UMBUZEIRO (SPONDIAS TUBEROSA): A SYSTEMATIC REVIEW
Texto
Imagem

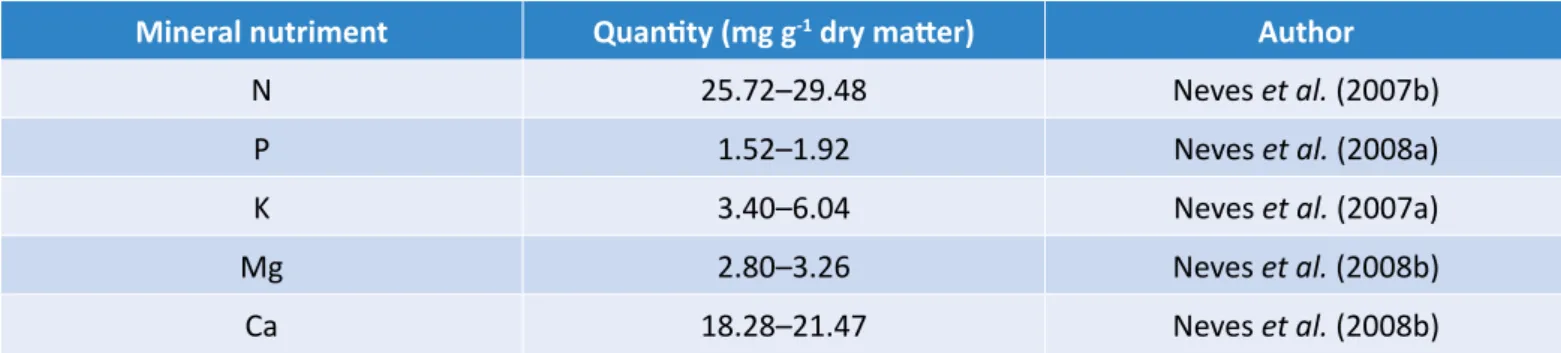
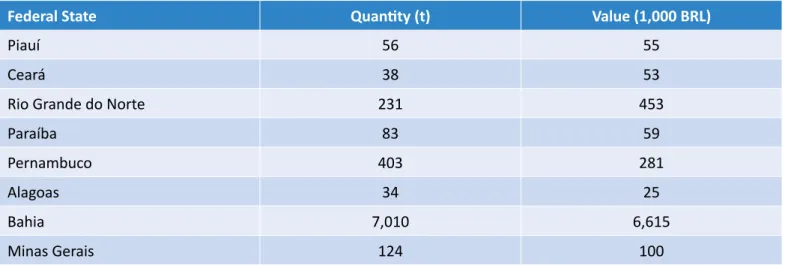
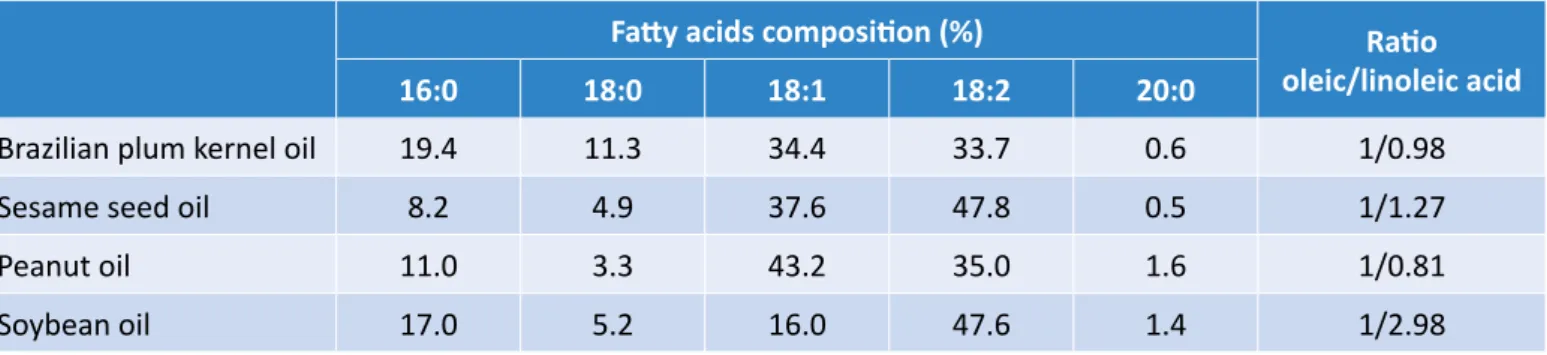
Documentos relacionados
Capella (1998), com base em uma pesquisa realizada com a metodologia construtivista, chega à formulação de um modelo capaz de romper com essa forma
Resíduos de mistura de betão, ladrilhos, telhas e materiais cerâmicos foram produzidos praticamente durante todo o período de edificação. Variação da produção
Os modelos desenvolvidos por Kable & Jeffcry (19RO), Skilakakis (1981) c Milgroom & Fry (19RR), ('onfirmam o resultado obtido, visto que, quanto maior a cfiráda do
Com expressivos valores de exportação de carne de frango brasileira torna- se evidente a necessidade da geração de previsões de vendas destes produtos, para o mercado externo, bem
Para estabelecer a presença ou ausência de DTM, foi utilizado o Research Diagnostic Criteria for Temporomandibular Disorders (RDC/TMD); além de registrar a
O candidato selecionado oriundo de país não lusófono onde não ocorra aplicação do exame Celpe-Bras deverá freqüentar, durante o ano letivo de 2015, curso de língua
Varnhagen, como já nos referimos reiteradas vezes, não fabrica sua História Geral do Brasil, no intuito de “apenas” narrar o passado; seus objetivos são mais amplos, pois
A Figura 5.24, referente a amostra dopada com cério, mostra que a borda de absorção se desloca para alta energia com o aumento da pressão.. Isso sugere que a o rênio nesse
