ótica TR – Tempo de Repetição TE – Tempo de Eco
RM – Ressonância Magnética
PET – Tomografia por Emissão de Positrões PFC – Córtex Pré-Frontal
GLM – Modelo Linear
SPM – Mapas Estatísticos Paramétricos FWER – Taxa de Erro Familywise RFT – Teoria de Campo Aleatório WCST – Teste de Cartas de Wisconsin
1. I
NTRODUÇÃO
Neste capítulo é apresentada a motivação para a elaboração do trabalho desenvolvido ao longo desta dissertação, bem como os objetivos definidos. São ainda enumeradas as contribuições alcançadas com este trabalho e descrita a organização da dissertação.
1.1 Motivação
Nos países mais desenvolvidos, o envelhecimento é um dos flagelos sociais que recentemente tem afetado estas sociedades. Os vários avanços tecnológicos alcançados neste último século na área da medicina, no tratamento de várias doenças que afetam principalmente pessoas jovens ou de meia-idade, bem como a melhoramento das condições sanitárias tanto das casas como dos hospitais, fizeram com que a esperança média de vida atingisse os valores mais elevados de sempre. De acordo com estatísticas oficiais, o número de pessoas em Portugal com idade superior a 65 anos por cada 100 pessoas com idades entre os 0 e os 14 anos aumentou 102,4% de1960 a 2012 (PORDATA, 2012b). Este rácio denomina-se índice de envelhecimento, e os restantes países da União Europeia seguem esta tendência. Em paralelo com o envelhecimento etário, verifica-se um envelhecimento cognitivo. O cérebro, tal como qualquer tecido humano, sofre um processo natural de envelhecimento. Vários estudos, recorrendo principalmente à técnica de ressonância magnética funcional, afirmam que apesar do cérebro envelhecer, este tem a capacidade de manter um valor considerável de plasticidade das várias funções cognitivas. Porém, ainda não é totalmente compreendido como é que as diferenças etárias afetam a utilização destas funções cognitivas, ou seja, afetam o desempenho cognitivo dos sujeitos mais idosos em comparação com os mais jovens (C. Grady, 2012). Vários estudos relacionam este declínio cognitivo, manifestado na sua maioria através de perda de memória episódica, diminuição na capacidade de memória de trabalho entre outras alterações cognitivas, com transformações neuroquímicas, estruturais e funcionais do cérebro (Nyberg, Lövdén, Riklund, Lindenberger, & Bäckman, 2012; Rossini, Rossi, Babiloni, & Polich, 2007; Tomasi & Volkow, 2012). O declínio cognitivo resultante do envelhecimento afeta grande parte da população, sendo que o principal fator de risco é a doença de Alzheimer (Yankner, Lu, & Loerch, 2008).
Esta dissertação pretende contribuir para a compreensão da relação entre o declínio cognitivo e a idade, através do estudo dos efeitos da idade em correlatos eletrofisiológicos e em imagem de
1.2 Objetivos
Tendo em conta a motivação desta dissertação, vários objetivos precisam de ser estabelecidos e alcançados. Um dos principais objetivos é a implementação e estabilização da técnica de aquisição de ressonância magnética funcional em simultâneo com a técnica de eletroencefalografia (EEG) no Hospital de Braga.
Através da realização de uma tarefa que envolva alguns dos principais mecanismos cognitivos afetados pela idade, nomeadamente memória de trabalho e flexibilidade cognitiva, pretende-se estudar os efeitos da idade no desempenho cognitivo através da análise de características da eletroencefalografia e da ressonância magnética funcional.
Por fim, pretende-se compreender se as características da eletroencefalografia e da ressonância magnética funcional selecionadas podem ser interpretadas como correlatos de envelhecimento saudável.
1.3 Contribuições
A partir desta dissertação foram implementadas com sucesso as técnicas combinadas de eletroencefalografia e ressonância magnética funcional no Hospital de Braga. Considero que esta seja uma contribuição com relevância pois “abre portas” a novos e diferentes estudos na Universidade do Minho e na sua parceria com o Hospital de Braga, materializada através do Centro Clínico-Académico (C2A), onde foram realizadas todas as recolhas de dados.
Para além da implementação e estabilização das técnicas combinadas de eletroencefalografia e ressonância magnética funcional, foi construído um dispositivo específico para a ressonância magnética do Hospital de Braga, Siemens Magnetom Avanto 1,5T (Siemens AG, Munique, Baviera, Alemanha), poder fazer comunicação dos eventos das aquisições com o computador de recolha dos dados eletroencefalográficos. Este dispositivo consiste num conversor de fibra ótica em TTL. Mais informações sobre este dispositivo encontram-se disponibilizadas no Anexo I – Conversor fibra ótica em TTL.
A análise dos efeitos da idade sobre a componente P300 durante o desempenho do Teste de Cartas de Wisconsin (WCST) motivou a submissão de um artigo de proceedings para a Neural Engineering Conference 2015, intitulado “Aging and Attentional Set Shifting on WCST: An event-related EEG study”. Artigo integral no Anexo III – Artigo de Proceedings para Conferência.
1.4 Estrutura da Dissertação
Esta dissertação está dividida em 6 capítulos. No presente capítulo estabelece-se quais as motivações inerentes ao desenvolvimento desta dissertação, bem como os objetivos principais que se pretendem atingir com a mesma. São ainda enumeradas as contribuições alcançadas com este trabalho. No segundo capítulo abordam-se os fundamentos teóricos que servem de suporte ao trabalho realizado. Começa-se por descrever os conceitos de neurónio e transmissão de informação e descrição dos vários lobos constituintes do cérebro. De seguida descrevem-se as técnicas utilizadas durante a realização deste trabalho, ou seja, a técnica de eletroencefalografia e a técnica de ressonância magnética funcional, bem como as suas vantagens e desvantagens. No capítulo 2 é ainda abordado o tema do envelhecimento cognitivo e os seus efeitos no eletroencefalograma e nas imagens resultantes da ressonância magnética funcional. Por fim, descreve-se a tarefa cognitiva realizada pelos sujeitos, o teste de cartas do Wisconsin, bem como a motivação para a escolha da mesma. No capítulo 3 é estabelecido o estado-da-arte.
No capítulo 4 é feita a análise dos efeitos da idade na componente P300 do potencial evocado durante o desempenho do teste de cartas de Wisconsin, com aquisição simultânea de EEG. São ainda apresentados os resultados da análise e feita a discussão dos mesmos.
No capítulo 5, os efeitos da idade na componente P300 evocada durante o WCST são analisados com recurso às técnicas de eletroencefalografia e ressonância magnética funcional em simultâneo. São também apresentados os resultados da análise e feita a discussão dos mesmos.
No último capítulo são apresentadas as principais conclusões de ambas as análises, bem como as perspetivas futuras nesta área de estudo.
2. C
ONCEITOS TEÓRICOS
2.1 Neurónio
O neurónio é um constituinte fundamental do sistema nervoso central (CNS), pois é o responsável pelo transporte de informação ao longo do corpo. Este transporte de informação é efetuado através da transmissão de impulsos elétricos ao longo da célula nervosa como resposta a um estímulo. O sistema nervoso central é também constituído pelas células de glia, que se dispõem entre os neurónios. Uma das principais funções destas células, nos adultos, é a formação do revestimento de mielina à volta dos axónios dos neurónios (K. R. Jessen, 2004; Pfaff, 2013).
Em termos anatómicos, os neurónios são constituídos pelo corpo celular, dendrites e axónios (Figura 1). O corpo celular possui apenas um núcleo e é responsável por grande parte de todo o metabolismo celular. O axónio tem forma cilíndrica e é bastante longo quando comparado com os restantes constituintes da célula nervosa. Nos humanos, os axónios podem atingir mais de um metro de comprimento. Este é responsável pela condução dos impulsos elétricos desde o corpo celular até ao terminal do axónio, ou seja, até às dendrites do próximo neurónio. Por fim, as dendrites estão diretamente ligadas ao axónio ou a outras dendrites de outras células nervosas e permitem fazer a receção dos estímulos que chegam ao neurónio. A conexão entre um neurónio e as outras células nervosas “vizinhas” faz com que se forme uma vasta rede de transmissão de informação (Sanei, 2013). Existem três tipos principais de neurónios de acordo com a sua função: neurónios motores ou eferentes, que transmitem informação motora desde o sistema nervoso central até aos órgãos motores; neurónios sensoriais ou aferentes, que transmitem informação sensorial; interneurónios ou associativos que transmite informação entre diferentes tipos de neurónios (Pfaff, 2013).
Figura 1. Morfologia de um neurónio e fluxo do impulso elétrico através deste. Adaptado de (Sanei, 2013).
Em termos fisiológicos, a principal função do neurónio é processar o estímulo recebido e transportá-lo até à próxima célula nervosa. Este transporte até ao próximo neurónio é realizado nas fendas sinápticas, ou sinapses, que separam o axónio de outras dendrites, ou dendrites de outras dendrites (Sanei, 2013).
A transmissão dos impulsos elétricos ao longo de um neurónio, potencial de ação, é um fenómeno temporário e repentino, e assenta no facto de a membrana do axónio possuir internamente um potencial de -60 a -70 mV (potencial de repouso). Com a chegada do potencial de ação ao início do axónio, o potencial de repouso é perturbado, fazendo com que comece a despolarização da membrana. A despolarização provoca o desvio do potencial interno da membrana para valores ligeiramente positivos. Após a despolarização (passagem do potencial de ação) verifica-se a repolarização, que é o retorno do potencial interno da membrana para valores de repouso. O potencial, inclusive, desce para valores mais baixos que o potencial de repouso (hiperpolarização) de modo a evitar que o potencial de ação se desloque no sentido inverso ao sentido normal de propagação, ou seja, do corpo celular para as dendrites. Esta sequência de despolarizações e repolarizações estende-se ao longo do axónio formando uma onda de transmissão do potencial de ação, percorrendo assim toda a célula nervosa (Sanei, 2013).
Figura 2. Potencial de ação (potencial na membrana do axónio) de uma lula gigante. Adaptado de (Sanei, 2013).
Um potencial de ação só ocorre caso, devido a um estímulo, o potencial interno da membrana
ultrapasse um determinado threshold, que normalmente situa-se à volta de -55 mV. Quando este
threshold é superado verifica-se a abertura dos canais de sódio, entrando iões positivos de sódio, ou seja, fazendo com que o interior membranar fique mais positivo (despolarização). Com este aumento de potencial, os canais de potássio abrem e os de sódio fecham pois foi atingida a concentração de equilíbrio para este ião. A abertura dos canais de potássio existentes na membrana do axónio leva à saída de iões positivos de potássio, ficando o interior membranar cada mais negativo (repolarização), ultrapassando inclusive o potencial de repouso. Estes canais fecham quando é atingido a concentração de equilíbrio destes iões. Durante a hiperpolarização os canais de sódio e potássio estão fechados e apenas estão a funcionar as bombas de sódio/potássio, fazendo com que o potencial atinja o potencial de repouso da membrana (Sanei, 2013).
Depois de o potencial de ação percorrer a totalidade da célula nervosa, este terá de ser transferido para o próximo neurónio. Esta transferência ocorre na fenda sináptica, como foi referido anteriormente, através de neurotransmissores químicos (serotonina, acetilcolina, dopamina, entre outros). Os neurotransmissores são sintetizados no terminal do axónio e são recebidos em recetores específicos situados nas dendrites da próxima célula nervosa. Desta forma, é possível verificar que, neste caso, os neurónios estão a responder a estímulos químicos e não elétricos (Kandel, Schwartz, & Jessell, 2000).
2.2 Cérebro
O sistema nervoso é constituído em duas partes: o sistema nervoso periférico e o sistema nervoso central. O primeiro é a parte do sistema nervoso constituído pelas células nervosas e de glia que estão situadas fora do cérebro e da medula espinal. A função do sistema nervoso periférico assenta na distribuição bidirecional dos impulsos elétricos provenientes do sistema nervoso central até aos membros superiores, inferiores e aos vários órgãos, ou seja, na troca de “informação” entre o sistema nervoso central e as extremidades e órgãos (Kandel et al., 2000).
Figura 3. a) Ilustração de uma vista sagital do cérebro humano. Adaptado de (Kandel et al., 2000). b) Vista sagital do cérebro humano através de uma ressonância magnética Adaptado de (Kandel et al., 2000).
O sistema nervoso central é constituído pelo cérebro e pela medula espinal. Pelo facto de este sistema ser o mais importante do nosso organismo, estes elementos encontram-se protegidos por osso, sendo que o cérebro tem o crânio como proteção e a medula espinal as vértebras. A elevada relevância atribuída ao sistema nervoso central deve-se ao facto de este ser o centro de todo o processamento de informação necessário ao normal funcionamento do organismo. A medula espinal recebe informação sensorial dos vários músculos e pele existentes no tronco e extremidades e é responsável pelos movimentos voluntários e involuntários devido à presença dos neurónios motores na sua constituição (Kandel et al., 2000).
O cérebro, ilustrado na Figura 3.a), é o principal e mais complexo constituinte do sistema nervoso central e pode ser dividido em 6 partes: bulbo raquidiano, ponte de Varólio, mesencéfalo, cerebelo, diencéfalo e telencéfalo. Cada uma destas 6 divisões podem ainda ser subdividas em áreas anatomicamente e funcionalmente distintas. O bulbo raquidiano, ponte de Varólio e o mesencéfalo são os constituintes do denominado tronco cerebral, que se encontra entre o cérebro e a medula espinal. As principais funções deste constituinte são as funções de interligação entre o cérebro e a medula espinal
e a função de regulação dos mecanismos homeostáticos mais básicos, como o ritmo cardíaco, a dor ou a respiração. O cerebelo encontra-se entre a ponte de Varólio e a parte mais occipital do cérebro e tem funções bastante específicas. Este é responsável pela postura bípede dos humanos, pela coordenação dos movimentos da cabeça e olhos e ainda possui uma função importante na regulação motora, bem como na sua aprendizagem (Kandel et al., 2000). O diencéfalo é constituído por dois componentes principais, o tálamo e o hipotálamo. As principais funções deste constituinte do cérebro são a interligação entre a informação sensorial proveniente das extremidades sensoriais e as regiões sensoriais do telencéfalo, e ainda a regulação da homeostasia, de vários processos metabólicos e hormonais (Kandel et al., 2000).
O telencéfalo, ou hemisférios cerebrais, é responsável pelas funções conscientes do sistema nervoso central, isto é, as várias funções cognitivas e ainda algumas motoras. A memória e formação de emoções são exemplos da complexidade das funções inerentes ao telencéfalo. Este é o maior constituinte do cérebro e divide-se em dois hemisférios, o hemisfério direito e o esquerdo. A ligação entre os dois hemisférios é feita através do corpo caloso, cuja principal função é mesmo a de intercomunicação entre regiões similares de ambos os hemisférios. A parte mais exterior do telencéfalo é denominada córtex cerebral e pode ser dividida em 4 lobos: o frontal, parietal, temporal e occipital (Kandel et al., 2000) (Figura 4).
Figura 4. Divisões do sistema nervoso central. Adaptado de (Kandel et al., 2000).
O lobo frontal situa-se, tal como o nome indica, na zona frontal do cérebro e é delimitado pelo sulco central e pela fissura lateral. A zona imediatamente junta ao sulco central denomina-se córtex motor primário e é responsável pela regulação dos movimentos voluntários. Duas áreas também bastante importantes são o córtex pré-motor e a área pré-frontal. A primeira tem como função a coordenação da
fala (um dos movimentos mais complexos do nosso organismo), entre outros movimentos. A área pré-frontal está relacionada com atividades cerebrais de grande complexidade como o pensamento e raciocínio (John W. Clark, 2010). Este lobo também está especializado no processamento da memória de curto-prazo e no planeamento de ações futuras (Kandel et al., 2000).
O lobo parietal encontra-se na parte posterior do cérebro e é delimitado pelo sulco central, pelo lobo temporal e pelo lobo occipital. A zona imediatamente após o sulco central, no giro central posterior, denomina-se córtex somatossensorial primário e é responsável pela perceção de sensações (como por exemplo pressão, dor ou toque) dos diferentes recetores sensoriais espalhados pela pele. Outras funções importantes deste lobo são a sensação espacial, navegação (função propriocetiva) e ainda desempenha um papel secundário no sistema visual e na compreensão da linguagem (John W. Clark, 2010; Kandel et al., 2000).
O lobo temporal encontra-se na parte inferior do cérebro e é delimitado pela fissura lateral. Este lobo é bastante importante no sistema auditivo através do córtex auditivo primário. O córtex auditivo secundário tem como função associar-se a outras áreas cerebrais de forma a possibilitar o reconhecimento dos sons. Pelo facto de englobar o hipocampo e a amígdala, desempenha também um papel importante nos processos de aprendizagem, de memória e das emoções (John W. Clark, 2010; Kandel et al., 2000).
Por fim, o lobo occipital situa-se na parte mais posterior do cérebro e é delimitado pelos lobos parietal e temporal. Para além do papel importante no sistema visual (córtex visual primário), possui também funções associativas como por exemplo o reconhecimento facial (córtex visual secundário) (John W. Clark, 2010; Kandel et al., 2000).
O cérebro é responsável por diversas funções executivas, sendo que aqui apenas serão introduzidas a flexibilidade cognitiva, atenção e memória de trabalho, por serem fundamentais para um bom desempenho no teste de cartas de Wisconsin.
A flexibilidade cognitiva é um mecanismo importante na resolução de problemas complexos como por exemplo a realização simultânea de múltiplas tarefas, descoberta de novas soluções para um problema ou adaptação da solução tendo em visto as novas exigências do problema (Ionescu, 2012). Esta função executiva tem sido, ao longo dos anos, associada a uma cognição humana superior, ou seja, a uma inteligência e criatividade maior (Boroditsky, Neville, Karns, Markman, & Spivey, 2010; Deak, 2003; Jordan & Morton, 2008). Vários estudos associam a região anterior do cérebro à flexibilidade cognitiva, especificamente à região do córtex cingulado anterior e do córtex pré-frontal (Botvinick, Cohen,
& Carter, 2004; Cerqueira, Almeida, & Sousa, 2008; O’Reilly, 2006; Rougier, Noelle, Braver, Cohen, & O’Reilly, 2005; Woodward, Ruff, & Ngan, 2006).
A atenção refere-se à capacidade seletiva de um humano se focar na deteção e resposta a um dado estímulo, em detrimento de outros. Considera-se usualmente a existência de dois tipos de atenção, a atenção “top-down”, onde as respostas atencionais são provocadas por uma expectativa prévia do sujeito para um estímulo relevante, e a atenção “bottom-up”, onde são os atributos físicos de um estímulo saliente que direcionam a atenção do sujeito. Vários mecanismos de atenção têm sido principalmente associados ao córtex pré-frontal (Benchenane, Tiesinga, & Battaglia, 2011; Katsuki & Constantinidis, 2012; E. K. Miller & Cohen, 2001; Pollmann, 2004; Schafer & Moore, 2011).
A memória de trabalho refere-se a um armazenamento e manipulação de curto-prazo de informação sensorial, com duração de apenas alguns segundos. Pode ser contextualizada, portanto, como um recurso limitado com o objetivo de manter em memória um determinado número de “objetos” enquanto se realiza manipulação, atualização ou aprendizagem sobre os mesmos (Baddeley, 2010; Ma, Husain, & Bays, 2014). Nas últimas décadas, a quantidade de itens que a memória de trabalho consegue armazenar tem sido alvo de muita discussão (Cowan, 2001; G. A. Miller, 1956), mas parece ser a qualidade das representações temporárias, e não a quantidade, a determinar o desempenho desta função (Ma et al., 2014). Além disto, vários estudos demonstram que o desempenho desta função cognitiva é comprometida pelo envelhecimento (Nyberg et al., 2012; Salthouse, 1994). A memória de trabalho tem vindo a ser associada à atividade neuronal em duas regiões cerebrais principais: córtex pré-frontal e córtex parietal posterior, sendo que a primeira parece estar mais relacionado com os mecanismos atencionais da memória de trabalho e a segunda com os mecanismos de armazenamento da informação (Cerqueira, Mailliet, Almeida, Jay, & Sousa, 2007; Linden et al., 2003; Todd & Marois, 2004; Xu & Chun, 2006). Vários estudos relatam ainda a interação entre memória de trabalho e atenção (Awh, Vogel, & Oh, 2006; Curtis, 2006; Linden et al., 2003), tendo sido demostrado que existem vias neuronais recíprocas a interligar ambos os mecanismos (Schwartz & Goldman-Rakic, 1984).
2.3 Eletroencefalografia
O termo eletroencefalografia (EEG) refere-se ao estudo dos potenciais elétricos, adquiridos através de elétrodos situados no couro cabeludo, que percorrem o córtex cerebral durante as excitações sinápticas das dendrites dos neurónios. Aquando de um potencial de ação nas células nervosas, existem correntes iónicas entre os meios intracelular e extracelular Estas correntes extracelulares criam assim potenciais elétricos, normalmente com frequência abaixo dos 100 Hz, que quando somadas para um grande número de células dá origem ao EEG. As mesmas correntes criam também campos magnéticos que são mensuráveis através de um magnetoencefalograma (MEG) (Sanei, 2013; Schomer & Silva, 2012; Webster, 2009).
A eletroencefalografia, tal como hoje é conhecida, foi resultado de um longo processo de evolução que dura há cerca de 139 anos. O primeiro fenómeno eletroencefalográfico foi apresentado pelo inglês Richard Caton em 1875, quando demonstrou em vários animais (coelhos, gatos e macacos), com recurso a um galvanómetro e dois elétrodos na superfície do crânio, que a agulha deste se movia devido ao fenómeno de EEG. O primeiro a demonstrar o fenómeno nos humanos foi o neuropsiquiatra alemão Hans Berger, em 1929, que gravou em papel fotográfico sinais eletroencefalográficos com a duração de 1-3 minutos. Ainda em 1929, Berger publicou que o ritmo alfa era a componente mais significativa de
um sinal eletroencefalográfico. Nos anos seguintes Berger desenvolveu bastante trabalho em várias
condições/distúrbios cerebrais (tais como a hipoxia, epilepsia, sono ou anestesia) e concluiu que o estado elétrico cerebral dependia de tais condições. De salientar que o mesmo neuropsiquiatra se interessou também na localização de tumores cerebrais através desta técnica. Um marco importante para a evolução da técnica foi o reconhecimento, por parte de Kornmuller, da importância das aquisições multicanal e do uso de um elevado número de elétrodos para avaliar uma maior área cerebral. Apesar
das conclusões dos estudos de Berger sobre vários distúrbios cerebrais, a pessoa considerada como
pioneira da utilização da eletroencefalografia na área clínica é o inglês W. Gray Walter. Walter verificou que os elétrodos mais próximos de um tumor cerebral apresentavam atividade elétrica anómala e ainda descobriu a origem cerebral da atividade elétrica de baixa frequência (ondas delta) (Sanei, 2013; Schomer & Silva, 2012).
Os sistemas de aquisição de EEG modernos são inteiramente distintos dos primeiros sistemas de EEG utilizados, em que os seus principais componentes são: elétrodos, amplificadores, módulo de controlo, armazenamento e processamento (computador) e um monitor para monitorização dos sinais adquiridos. Talvez a diferença mais notória entre ambos os sistemas seja a digitalização de sinal observada nos sistemas mais atuais. Os sinais analógicos, depois de amplificados, são convertidos para
digital através de conversores analógico-digital (ADC). A partir deste momento, os sinais são armazenados
em memórias e poderão ser processados por software especialmente desenvolvido para o efeito,
podendo obter-se uma grande variedade de resultados (Sanei, 2013; Schomer & Silva, 2012).
Os elétrodos utilizados nos sistemas modernos de EEG, dependendo do grau de invasão corporal, podem ser classificados em três tipos: não-invasivos, invasivos e minimamente invasivos. Os elétrodos não-invasivos são aqueles que são dispostos no couro cabeludo após limpeza cuidada do mesmo. Estes elétrodos consistem normalmente em discos até 3 milímetros de diâmetro e são produzidos em prata com cloreto de prata (Ag/AgCl) ou ouro (Au). Para este tipo de elétrodos é necessária uma amplificação elevada do sinal e um bom acoplamento das impedâncias (geralmente com recurso a aplicação de gel condutor). Os elétrodos invasivos são usualmente denominados de corticais e consistem em finas agulhas que são implantadas diretamente no córtex cerebral em operações minimamente invasivas. Os elétrodos minimamente invasivos situam-se intermediamente em comparação com os elétrodos anteriores pois apenas penetram a camada superficial da pele (stratum corneum), evitando assim a resistência desta camada à condução elétrica. Os elétrodos utilizados em eletroencefalografia não-invasiva podem ainda ser divididos em ativos ou passivos. Os elétrodos ativos são aqueles que possuem um pré-amplificador e ainda alguma filtragem inseridos no próprio elétrodo. Por outro lado, os elétrodos passivos não possuem qualquer tipo de eletrónica embutida no mesmo. A maioria dos sistemas de EEG utiliza elétrodos passivos pois o seu custo de produção e manutenção é menor em comparação com os ativos. Porém, é necessária uma preparação cuidada do local onde o elétrodo será colocado, normalmente com o uso de géis eletrolíticos com o intuito de diminuir a impedância entre o elétrodo e a pele. Os elétrodos ativos, pelo facto de possuírem uma interface capacitiva de impedância elevada, não necessitam de uma preparação tão extensa como no caso dos elétrodos passivos. Pelo facto da impedância da pele (seca, ou seja, sem géis) ser bastante alta, esta pode ser negligenciada com o uso de uma impedância de entrada alta no pré-amplificador. Estes elétrodos são normalmente produzidos recorrendo a materiais altamente dielétricos (como os óxidos metálicos). O uso deste tipo de elétrodos permite uma razão sinal-ruído mais elevada, melhorando assim a qualidade das aquisições. Outra razão a favor dos elétrodos ativos é a segurança devida à impedância bastante elevada da barreira pele/elétrodo (Dias, 2009; Sanei, 2013; Schomer & Silva, 2012).
O sinal de EEG, desde que se origina nos neurónios até á sua leitura nos elétrodos, percorre uma grande quantidade de camadas resistivas, tais como o crânio, o couro cabeludo e ainda várias camadas intermédias de pele. O córtex cerebral, a camada mais exterior do cérebro, tem uma espessura de
0,1-resistividade de 10-25 kΩ, sendo a camada que mais se opõem à condução elétrica (o crânio atenua o sinal cerca de mil vezes mais que o tecido mole). Por fim, o couro cabeludo tem uma espessura de 0,2-0,5 cm e uma resistividade de 300-400 Ω. O efeito de todas estas camadas faz com que o sinal seja fortemente atenuado quando chega ao elétrodo, fazendo com que seja necessários amplificadores para tornar o sinal adequado para futuro processamento e análise. Existe ainda a atenuação originada pelas propriedades condutoras dos elétrodos e pela interface destes com o couro cabeludo (Sanei, 2013).
Os amplificadores para além de amplificarem os sinais elétricos desejados, também amplificam ruído (sinal indesejado). O ruído pode ter duas fontes: interna ou externa. O ruído interno também se poderá denominar de ruído fisiológico pois é obtido maioritariamente de movimentos corporais do sujeito. Como ruído fisiológico considera-se o movimento ocular (EOG), o movimento muscular (EMG, maioritariamente dos músculos da cabeça e da face), a interferência do ECG e ainda o efeito cardiobalístico (BCG). Podem ainda encontrar-se outros artefactos fisiológicos como o efeito da respiração, da língua e ainda os potenciais gerados pelas glândulas sudoríparas. O ruído externo pode ter várias origens como outros dispositivos elétricos que se encontram perto do sistema, o ruído gerado pela alimentação elétrica do sistema de EEG (50 Hz), flutuações nas impedâncias, possíveis defeitos nos cabos, entre outros (Sanei, 2013; Schomer & Silva, 2012; Webster, 2009).
Perante as atenuações na amplitude de sinal (a amplitude do sinal que é lido nos elétrodos, sem qualquer tipo de amplificação, ronda os 1-100 μV) e as fontes de ruído supracitadas, a amplificação e o acondicionamento do sinal são fases muito importantes para aumentar a razão sinal-ruído (SNR), de forma a poder fazer-se a leitura e análise corretas dos sinais. Os sinais de EEG possuem componentes de frequência que podem atingir os 300 Hz em condições de ausência de ruído externo. Frequentemente, filtros analógicos ou digitais passa-alto a 0,5 Hz, passa-baixo a 60 Hz e Notch com frequência nula de 50 Hz são usados em gravações de EEG. O filtro passa-alto rejeita as muito baixas frequências como por exemplo a componente de frequência da respiração. O filtro passa-baixo rejeita as altas frequências que são consideradas não pertencer à atividade cerebral. O filtro Notch tem o objetivo de rejeitar a frequência da alimentação elétrica do sistema. Sendo que as frequências de interesse num EEG rondam muitas vezes os 30 Hz, pela teoria de Nyquist (Sanei, 2013), a frequência de amostragem do sinal tem de ser no mínimo 60 Hz. As frequências de amostragem usualmente utilizadas são de 250, 500 e 1000 amostras por segundo (Sanei, 2013).
Dois conceitos também bastante importantes quando se adquire e analisa sinais de EEG são a resolução temporal e a resolução espacial dos sinais adquiridos. A técnica de eletroencefalografia possui uma resolução temporal elevada, quando comparada por exemplo, com a ressonância magnética
funcional ou o tomografia por emissão de positrões (PET). Por um lado, a resolução temporal do EEG é dada pela frequência de amostragem que, como foi referido anteriormente, ronda as 500 amostras por segundo, ou seja, uma resolução temporal de 2 milissegundos, razão pela qual é considerada uma técnica eficiente para estudar as dinâmicas temporais do cérebro. Por outro lado, a resolução espacial do EEG é dada pela quantidade de elétrodos e pela sua disposição no couro cabeludo, o que mesmo para uma touca de elevada densidade de elétrodos (128 elétrodos, por exemplo), apenas se consegue uma resolução espacial na ordem dos centímetros. Comparativamente, a ressonância funcional consegue possuir uma resolução espacial maior, na ordem dos milímetros. (Huettel, Song, & McCarthy, 2004).
A forma como se adquire os sinais, ou seja, a relação entre sinais provenientes dos elétrodos, também pode ser dividia em dois tipos: o registo referencial e o registo bipolar ou diferencial. No registo referencial, uma das entradas do amplificador diferencial é a do elétrodo em questão enquanto a outra tem origem num elétrodo de referência. Existem várias localizações para o elétrodo de referência encontradas na literatura: a localização “Cz” do sistema de localização convencional 10-20, a média de dois elétrodos nos lóbulos das orelhas, a média de dois elétrodos nos mastoides, um elétrodo na mão ou na perna ou ainda um elétrodo situado na ponta do nariz (Swartz Center for Computational Neuroscience, 2014) . Existe ainda a referência média que se baseia na média dos sinais de todos os elétrodos. A escolha da referência e sua localização merecem cautela pois caso esta não seja neutra há possibilidade do aparecimento de distorção topográfica. No registo bipolar, ambas entradas do amplificador diferencial são de dois elétrodos. Desta forma, o amplificador é capaz de traduzir os sinais de entrada e cancelar aqueles que são idênticos entre os dois elétrodos, ou seja, obtemos como resultado a diferença de atividade elétrica entre ambos elétrodos (Sanei, 2013).
Existem dois tipos principais de dados que se retiram dos sinais de EEG: ritmos nas diferentes bandas de frequência e potenciais evocados (event-related potential - ERP), a partir dos quais podem ser extraídos vários tipos de características e resultados.
Em pessoas saudáveis, os ritmos cerebrais podem ser usados para distinguir níveis de alerta desde a sonolência até elevados níveis de atenção (Sanei, 2013). As amplitudes de um sinal de EEG pode atingir 100 µV quando o elétrodo está posicionado no couro cabeludo, enquanto as frequências destes sinais vão desde os 0,5 a 100 Hz. Porém, tal como referido anteriormente, estes valores são dependentes do estado do sujeito. Frequentemente são analisados 5 ritmos de EEG, divididos de acordo com os seguintes limites de frequências (Sanei, 2013):
Teta: 4 – 7,5 Hz; Alfa: 8 - 13 Hz; Beta: 14 - 26 Hz; Gama: 30 - 80 Hz;
Uma grande parte dos autores utiliza estes limites para os diferentes ritmos, porém existem outros que discordam com a extensão destas bandas (Sanei, 2013). Na Figura 5 é ilustrada a decomposição de um sinal de EEG nos quatro principais ritmos cerebrais.
Figura 5. Decomposição de um sinal de EEG nos quatro principais ritmos cerebrais (delta, teta, alfa e beta). Adaptado de (Sanei, 2013).
Para além dos ritmos cerebrais, podem também ser extraídos os ERP. Os ERPs são uma resposta do EEG a uma determinada estimulação e têm sido importantes como indicadores de diagnóstico nas diversas áreas, desde a psiquiatria, neurologia ou como marcadores numa interface cérebro-computador (Sanei, 2013). Um ERP é uma sequência de componentes com amplitudes positivas ou negativas que refletem vários processos sensoriais, cognitivos ou motores. Ou seja, é a resposta elétrica direta do córtex cerebral a eventos sensoriais, cognitivos ou motores e normalmente com origem externa. Em termos
fisiológicos, estes potenciais positivos ou negativos são somas de vários potenciais de ação que estão sincronizados com determinado evento. Dependendo de como o sistema sensorial recolhe os estímulos, os ERP podem ser classificadas como visuais, auditivos ou somatossensoriais (Luck, 2005; Sanei, 2013). Comparando a amplitude destes potenciais com os ritmos do sinal de EEG, pode-se afirmar que esta é bastante pequena (frequentemente menos de metade da amplitude). Devido a este facto, usualmente
calculam-se ERPs através da média de vários segmentos de EEG em resposta a um estímulo que se
pretende estudar.
Habitualmente são extraídas 3 características distintas de uma componente do ERP: amplitude, latência e distribuição topográfica. A amplitude fornece informação acerca da extensão da ativação neuronal, ou seja, qual o grau de resposta de determinada região cerebral a dado estímulo. Como amplitude considera-se a diferença entre o maior pico positivo da componente e o valor médio da baseline antes do estímulo. A latência dá-nos informação da rapidez com que a ativação foi executada (nesta medição considera-se o maior pico da curva da componente) em relação ao momento do estímulo. A distribuição topográfica fornece-nos um padrão de ativação neuronal no escalpe, isto é, em cada instante mostra-nos a amplitude das voltagens para cada região cortical (Luck, 2005; John Polich, 2007; Sanei, 2013).
A classificação das componentes do ERP é feita, normalmente, tendo em conta dois fatores. Caso o sinal tenha voltagem positiva, a classificação começa com a letra P e caso apresente voltagem negativa, classifica-se com a letra N. O segundo fator corresponde à latência do pico do sinal, em milissegundos. Exemplificando, um sinal com voltagem negativa e com o pico 100 milissegundos após o estímulo classifica-se como componente N100.
As componentes de ERP, principalmente a latência e a amplitude, têm sido utilizadas como
indicadores da função do sistema neuroquímico cerebral ou como indicadores de processos cognitivos (John Polich, 2007; Sanei, 2013). Por exemplo, num estudo realizado em 2007 (resultado ilustrado na Figura 6) é possível distinguir entre duas condições, pessoas alcoólicas e não alcoólicas, através da
amplitude de uma componente do ERP, em resposta a uma tarefa visual do tipo oddball. Os sujeitos
Figura 6. Diferenças numa componente do ERP entre sujeitos alcoólicos e não alcoólicos. Adaptado de (Rangaswamy et al., 2007).
Outro estudo explorou o efeito da fadiga mental nas diferenças de amplitude num ERP em 40 sujeitos,
através de uma tarefa auditiva do tipo oddball (Sanei, 2013 - Figura 7).
Figura 7. ERP médio de 40 sujeitos com fadiga mental e antes de possuírem esta condição. Adaptado de (Sanei, 2013).
É percetível que quando os sujeitos estão mentalmente fatigados há a tendência para a latência das
componentes do ERP serem menores. A mesma tendência parece acontecer com a amplitude destas
componentes, a qual é menor quando os sujeitos estão fatigados.
O primeiro estudo que reporta a gravação de ERPs sensoriais em humanos foi em 1935-1936 por Pauline e Davis (Luck, 2005). Desde essa data até 1964 pouco trabalho foi desenvolvido nesta área devido maioritariamente à segunda guerra mundial. A era moderna da investigação dos ERPs tal como conhecemos hoje foi iniciada em 1964 por Grey Walter quando este relatou a aquisição do primeiro ERP com origem cognitiva (Luck, 2005). O próximo grande avanço nesta área foi a descoberta da componente
P300 do ERP um ano depois da aquisição de Grey Walter (Sutton, Braren, Zubin, & John, 1965). No
estudo realizado, os autores reparam que os sujeitos apresentavam uma componente sempre que não conseguiam prever se o próximo estímulo a ser apresentado era visual ou auditório, e que essa
componente possuía uma voltagem positiva e com o pico do sinal 300 milissegundos após o estímulo. A esta componente nomearam de P300 (Luck, 2005).
O impacto que a descoberta da componente P300 teve na investigação foi impressionante, com mais de 20 mil artigos publicados sobre esta temática ou relacionados com a mesma. Juntando a estes números, o artigo original de Sutton et al. já foi citado cerca de 8 mil vezes (Luck, 2005). A componente P300 é, portanto, uma componente que tem atraído muita atenção e muito dinheiro. Esta componente está ilustrada na Figura 8.
Figura 8. Subcomponentes P3a e P3b da componente P300 do ERP. Adaptado de (John Polich, 2007).
Os estudos realizados após Sutton et al. centraram-se bastante nos paradigmas do tipo oddball e mostraram que as características desta componente refletem bastante a probabilidade de aparecimento do estímulo, bem como a relevância do mesmo (John Polich, 2007). Esta componente é ainda considerada um potencial endógeno pois a sua ocorrência não está relacionada com os atributos físicos do estímulo, mas sim como a pessoa reage a esse estímulo. As componentes com latência inferior a 100 milissegundos após o estímulo são consideradas exógenas e são normalmente influenciadas pelos atributos físicos do estímulo (intensidade, tipo, taxa de apresentação, entre outros). As componentes endógenas aparecem depois dos 100 milissegundos e as suas características variam de acordo com a reação das pessoas aos estímulos (John Polich, 2007; Sanei, 2013). No caso do P300, pensa-se que esta componente reflita os processos envolvidos na avaliação e categorização dos estímulos.
A subcomponente P3b surge após um estímulo raro ou relevante num conjunto de estímulos frequentes e/ou irrelevantes. A sua latência situa-se entre os 300-600 ms após o aparecimento do estímulo e possui uma distribuição topográfica na região parietal. A subcomponente P3a reflete a orientação automática da atenção para um estímulo que seja saliente, mesmo não sendo relevante. O
P3a possui uma latência menor que a subcomponente P3b e a sua distribuição topográfica é mais anterior (em regiões centro-frontais) (John Polich, 2007).
O modelo atualmente aceite na área da investigação para a componente P300 foi proposta por Donchin em 1981 (John Polich, 2007). O modelo da atualização contextual, como é denominada, assenta na teoria de que a componente P300 indica a atividade cerebral respeitante à revisão e atualização de uma representação mental, sempre que um novo estímulo aparece (John Polich, 2007). O fluxograma da Figura 9 ilustra os processos neuronais desde o aparecimento do estímulo até á resposta neuronal.
Figura 9. Modelo da atualização contextual. Adaptado de (John Polich, 2003).
Este modelo explica que após um processamento sensorial inicial (componentes N100, P200 e N200), existe um processo de comparação atencional implementado na memória de trabalho onde se avalia a representação do estímulo anterior, comparando-a com o novo estímulo. Se os atributos do novo estímulo forem iguais à representação existente em memória, esta não é atualizada e apenas as componentes do ERP sensoriais são adquiridas. Caso o novo estímulo seja diferente, processos atencionais fazem com que seja feita uma atualização da representação presente na memória de trabalho e a componente P300 é também adquirida (John Polich, 2007).
A amplitude do P300 parece refletir as características do estímulo (a sua saliência, entre outras características físicas do mesmo), bem como a sua probabilidade de aparecimento (quanto mais improvável, maior a amplitude do P300). Esta característica do P300 também é influenciada pelas
exigências cognitivas durante a tarefa, visto que a solicitação de recursos atencionais desempenha um papel importante na geração da componente (John Polich, 2007). Por sua vez, a latência da componente P300 é proporcional ao tempo necessário pelo cérebro para a avaliação do estímulo. A latência é influenciada pelas capacidades cognitivas do sujeito (a idade do mesmo é um fator determinante), bem como pelas exigências de processamento requeridas pela tarefa (John Polich, 2007).
Para além de estudos eletroencefalográficos, os ERP também podem ser estudados a partir de
uma técnica imagiológica como a ressonância magnética funcional. Porém, esta técnica apresenta
algumas desvantagens em relação à eletroencefalografia na aquisição dos ERP cerebrais: a resolução
temporal é baixa, apesar de ser possível utilizar um paradigma de eventos; as áreas ativadas nesta técnica são baseadas na hemodinâmica, o que nem sempre corresponde às áreas neuronais
responsáveis pelos ERP; por último, o fMRI não consegue ter sensibilidade suficiente para distinguir o
tipo de estímulo apresentado (estímulo alvo, estímulo irrelevante ou um novo estímulo) (Sanei, 2013).
2.4 Ressonância Magnética Funcional
A ressonância magnética funcional (fMRI) é uma técnica de neuroimagiologia funcional que utiliza a tecnologia da ressonância magnética para medir alterações na atividade cerebral. Esta técnica imagiológica tem a capacidade de detetar variações, em relação ao estado basal, do fluxo sanguíneo em determinada região cerebral.
A primeira máquina de ressonância magnética foi desenvolvida em meados do século XX pelo
estudante de doutoramento Herman Carr (Carr, 1953), porémo grande avanço para o desenvolvimento
da técnica da ressonância magnética nuclear foi feito por Pauli, em 1924, quando este sugeriu que os núcleos atómicos possuíam spins magnéticos (Buxton, 2009). Nos anos seguintes esta hipótese proposta por Pauli foi testada, independentemente, por Rabi, em 1938, que conseguiu desenvolver um método de deteção dos spins magnéticos em gases (Huettel et al., 2004); por Purcell e Bloch que concluíram que a deteção e medição dos spins magnéticos também seria possível em sólidos (Buxton, 2009). Foi então que, em 1953, o estudante Herman Carr desenvolveu a primeira máquina que permitia criar estas imagens (Carr, 1953; Wehrli, 1992). Outros cientistas, nos anos seguintes, desenvolveram máquinas idênticas: Vladislav Ivanov (MacWilliams, 2003) em 1960 ou Raymond Damadian em 1971. Em 1974, Damadian obteve a patente da primeira máquina de ressonância magnética suficientemente grande para conter um humano (Damadian, 1974).
A ressonância magnética é uma técnica de imagiologia, não necessariamente neuronal, que permite, através de fortes campos magnéticos, criar imagens de tecidos biológicos. É, portanto, um instrumento não-invasivo que permite estudar a anatomia e fisiologia de seres vivos (Prasad, 2006). O princípio fundamental desta técnica assenta no fenómeno da ressonância magnética nuclear, onde os núcleos atómicos expostos a um campo magnético forte absorvem e emitem, posteriormente, ondas eletromagnéticas a uma dada frequência (frequências rádio). Pelo facto de usar campos magnéticos e ondas rádio, aos quais não se atribuem quaisquer efeitos nocivos, esta técnica é considerada bastante segura para estudos em humanos quando comparada com outras técnicas de imagiologia como o raio-X ou a tomografia computorizada, que usam radiações ionizantes (Prasad, 2006). O sinal medido por uma máquina de ressonância magnética baseia-se nas propriedades magnéticas do núcleo dos átomos de hidrogénio existentes nos tecidos. Aliás, 70% do nosso corpo é composto por água, sendo que esta possui dois átomos de hidrogénio, podendo por isso utilizar-se a técnica de ressonância magnética em qualquer região corporal. O núcleo de hidrogénio é composto apenas por um protão, não possuindo neutrões. Pelo facto de este protão ser uma partícula carregada e giratória, este produz um campo magnético normalmente denominado de momento magnético. Habitualmente estes protões estão orientados ao acaso, fazendo com que não exista qualquer campo magnético médio bem definido (Mulert & Lemieux, 2010; Prasad, 2006).
Os componentes principais de uma máquina de ressonância magnética são o campo magnético primário, os campos magnéticos de gradiente e ainda os pulsos de radiofrequência (Mulert & Lemieux, 2010; Prasad, 2006).
O campo magnético primário, 𝐵0, produzido pela máquina de ressonância magnética é estático,
permanente e tem um valor fixo. Esse valor é medido em Tesla (T), sendo que as máquinas de ressonância magnética utilizadas na prática clínica possuem campos magnéticos na ordem dos 1,5T a 7T. Este valor produz um campo magnético bastante forte, superiorizando-se ao do planeta Terra cerca de 100000 vezes (utilizando um campo magnético de 3T). Na área de investigação já se utilizam máquinas com campos magnéticos mais fortes que os 7T. A partir do efeito do campo magnético primário, os átomos de hidrogénio orientam-se, na sua maioria, paralelamente (para um estado de baixa
energia) em relação a este campo, criando um vetor de magnetização 𝑀0 com a mesma direção e
sentido do campo magnético primário. Esta orientação denomina-se de magnetização longitudinal. Um fenómeno também importante e que acontece simultaneamente é o de precessão, que se define como sendo um movimento oscilante que ocorre quando um objeto giratório é sujeito a uma força externa. Quando os protões estão sujeitos ao campo magnético primário, estes precessam sobre o seu eixo a
uma dada frequência angular, a frequência de Larmor 𝜔𝐿. De salientar ainda que esta frequência altera-se consoante a força do campo magnético prealtera-sente (por exemplo, para um campo magnético de 1,5T,
a frequência de Larmor é de 63,9 MHz) (Mulert & Lemieux, 2010; Prasad, 2006). Este fenómeno de
precessão encontra-se ilustrado na Figura 10.
Figura 10. Precessão de uma partícula atómica giratória na presença do campo magnético primário 𝐵0. Adaptado de (Prasad, 2006).
Os campos magnéticos de gradiente atuam sobre o campo magnético primário, alterando as suas propriedades, e dispõem-se, relativamente a este campo magnético, sobre 3 diferentes eixos (x, y e z) de forma a poder obter-se uma imagem tridimensional. Cada um dos campos magnéticos de gradiente é criado por uma bobine na direção do eixo pretendido. Estes campos magnéticos são os responsáveis pelo som bastante ruidoso que se ouve durante uma aquisição de ressonância magnética. Com os campos magnéticos de gradiente é então possível fazer uma determinada seleção de fatias (slices), ou seja planos de corte, e ainda localização de acordo com os eixos ortogonais. De acordo com o ilustrado
na Figura 11, planos de corte perpendiculares ao eixo ortogonal z permitem criar imagens de planos
axiais, planos de corte perpendiculares ao eixo ortogonal y permitem criar imagens de planos coronais e planos de corte perpendiculares ao eixo ortogonal x permitem criar imagens de planos sagitais (Mulert & Lemieux, 2010; Prasad, 2006).