Efeitos da dimensão e distância de fragmentos de habitats não-matriz para a diversidade de mamíferos do montado de sobro da Serra da Grândola
51
0
0
Texto
(2) ÍNDICE. Agradecimentos. 3. Resumo. 4. Summary Introdução. 7. Região mediterrânica Montado Heterogeneidade da paisagem e qualidade do habitat Pequenos mamíferos Carnívoros Objectivos Área de estudo. 14. Metodologia. 17. Pequenos mamíferos Carnívoros Análise de dados Resultados. 20. Riqueza específica e diversidade Pequenos mamíferos Carnívoros Discussão dos resultados. 33. Riqueza específica e diversidade Pequenos mamíferos Carnívoros Considerações Finais. 38. Bibliografia. 40. 2.
(3) AGRADECIMENTOS. À orientadora, Professora Doutora Margarida Santos-Reis por tornar possível a realização deste projecto, pela disponibilidade, apoio e transmissão de conhecimentos.. Ao CBA por permitir a utilização das suas instalações na estação de campo da Herdade da Ribeira Abaixo.. À Sara pela companhia, apoio e amizade.. Ao Miguel Rosalino pela disponibilidade e ajuda.. Ao João pela paciência e ajuda preciosa no campo.. Ao Marco e ao Fernando pela ajuda no campo.. À Alice e restantes companheiros da herdade pela animação.. 3.
(4) RESUMO. A bacia do Mediterrâneo é considerada um “hotspot” da biodiversidade mundial devido à sua elevada riqueza específica e à presença de espécies endémicas, em particular, plantas e vertebrados (Myers et al. 2000). O montado é um ecossistema tipicamente mediterrânico e constitui o maior sistema agro-silvo-pastoril da Europa, sustentando uma elevada biodiversidade (Diaz et al. 1997). A heterogeneidade e configuração da paisagem, juntamente com a qualidade do habitat são condições determinantes na distribuição, diversidade e estrutura das comunidades faunísticas. A fragmentação do habitat, aliada à redução da heterogeneidade, constitui, actualmente, uma das maiores ameaças à conservação da biodiversidade (Wilcox & Murphy 1985, Paniagua et al. 2004). O objectivo deste trabalho foi avaliar a participação dos vários tipos de habitat na diversidade e abundância de mamíferos na paisagem do montado. Pretendeu-se analisar se a existência de fragmentos de diferentes tipos de habitat teria algum efeito sobre a abundância e diversidade de mamíferos; e também se a utilização das manchas pelos mamíferos estaria relacionada com o tipo de habitat, a distância entre manchas e a dimensão das mesmas. Em geral, pode-se concluir que o montado por si só não garante a conservação de mamíferos em ecossistemas mediterrânicos, são os fragmentos de habitat não-matriz que asseguram a abundância e diversidade das espécies. Constatou-se que as galerias ripícolas dão um importante contributo para a riqueza específica e diversidade de mamíferos, pelo que, é fundamental contemplar este tipo de habitat nos planos de conservação em regiões mediterrânicas. No sentido de conservar a diversidade de mamíferos típica do montado é ainda necessário considerar o tamanho e distância entre as manchas de habitat não-matriz, uma vez que é fundamental maximizar a área (em especial das manchas cultivadas) e minimizar a distância aos restantes fragmentos.. 4.
(5) Palavras-chave: Biodiversidade; heterogeneidade; pequenos mamíferos; carnívoros; montado. 5.
(6) SUMMARY. The Mediterranean basin is considered a world biodiversity hotspot due to its high specific richness and to the presence of endemic species, particularly plants and vertebrates (Myers et al. 2000). The montado is a mediterranean environment and constitutes the largest system of lands for agriculture, forestry and pasture in Europe, sustaining a high level of biodiversity (Diaz et al. 1997). The landscape heterogeneity and configuration, along with habitat quality play determinant roles in distribution, diversity and structure of faunistical communities. The habitat fragmentation, along with heterogeneity reduction, is nowadays one of the biggest threat do biodiversity conservation (Wilcox & Murphy 1985, Paniagua et al. 2004). The aim of this project was to evaluate the participation of several kinds of habitat in the diversity and abundance of mammals in the montado landscape. We intended to analyse if the existence of different habitat patches would have some effect on the abundance and diversity of mammals; and if the use of the patches would be related to the habitat type, the distance between patches and its dimension. We concluded that montado on its own does not guarantee mammals conservation in mediterranean environments, the abundance and diversity of mammals are provided by the patches of non-matrix habitat. Riparian areas are important for mammals specific richness and diversity, so it is crucial to take these patches into account when promoting conservation actions in mediterranean areas. To promote the diversity of mammal species in the montado it is also important to consider the size and distance between patches of non-matrix habitat, since it is essential to maximize the area and minimize the distance to other patches.. Keywords: Biodiversity; heterogeneity; small mammals; carnivores; montado. 6.
(7) INTRODUÇÃO. Este estudo insere-se no projecto “Valor aditivo dos fragmentos não matriz. para. a. biodiversidade. do. montado. de. sobro:. implicações. conservacionistas e de gestão” (FCT - POCI/BIA-BDE/61122/2004), cujo principal objectivo é demonstrar a contribuição de fragmentos de habitat nãomatriz para a biodiversidade total das paisagens de montado de sobro.. Região Mediterrânica. A bacia do Mediterrâneo é considerada um “hotspot” da biodiversidade mundial devido à sua elevada riqueza específica e à presença de espécies endémicas, em particular, plantas e vertebrados (Myers et al. 2000). Para este facto contribuiu a elevada heterogeneidade da paisagem mediterrânica, a nível geológico, climático e fitossociológico, assim como factores históricos, tais como a ocorrência de espécies de outras regiões e a especiação in situ (Del Bove & Isotti 2001). O sudoeste da Península Ibérica é disso exemplo, uma vez que aí convergem espécies originalmente provenientes do norte da Europa e de África, sendo o limite meridional das primeiras e o limite setentrional das segundas (Lawton 1993, Borralho et al. 1995). No entanto, a forte influência humana tem contribuído para o aumento de variabilidade nas áreas mediterrânicas, consequência de mudanças drásticas causadas, por exemplo, por fogos e conversão de terrenos para exploração. agrícola,. pecuária. ou. silvícola. (Corominas. 2004),. com. consequências nefastas a nível da biodiversidade (Pereira & Fonseca 2003).. Montado. O montado é o sistema agro-silvo-pastoril mais extenso da Europa, estando contemplado no Anexo I da Directiva Habitats da União Europeia (nº 92/43/EEC), devido à elevada biodiversidade que sustenta (Diaz et al. 1997). É 7.
(8) um exemplo de integração da natureza e da sua utilização por parte do Homem (Pereira & Fonseca 2003), uma vez que consiste no maquis original que foi sendo gradualmente alterado para o tornar adequado a actividades de cariz antropogénico (Pinto-Correia 2000). Estando bem adaptado às características da região mediterrânica (Diaz et al. 2003), o montado é um dos usos do solo dominante em Portugal, especialmente no sul, podendo dividir-se em 2 tipos: montado de sobro (dominado pelo sobreiro Quercus suber) e montado de azinho (dominado pela azinheira Quercus ilex), presentes junto à costa e no interior do país, respectivamente (Pinto-Correia 2000). É um biótopo que suporta uma elevada biodiversidade, sendo esta reconhecidamente superior à suportada por ecossistemas naturais. e. humanizados em separado (Diaz et al. 2003, Plieninger et al. 2003). A justificação deste facto reside na heterogeneidade típica da paisagem do montado, com manchas de bosque, pasto e áreas cultivadas, o que permite a sobrevivência de espécies com requisitos muito diferentes (Diaz et al. 1997, Diaz et al. 2003, Plieninger et al. 2003). Aparentemente, a intervenção humana tem um papel benéfico, aumentando a variedade de recursos disponíveis. Vários estudos comprovam que o montado é uma das áreas com maior biodiversidade animal em Portugal (Santos-Reis & Correia 1999, Rosalino et al. 2005), nomeadamente, no que se refere às aves (Pereira & Fonseca 2003) e aos carnívoros (suporta 70% dos carnívoros de porte médio - Santos-Reis et al. 1999), sendo inclusivamente o biótopo onde foi encontrado o último indício de presença do felídeo mais ameaçado do mundo (Nowell & Jackson 1996), o lince (Lynx pardinus) em Portugal (Santos-Reis 2003). Uma vez que a paisagem do montado é caracterizada por um diversificado mosaico de biótopos em que a abundância de recursos é variada e sazonal, os carnívoros que vivem neste ecossistema são, maioritariamente, omnívoros (e. g. Rosalino et al. 2004, Santos et al. 2007).. 8.
(9) Heterogeneidade da Paisagem e Qualidade do Habitat. A heterogeneidade e configuração da paisagem, juntamente com a qualidade do habitat (sensu fonte de recursos para as comunidades animais) são factores determinantes na diversidade, estrutura e funcionamento das comunidades faunísticas, pelo que é essencial conhecer e prever a resposta das espécies a alterações destes factores (Law & Dickman 1998, Dauber et al. 2003, Corbalán 2005, Tabeni et al. 2005). A heterogeneidade da paisagem deve-se às características do solo, topografia e factores microclimáticos, que determinam o(s) tipo(s) de formação vegetal e, consequentemente, o tipo de habitat disponível (Law & Dickman 1998). Vários estudos têm abordado a heterogeneidade dos habitats e o seu impacto. na. diversidade,. tendo. sido. desenvolvida. a. Hipótese. da. Heterogeneidade do Habitat (Simpson 1949, MacArthur & Wilson 1967, Lack 1969), que tem como pressuposto que os habitats mais complexos suportam uma maior variedade de nichos e, consequentemente uma diversidade mais elevada (Bazzaz 1975, August 1983, Burel et al. 1998), o que foi comprovado num estudo em que se detectou uma diversidade florística mais elevada em paisagens com maior grau de heterogeneidade (Bazzaz 1975). Enquanto que algumas espécies dependem unicamente de um tipo de habitat, outras necessitam de vários recursos, por exemplo durante diferentes fases do ciclo de vida, pelo que é importante que consigam aceder a diferentes habitats numa área relativamente pequena (Law & Dickman 1998). O meio físico regula a distribuição dos animais de duas formas: constituindo a fonte de alimento e sendo o meio através do qual se dispersam. Assim, adicionalmente às características das manchas de diferentes habitats (coberto vegetal, área, grau de isolamento e tipo de solo), a composição da matriz onde estas se dispõem, também contribui para a presença das espécies, tal como demonstrado em estudos de comunidades de insectos (Dauber et al. 2003). Dependendo das características físicas e da biologia da espécie em causa, a matriz pode funcionar como barreira, aprisionando os indivíduos num fragmento de habitat adequado, ou ter qualidade suficiente para permitir a. 9.
(10) dispersão, caso inclua algumas fontes de alimento e/ou locais de refúgio (Antongiovanni & Metzger 2004, Revilla et al. 2004). A distribuição dos animais é fruto da selecção do habitat, ou seja, do mecanismo através do qual um animal escolhe entre os recursos de habitat que encontra disponíveis (Orrock et al. 2000). Segundo a Teoria da Selecção do Habitat (Fretwell & Lucas 1972), os indivíduos tendem a estabelecer-se em habitats que lhes conferem fitness e sobrevivência óptimos (Corbalán et al. 2006); assim pode-se assumir que as densidades mais elevadas ocorrem nos habitats de maior qualidade (Corbalán et al. 2006). Em paisagens muito heterogéneas alguns fragmentos de habitat podem funcionar como estruturas keystone, ou seja, locais que proporcionam recursos para determinada espécie; a identificação destas estruturas é muito útil para direccionar esforços de conservação (Tews et al. 2004). Existem evidências de que áreas cultivadas e ripícolas possam, frequentemente, desempenhar esta função (Hellegers, Virgós 2001, Marshall et al. 2006), sendo habitats aos quais os animais não estão restringidos mas que são regularmente utilizados, nomeadamente como local de alimentação e/ou refúgio (Martin 2002). A. fragmentação. do. habitat,. juntamente. com. a. redução. da. heterogeneidade, constitui, actualmente, uma das maiores ameaças à conservação da biodiversidade (Wilcox & Murphy 1985, Paniagua et al. 2004). Para isso contribui também o aumento da sensibilidade e diminuição da capacidade de resposta a alterações ambientais por parte das espécies, que é função de vários factores: redução da área de habitat adequado (Paniagua et al. 2004), aumento do grau de isolamento das manchas de habitat favorável (Paniagua et al. 2004), reduzido tamanho da população (Lande 1993), reduzida variabilidade genética e elevado inbreeding (Lacy 1993, Saccheri et al. 1998), alterações na dinâmica intra e inter-populacional (Temple & Cary 1988, McCarthy et al. 1994, Paton 1994, Lacy 2000) e alterações em processos ecológicos essenciais para a manutenção de populações viáveis (Klein 1989). Apesar da fragmentação do habitat ter, em geral, um impacto negativo, as paisagens fragmentadas oferecem um mosaico de habitats diferentes para a fauna (Gascon et al. 1999), o que pode ser benéfico, nomeadamente, para espécies generalistas, que assim têm na mesma área uma maior diversidade de recursos tróficos para explorar (Bright 1993, Virgós 2002). Andrén (1994) 10.
(11) detectou uma maior densidade de corvos (Corvus corax) em paisagens mistas de zonas cultivadas e floresta do que em qualquer dos dois habitats isoladamente. Finalmente,. a. disposição. dos. recursos. em. mosaico. afecta. o. comportamento dos indivíduos, a Optimal Foraging Theory (Emlen 1966, MacArthur & Pianka 1966) evidencia que este varia em função da fome, disponibilidade, distribuição e qualidade do alimento e que os animais tendem a agir de forma maximizar os ganhos, minimizando os custos. Adicionalmente, a Patch Foraging Theory (Hassell & Southwood 1978) demonstra que, apesar da busca de alimento se iniciar numa direcção aleatória, verifica-se uma tendência dos indivíduos para centralizar as trajectórias dos movimentos em apenas um sentido (Kareiva 1985, Fromm & Bell 1987, Bell 1990, Bell 1991, Cartar & Real 1997, Cresswell 1997, McIntyre & Wiens 1999).. Pequenos Mamíferos. Os pequenos mamíferos desempenham um papel muito importante no funcionamento dos ecossistemas, através do consumo e dispersão de sementes, por um lado, e como fonte de alimento essencial para muitos predadores, tanto mamíferos como aves, por outro (Diaz 1992, Blanco 1998, Corominas 2004, Converse et al. 2006). Apesar de ser um grupo heterogéneo, as espécies de pequenos mamíferos partilham as características biológicas definidas pelo seu tamanho reduzido, elevada taxa metabólica e territórios de pequena dimensão (Corominas 2004). A distribuição espacial e a estrutura da comunidade de pequenos mamíferos são fortemente dependentes de características do habitat, tais como a cobertura vegetal que, além de ser fonte de alimento, condiciona o refúgio e a facilidade de fuga aos predadores (Gonnet 1998, Corbalán 2005, Tabeni & Ojeda 2005, Corbalán et al. 2006). Assim, a abundância e riqueza específica dos pequenos mamíferos são superiores em habitats com elevada cobertura vegetal, pois proporcionam melhores refúgios e mais fontes de alimento, como. 11.
(12) folhas, frutos, sementes ou insectos (Kotler et al. 1994, Gonnet & Ojeda 1998, Corominas 2004). Existem estudos que comprovam que as espécies de pequenos mamíferos têm a capacidade de distinguir os principais tipos de habitat num mosaico (Gonnet & Ojeda 1998, Tabeni & Ojeda 2005) e em paisagens heterogéneas conseguem sobreviver em manchas pequenas ou percorrer grandes distâncias para encontrar habitat adequado (Burel et al. 1998). Na região mediterrânica, o número de espécies de pequenos mamíferos é reduzido, com preponderância de espécies ubíquas e generalistas (Corominas 2004). As comunidades são formadas usualmente por um insectívoro. generalista. (e.g.. Crocidura. russula),. um. generalista. granívoro/insectívoro (e.g. Apodemus sylvaticus) e um granívoro de meios abertos (e.g. Mus spretus) (Corominas 2004).. Carnívoros. Os papéis dos carnívoros no ecossistema são variados. Entre outros, funcionam como reguladores das populações de presas (Blanco 1998, Paniagua et al. 2004), podendo induzir-lhes mudanças ao nível do comportamento e nas regiões onde complementam a dieta com frutos favorecem o transporte e dispersão das sementes (Paniagua et al. 2004). Os carnívoros constituem um grupo vulnerável, encontrando-se grande parte das espécies com problemas de conservação devido à perseguição e destruição do habitat (Gittleman et al. 2001), cujos efeitos são ampliados pelo tamanho relativamente grande das espécies (Virgós et al. 2002). No entanto, o estudo e monitorização destas populações têm vários constrangimentos: Reduzida densidade, o que diminui a probabilidade de captura, além de aumentar a vulnerabilidade a acontecimentos estocásticos (Paniagua et al. 2004, Christophersen et al. 2005, Rosalino et al. 2005) Comportamento secretivo (Paniagua et al. 2004, Rosalino et al. 2005) Taxas reprodutivas reduzidas, o que resulta numa reacção lenta às alterações no ambiente (Christophersen et al. 2005) Tamanho dos territórios elevado (Christophersen et al. 2005) 12.
(13) Os carnívoros, na sua maioria, têm hábitos tróficos generalistas, alimentam-se de pequenos mamíferos, quando estes estão disponíveis, mas têm capacidade de explorar outros recursos (Corominas 2004), o que se verifica em zonas de montado, onde consomem uma grande quantidade de frutos (Santos 1998). Ainda assim, a selecção do habitat por parte dos carnívoros é claramente influenciada pela abundância de pequenos mamíferos (Murray et al. 1994).. Objectivos. O objectivo deste trabalho foi avaliar a contribuição de vários tipos de habitat para a diversidade e abundância de mamíferos na paisagem de montado de sobro típica da região mediterrânica. Pretendeu-se analisar se a existência de manchas de diferentes tipos de habitat teria algum efeito sobre a comunidade de mamíferos e ainda se a utilização das manchas pelos mamíferos estaria relacionada com o tipo de habitat, distância entre manchas e dimensão das mesmas. Finalmente, pretendeu-se compreender a importância dos fragmentos de áreas cultivadas, de forma a poder desenvolver e implementar medidas que promovam a conservação dos mamíferos em regiões com características semelhantes. Foi também objectivo confirmar a hipótese de que a ocorrência de manchas de habitat não-matriz promove a abundância e diversidade de mamíferos no ecossistema mediterrânico, sendo o seu contributo proporcional à área de cada mancha e inverso à distância entre as mesmas. Seleccionaram-se os mamíferos como modelo de estudo devido à variedade de nichos tróficos e ecológicos que realizam, à sua intervenção na sustentabilidade do montado e à sua sensibilidade a perturbações ambientais e antropogénicas.. 13.
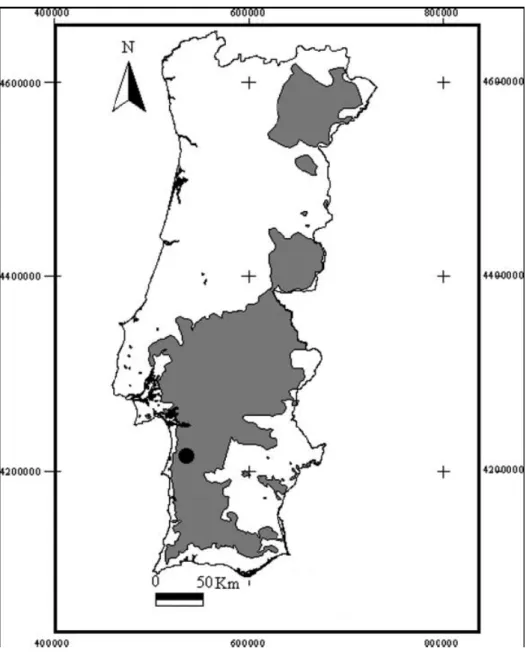
(14) ÁREA DE ESTUDO. O trabalho de campo teve lugar na Serra de Grândola. A topografia é moderada, variando as elevações entre 150 e 240m acima do nível do mar (Correia & Santos-Reis 1999). O clima característico é mediterrânico com influências atlânticas. A sazonalidade é bastante marcada (Blondel & Aronson 1999), sendo o Verão quente e seco e o Inverno frio e húmido; a temperatura e precipitação médias são 15.6ºC e 500mm/ano, respectivamente (Correia & Santos-Reis 1999). A região faz parte de uma das maiores áreas de montado de Portugal, sendo caracterizada por sobreiro (Quercus suber) ao qual se associa por vezes a azinheira (Quercus ilex) e o carvalho-cerquinho (Quercus faginea), existindo também pequenas áreas de eucalipto (Eucalyptus globulus) e pinheiro bravo (Pinus pinaster). A nível do subcoberto marcam presença a esteva (Cistus spp.), o rosmaninho (Lavandula stoechas), o medronheiro (Arbutus unedo) e a urze (Erica spp.) (Correia & Nisa 1999). Para além da matriz de montado existem fragmentos de hortas e olivais de pequenas dimensões dispersos pela paisagem, maioritariamente abandonados. Para além da oliveira (Olea europaea), encontram-se nestas manchas árvores de fruto, sendo as mais frequentes a figueira (Ficus carica), a nespereira (Eriobotrya japónica), a laranjeira (Citrus sinensis) e o marmeleiro (Cydonia oblonga). Existem também algumas galerias ripícolas, onde estão presentes 0 amieiro (Alnus glutinosa), o choupo (Populus nigra), o salgueiro (Salix atrocinerea), o freixo (Fraxinus angustifolia) e silvas (Rubus ulmifolius). Observam-se ainda áreas de pasto com aira-cariofilia (Aira caryophyllea) e bole-bole (Briza spp) (Correia & Nisa 1999). A água disponível varia sazonalmente, no Verão a maioria dos cursos de água está seca, enquanto que no Inverno, é possível encontrar água em todos. Apesar da densidade populacional ser reduzida e dispersa em poucos núcleos urbanos, sendo o principal Santa Margarida da Serra, ocorrem várias actividades humanas causadoras de impacto como extracção de cortiça, pastorícia e caça.. 14.
(15) Figura 1 – Distribuição do montado de sobro em Portugal (sombreado) e localização da área de estudo (ponto) (Adaptado de dados de 1995 do Instituto Florestal). A fauna local de pequenos mamíferos é constituída pelo musaranho-dedentes-brancos - Crocidura russula (Hermann, 1780), o musaranho-anão-dedentes-brancos - Suncus etruscus (Savi, 1822), a rata-de-água - Arvicola sapidus (Miller, 1908), a toupeira - Talpa occidentalis (Cabrera 1907), o rato de Cabrera - Microtus cabrerae (Thomas, 1906), o rato-cego-mediterrânico Microtus duodecimcostatus (Sélys-Longchamps, 1839), o rato-do-campo Apodemus sylvaticus (Linnaeus, 1758), o rato-preto - Rattus rattus (Linnaeus,. 15.
(16) 1758), a ratazana - Rattus norvegicus (Berkenhout, 1796) e o rato-das-hortas Mus spretus (Lataste, 1883) (Cabral et al. 2005). Entre estes, o único com estatuto de ameaça em Portugal é o rato de Cabrera. (Cabral et al. 2005). As espécies mais abundantes na área de estudo são o rato-do-campo, o rato-das-hortas e o musaranho-de-dentes-brancos (Mathias & Ramalhinho 1999). Quanto aos carnívoros, verifica-se a ocorrência de raposa - Vulpes vulpes (Linnaeus, 1758), doninha - Mustela nivalis (Linnaeus, 1766), toirão Mustela putorius (Linnaeus, 1758), fuinha - Martes foina (Erxleben, 1777), texugo - Meles meles (Linnaeus, 1758), lontra - Lutra lutra (Linnaeus, 1758), geneta - Genetta genetta (Linnaeus, 1758) e sacarrabos - Herpestes ichneumon (Linnaeus, 1758) (Cabral et al. 2005). As espécies mais abundantes na área de estudo são a doninha, a fuinha, a geneta e o sacarrabos (Santos-Reis et al. 1999).. 16.
(17) METODOLOGIA. Para a realização deste estudo optou-se por uma área de estudo tão extensiva quanto possível tendo para tal sido seleccionadas as cartas militares 506 e 517. A selecção dos locais de amostragem foi feita com recurso a fotografias aéreas e ao software ArcView 3.1 GIS (ESRI 1996) e respectivas extensões, e posterior verificação no local. Os critérios tidos em conta para a selecção das manchas de habitat a amostrar foram os seguintes: tipo de habitat (montado, galeria ripícola e terreno cultivado), área, distância a outras manchas e reduzida influência de auto-correlação espacial.. Pequenos mamíferos. Os métodos directos, além de invasivos, são também mais laboriosos, exigindo mais tempo e recursos. No entanto, a armadilhagem é o método mais fiável na inventariação de pequenos mamíferos, pois são difíceis de observar devido ao pequeno tamanho e hábitos nocturnos e secretivos (Jones et al. 1996, Corominas 2004). Para amostrar a diversidade e abundância de pequenos mamíferos, entre Janeiro e Julho de 2007, foram realizadas sessões de captura, com permissão do Instituto da Conservação da Natureza e da Biodiversidade (ICNB), através de armadilhas de caixa (modelo Sherman) de dois tamanhos. Estas continham algodão e isco (mistura de sardinha em lata e flocos de aveia). Foi realizada uma sessão de amostragem (7 noites consecutivas) em cada local, sendo as armadilhas verificadas todas as manhãs e os animais capturados anestesiados, identificados à espécie, medidos, pesados, verificado o sexo, marcados por corte do pêlo e libertados no respectivo local de captura. Para possibilitar a comparação dos resultados, utilizou-se o mesmo número (50) e disposição de armadilhas (dispostas em cruz a partir do centro do fragmento e distando em média 10m entre si) em cada local de amostragem, independentemente da sua dimensão do fragmento (Pardini et al. 2005). 17.
(18) Foram armadilhados 25 locais (15 áreas cultivadas, 6 galerias ripícolas e 4 zonas de montado).. Carnívoros. Para prospectar os carnívoros utilizou-se o método dos transectos, pois a troca de informação entre os indivíduos é um aspecto muito importante da sua estratégia social, e é realizada através de várias formas: visual, auditiva e olfactiva, sendo a última a mais frequente (Macdonald 2005, Espírito-Santo et al. 2007). Este método permite analisar áreas relativamente grandes (Virgós 2001, Virgós et al. 2002) e em tipos de habitats variados (Sadlier et al. 2004) e está comprovada a boa relação entre os indícios detectados e a densidade populacional (Cresswell et al. 1989, Cavallini 1994, Staender 1998, Virgós et al. 2002, Wemmer et al. 1996). Uma vez que o método utilizado é menos laborioso, foi possível procurar indícios de presença de carnívoros nos mesmos locais onde se realizou a armadilhagem de pequenos mamíferos e ainda aumentar número de manchas amostradas com este método. Entre Janeiro e Julho de 2007, foram prospectados os indícios de presença de carnívoros ao longo de transectos de 1,5km de extensão, tendo por base a rede existente de caminhos de terra batida (Virgós et al. 2002, Paniagua et al. 2004), padronizando assim a probabilidade de detecção dos indícios nos diferentes habitats. De forma a minimizar os erros de identificação, utilizaram-se várias características distintivas na identificação dos indícios (morfologia, comprimento, largura, diâmetro, odor), e foram excluídos os de identificação dúbia (Kendall et al. 1992, Sadlier et al. 2004). Para avaliação da diversidade e abundância de carnívoros realizaram-se 47 transectos (25 em áreas cultivadas, 14 em galerias ripícolas e 8 em zonas de montado).. 18.
(19) Análise de dados. Calculou-se a riqueza específica e a diversidade (Índice de ShannonWiener – Krebs 1989) de mamíferos para os 25 locais onde se prospectaram pequenos mamíferos e carnívoros. Calculou-se a abundância de pequenos mamíferos e de carnívoros em cada local amostrado e a média e desvio padrão do mesmo índice por tipo de habitat (montado, galeria ripícola e cultura). Para verificar a ocorrência de diferenças na abundância média entre as três distintas categorias de manchas recorreu-se à ANOVA não paramétrica (Zar 1996). Para averiguar a associação entre a comunidade de pequenos mamíferos e de carnívoros e as manchas de habitat amostradas, efectuou-se uma análise de correspondências. Tendo as características das manchas de habitat (área, perímetro, distância à horta/olival mais próximo, mediana das distâncias às hortas/olivais mais próximos e distância à galeria ripícola mais próxima) como variáveis independentes, e os índices de diversidade e abundância, como variáveis dependentes, um modelo de regressão múltipla permitiu averiguar a contribuição de cada um dos descritores considerados na respectiva diversidade e abundância de pequenos mamíferos ou carnívoros.. Todas as análises estatísticas foram realizadas com recurso ao software STATISTICA 7 (StatSoft).. 19.
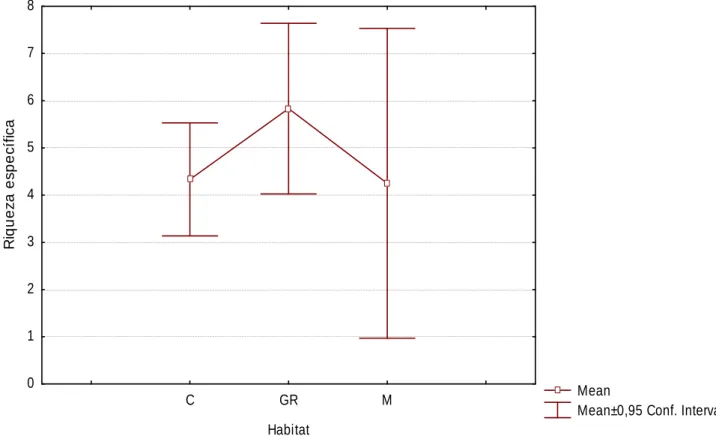
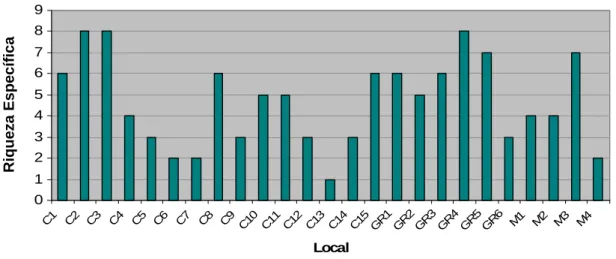
(20) RESULTADOS. Riqueza Específica e Diversidade de Mamíferos. Não se verificam diferenças significativas na riqueza específica entre os 3 habitats considerados (Kruskal-Wallis H. (2, N=25). = 2,485; p= 0,289); no. entanto, observa-se que as galerias ripícolas apresentam riqueza específica superior aos restantes habitats (Figura 1).. 8 7. Riqueza específica. 6 5 4 3 2 1 0 C. GR. M. Mean Mean±0,95 Conf. Interval. Habitat. Figura 1 – Média e desvio padrão da riqueza específica por habitat (C-Cultura, GR-Galeria Ripícola, M-Montado).. A nível local, a riqueza específica evidencia uma grande variabilidade entre os locais de amostragem nas culturas e no montado, verificando-se locais com riqueza específica de mamíferos muito elevada e muito reduzida (Figura 2); nas galerias ripícolas os valores de riqueza são relativamente mais homogéneos entre os vários sítios amostrados. 20.
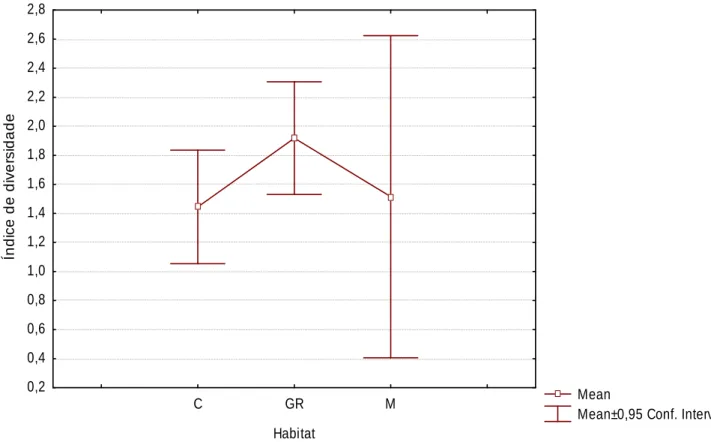
(21) Os locais que apresentam riqueza específica mais elevada (C2 e C3) são culturas abandonadas, com coberto vegetal denso, situadas junto a cursos de. água,. características. apelativas. para. vários. mamíferos,. pelas. potencialidades a nível de refúgio e de fontes de alimento. Quanto à zona ripícola (GR4), trata-se de uma ribeira permanente, com galeria relativamente desenvolvida, pelo que, constitui também habitat adequado para várias espécies. Quanto à amostra com menor riqueza específica (C13), trata-se de uma horta de pequena dimensão, bem cuidada e com coberto vegetal praticamente inexistente, além de se situar junto a uma casa e a uma estrada com alguma utilização.. R. Específic Riqueza Específica. 9 8 7 6 5 4 3 2 1 C 1 C 2 C 3 C 4 C 5 C 6 C 7 C 8 C 9 C 10 C 11 C 12 C 13 C 14 C 15 G R 1 G R 2 G R 3 G R 4 G R 5 G R 6 M 1 M 2 M 3 M 4. 0. Local. Figura 2 – Riqueza específica de mamíferos nos locais amostrados (C-Cultura, GR-Galeria Ripícola, M-Montado).. Tal como é explícito na figura 3, a diversidade de mamíferos nos locais prospectados não é significativamente diferente nas zonas cultivadas, galerias ripícolas e montado (Kruskal-Wallis H. (2, N=25). = 3,031; p= 0,2197). Ainda assim,. o índice de diversidade é semelhante nas áreas cultivadas e no montado, apresentando um valor mais elevado nas galerias ripícolas.. 21.
(22) 2,8 2,6 2,4. Índice de diversidade. 2,2 2,0 1,8 1,6 1,4 1,2 1,0 0,8 0,6 0,4 0,2 C. GR. M. Mean Mean±0,95 Conf. Interval. Habitat. Figura 3 – Média e desvio padrão do índice de diversidade (índice de ShannonWiener) por habitat (C-Cultura, GR-Galeria Ripícola, M-Montado).. A figura 4 ilustra uma grande variabilidade do índice de diversidade dentro de cada tipo de habitat, sendo mais evidente nos locais de culturas e de montado. O valor de diversidade mais elevado corresponde a uma área cultivada (C2) o que pode dever-se ao facto de a área em causa ser um olival abandonado, com vegetação densa e com uma galeria ripícola associada, pelo que, apresenta condições adequadas a uma grande variedade de espécies. Quanto aos locais com menor índice de diversidade (C6, C7 e C13), o primeiro situa-se junto à casa da Herdade da Ribeira Abaixo, pelo que é uma zona com elevado grau de perturbação humana; o segundo é uma zona de pastoreio intensivo de vacas, o que diminui a cobertura vegetal, tornando o local não aprazível, nomeadamente para os pequenos mamíferos; finalmente o C13 é uma horta de pequena dimensão, bem cuidada e com coberto vegetal praticamente inexistente, além de se situar junto a uma casa e a uma estrada.. 22.
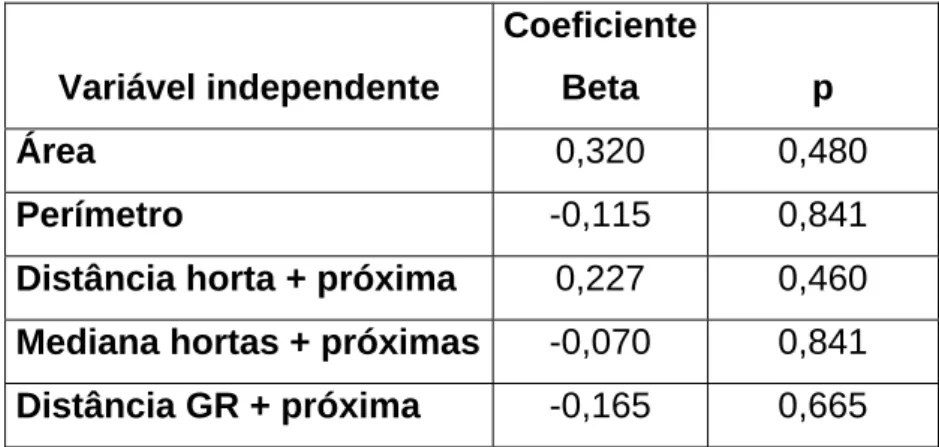
(23) Índice de Diversidade Índice de Diverssida. 3 2,5 2 1,5 1 0,5. C 1 C 2 C 3 C 4 C 5 C 6 C 7 C 8 C 9 C 10 C 11 C 12 C 13 C 14 C 15 G R 1 G R 2 G R 3 G R 4 G R 5 G R 6 M 1 M 2 M 3 M 4. 0. Local. Figura 4 – Índice de diversidade (índice de Shannon-Wiener) de mamíferos nos locais amostrados (C-Cultura, GR-Galeria Ripícola, M-Montado).. Apesar dos resultados não serem significativos (Tabela 1), o factor que mais influencia a diversidade de mamíferos nos fragmentos de habitat nãomatriz é a dimensão dos mesmos, seguida da distância à horta mais próxima; o factor com menos influência é a mediana das distâncias às hortas mais próximas.. Tabela 1 – Efeito da dimensão e distância entre fragmentos não-matriz na diversidade de mamíferos (análise de regressão múltipla).. Coeficiente Variável independente. Beta. p. Área. 0,320. 0,480. Perímetro. -0,115. 0,841. Distância horta + próxima. 0,227. 0,460. Mediana hortas + próximas. -0,070. 0,841. Distância GR + próxima. -0,165. 0,665. 23.
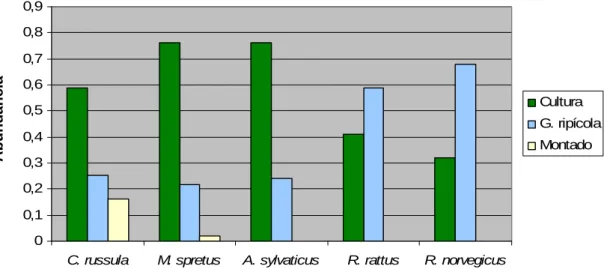
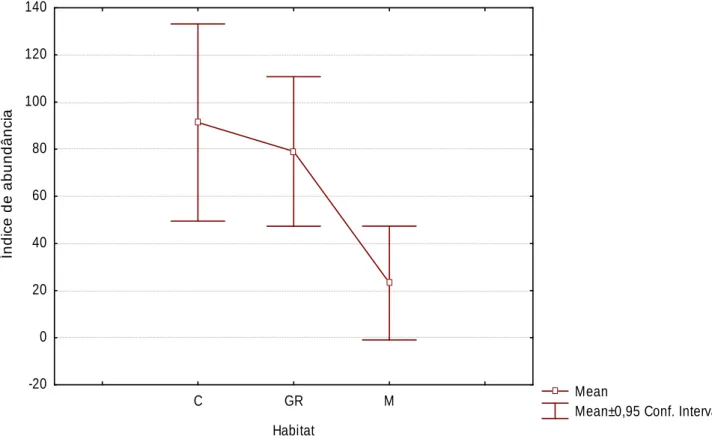
(24) Pequenos mamíferos. Capturaram-se 417 pequenos mamíferos de 5 espécies: musaranho-dedentes-brancos, rato-do-campo, rato-preto, ratazana e rato-das-hortas, sendo o último o mais abundante (61,4%) e rato-preto o menos abundante (0,7%). De uma forma geral, a figura 5 mostra uma abundância de pequenos mamíferos muito reduzida na matriz, para todas as espécies, sendo musaranho-de-dentes-brancos e rato-das-hortas as únicas espécies presentes neste habitat. É evidente a diferença entre as espécies do género Rattus e as restantes; enquanto que as primeiras são mais abundantes nas galerias ripícolas, as últimas estão mais presentes nas hortas/olivais.. 0,9 0,8. Abundância Abundânci. 0,7 0,6 Cultura. 0,5. G. ripícola 0,4. Montado. 0,3 0,2 0,1 0 C. russula. M. spretus. A. sylvaticus. R. rattus. R. norvegicus. Figura 5 – Abundância de pequenos mamíferos por espécie e por habitat. Não. se. verificam. diferenças. estatisticamente. significativas. na. abundância de pequenos mamíferos entre os 3 tipos de habitat (Kruskal-Wallis H. (2, N=25). = 3,848; p= 0,146). No entanto, em geral, o gráfico mostra. abundâncias bastante mais reduzidas nos locais de montado e as abundâncias mais. elevadas. registadas. pertencem. a. áreas. cultivadas. (Figura. 6).. 24.
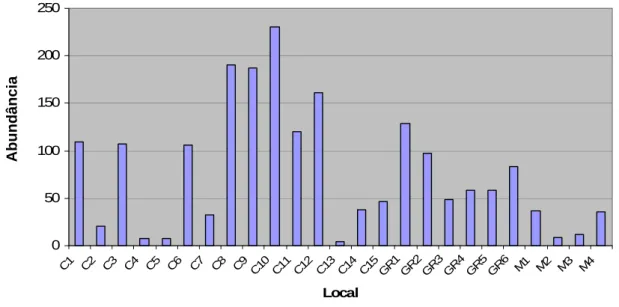
(25) 140 120. Índice de abundância. 100 80 60 40 20 0 -20 C. GR. M. Mean Mean±0,95 Conf. Interval. Habitat. Figura 6 – Média e desvio padrão dos índices de abundância de pequenos mamíferos em cada habitat (C-Cultura, GR-Galeria Ripícola, M-Montado).. À semelhança do observado para o índice de diversidade, também a abundância de pequenos mamíferos variou de forma notória nos vários locais amostrados para cada tipo de habitat, especialmente nas áreas cultivadas, que incluem os locais respectivamente com maior e menor abundância de pequenos mamíferos (Figura 7).. 25.
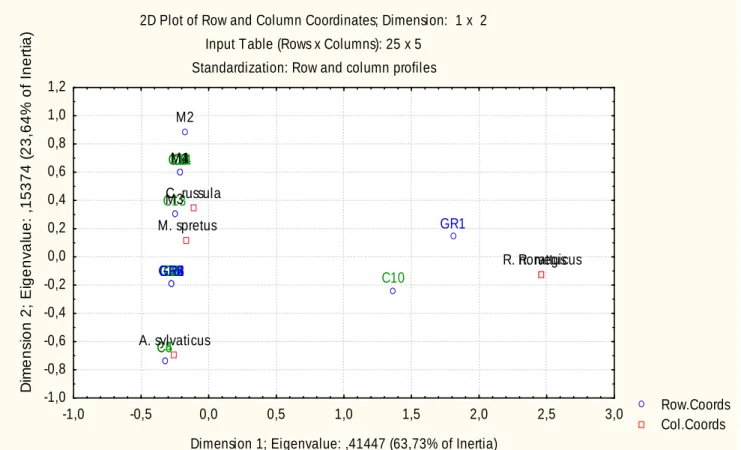
(26) 250. Abundância Abundânci. 200. 150. 100. 50. C 1 C 2 C 3 C 4 C 5 C 6 C 7 C 8 C 9 C 10 C 11 C 12 C 13 C 14 C 15 G R 1 G R 2 G R 3 G R 4 G R 5 G R 6 M 1 M 2 M 3 M 4. 0. Local. Figura 7 – Índice de abundância de pequenos mamíferos por local de amostragem (C-Cultura, GR-Galeria Ripícola, M-Montado).. A análise de correspondência gerou 2 eixos factoriais que, em conjunto, explicam cerca de 87% da variância total da abundância de pequenos mamíferos. Observa-se uma clara associação entre rato-preto e ratazana, sendo ambos fortemente influenciados pelo eixo 1. As restantes espécies estão mais correlacionadas com o eixo 2; verifica-se ainda uma associação entre musaranho-de-dentes-brancos e rato-das-hortas. As espécies do género Rattus encontram-se associadas a galerias ripícolas e zonas urbanizadas. O rato-do-campo está associado a áreas cultivadas e o rato-das-hortas e o musaranho-de-dentes-brancos estão predominantemente ligados a dois tipos de habitat, culturas e montado.. 26.
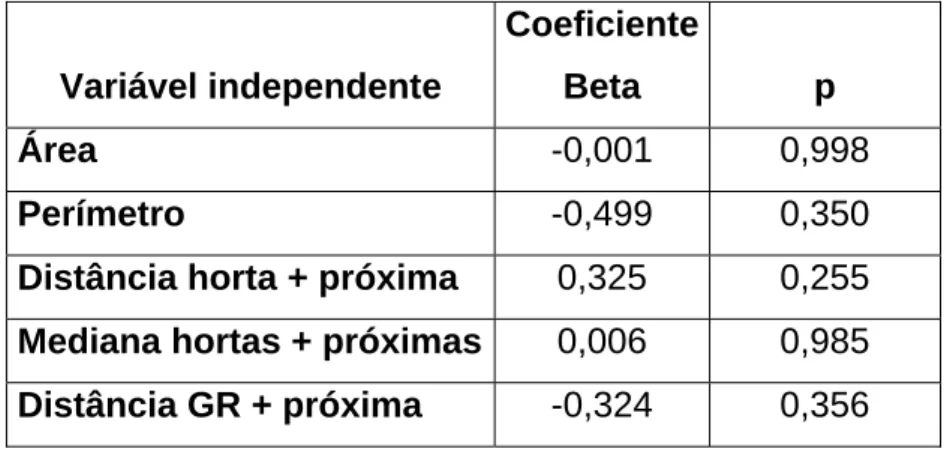
(27) Dimension 2; Eigenvalue: ,15374 (23,64% of Inertia). 2D Plot of Row and Column Coordinates; Dimension: 1 x 2 Input Table (Rows x Columns): 25 x 5 Standardization: Row and column profiles 1,2 1,0. M2. 0,8 M1 M4 C14 C11 C2 C7 C6. 0,6 0,4. C. russula M3 C13. 0,2. M. spretus. GR1. 0,0. R. R. norvegicus rattus. GR6 GR5 GR4 GR3 GR2 C12 C15 C1 C3 C9 C8. -0,2. C10. -0,4 A. sylvaticus C5 C4. -0,6 -0,8 -1,0 -1,0. -0,5. 0,0. 0,5. 1,0. 1,5. 2,0. 2,5. 3,0. Dimension 1; Eigenvalue: ,41447 (63,73% of Inertia). Figura 8 – Espaço factorial (eixos 1 e 2), incluindo espécies de pequenos mamíferos e locais de amostragem (C-Cultura, GR-Galeria Ripícola, MMontado).. A. abundância. de. pequenos. mamíferos. não. é. influenciada. significativamente por nenhum dos factores em análise (Tabela 2). No entanto, os dados indicam que as características da mancha que mais afectam a abundância são o perímetro e a distância à galeria ripícola mais próxima, que estão relacionadas negativamente, e a distância à horta mais próxima, que tem uma relação positiva. A variável que menos afecta a abundância de pequenos mamíferos é a área das manchas de habitat não-matriz.. 27. Row.Coords Col.Coords.
(28) Tabela 2 – Efeito da dimensão e distância entre fragmentos não-matriz na abundância de pequenos mamíferos (análise de regressão múltipla).. Coeficiente Variável independente. Beta. p. Área. -0,001. 0,998. Perímetro. -0,499. 0,350. Distância horta + próxima. 0,325. 0,255. Mediana hortas + próximas. 0,006. 0,985. Distância GR + próxima. -0,324. 0,356. Carnívoros. Foram recolhidos 182 indícios de presença de sete espécies de carnívoros: sacarrabos, fuinha, lontra, geneta, raposa, texugo e doninha; a espécie mais detectada foi a fuinha (28,0%) e a com menor número de registos foi a doninha (2,2%). A abundância de carnívoros, não difere significativamente consoante o tipo de habitat (Kruskal-Wallis H. (2, N=25). = 2, 326; p= 0,3125); no entanto o. montado apresenta a média mais reduzida, enquanto que o habitat com abundância média de carnívoros mais elevada é a galeria ripícola (Figura 9).. 28.
(29) 6. Índice de diversidade. 5. 4. 3. 2. 1. 0 C. GR. M. Mean Mean±0,95 Conf. Interval. Habitat. Figura 9 – Média e desvio padrão da abundância de carnívoros nos 3 habitats (C-Cultura, GR-Galeria Ripícola, M-Montado). Observa-se uma elevada variabilidade dos níveis de abundância encontrados individualmente em cada habitat (Figura 10), sendo que os três valores mais elevados de abundância representam os três habitats em análise. O local com maior abundância (GR1) corresponde a uma das maiores ribeiras permanentes da área de estudo, com galeria ripícola muito desenvolvida e o valor deve-se principalmente à presença de lontra. A cultura com abundância mais elevada (C3) é um olival abandonado junto a uma galeria pouco desenvolvida e o maior contributo é da geneta, confirmando a sua preferência por olivais e vegetação ripícola (Calzada 2002), provavelmente existe uma latrina nas imediações. O principal responsável pela abundância encontrada no montado (M4) é a raposa, o que é justificado pelos seus hábitos generalistas, podendo sobreviver da exploração de variadas fontes de alimento e pelo facto das áreas de montado não serem muito extensas, pelo que, podese inferir que as áreas vitais da raposa incluem habitat matriz e não-matriz.. 29.
(30) O facto de ocorrerem vários locais de amostragem com resultados nulos pode dever-se ao facto de serem áreas cultivadas situadas nas proximidades de habitações e estradas.. Abundância Abundânc. 12 10 8 6 4 2. M 8. M 6. M 4. C 9 C 11 C 13 C 15 C 17 C 19 C 21 C 23 G R 2 G R 4 G R 6 G R 8 G R 10 G R 12 G R 14 M 2. C 7. C 5. C 3. C 1. 0. Local Local. Figura 10 – Índice de abundância de carnívoros em cada local amostrado (CCultura, GR-Galeria Ripícola, M-Montado).. Os dois primeiros eixos factoriais da análise de correspondências relativa à abundância de carnívoros justificam aproximadamente 50% da variabilidade. As espécies mais correlacionadas com o eixo 1 são o sacarrabos e a fuinha, enquanto que, a raposa, o texugo e a geneta se encontram associadas ao eixo 2. A lontra está fortemente associada às galerias ripícolas amostradas. Verifica-se um agrupamento das restantes espécies de carnívoros, à excepção da doninha.. 30.
(31) Dimension 2; Eigenvalue: ,35879 (19,10% of Inertia). 2D Plot of Row and Column Coordinates; Dimension: 1 x 2 Input Table (Rows x Columns): 36 x 7 Standardization: Row and column profiles 3,0 2,5. M. nivalis. 2,0. C8. 1,5 GR7. 1,0 0,5 0,0 -0,5. L. lutra. -1,0. GR2 GR1 GR4. C18M2 H. ichneumon GR8 C13 C17 M7 C9 C7 M. C10 foina V. vulpes C1GR9 C3C4 M4 GR11 GR5 C5 M1C21 M3 C22 C23 C6M. meles M6 GR10 GR3 GR12 GR13 G. genetta C2 C16 M8. -1,5 -2,0 -3,0. -2,5. -2,0. -1,5. -1,0. -0,5. 0,0. 0,5. 1,0. 1,5. Dimension 1; Eigenvalue: ,57398 (30,56% of Inertia). Figura 11 – Diagrama da análise de correspondência dos eixos 1 e 2, incluindo espécies de carnívoros e locais de amostragem (C-Cultura, GR-Galeria Ripícola, M-Montado).. Pela análise da tabela 3 verifica-se que a mediana da distância às hortas mais próximas é a característica que mais afecta negativamente a abundância de carnívoros, seguida da distância à galeria mais próxima, apesar de nenhuma revelar valores significativos. O factor com menos influência na abundância de carnívoros é a dimensão de fragmentos de habitat não-matriz.. 31. Row.Coords Col.Coords.
(32) Tabela 3 – Efeito da dimensão e distância entre fragmentos não-matriz na abundância de carnívoros (análise de regressão múltipla).. Coeficiente Variável independente. Beta. p. Área. 0,027. 0,928. Perímetro. 0,080. 0,816. Distância horta + próxima. 0,151. 0,515. Mediana hortas + próximas. -0,313. 0,202. Distância GR + próxima. -0,189. 0,475. 32.
(33) DISCUSSÃO DOS RESULTADOS. Riqueza Específica e Diversidade de Mamíferos. Os resultados relativos tanto à riqueza como à diversidade de mamíferos revelam valores semelhantes para culturas e montado, o que pode dever-se ao facto das manchas de culturas apresentarem, em geral, pequenas dimensões e se encontrarem dispersas na matriz de montado, a riqueza específica correspondente às galerias ripícolas é ligeiramente superior, tal como era esperado, visto que os mamíferos utilizam frequentemente as zonas ripícolas como fonte de água, alimento e refúgio (Martin 2002). Estes resultados são concordantes com as conclusões obtidas noutros trabalhos realizados na mesma área (Santos-Reis et al. 2004, Pinto 1998, Pinheiro 2006). O. facto. de. não. serem. detectáveis. diferenças. estatisticamente. significativas nos três habitats em análise deve-se ao facto dos mamíferos utilizarem mais frequentemente as galerias ripícolas mas não estarem exclusivamente restritos a este habitat, utilizando também os restantes (Martin 2002). O factor que mais influencia a diversidade de mamíferos nos fragmentos de habitat não-matriz é a dimensão dos mesmos, verificando-se uma proporcionalidade positiva entre área da mancha e diversidade de mamíferos, tal como esperado (Virgós et al. 2002). A importância da distância à horta mais próxima justifica-se, pois numa menor área há uma maior concentração de diferentes fontes de alimento, que patrocina a sobrevivência de uma maior diversidade de espécies.. Pequenos mamíferos. As espécies mais abundantes no montado (musaranho-de-dentesbrancos e rato-das-hortas) são as expectáveis uma vez que estão tradicionalmente associadas a zonas de vegetação mediterrânica (Blanco 1998). O musaranho-de-dentes-brancos é frequente em áreas cultivadas e 33.
(34) aproxima-se de núcleos urbanos (Blanco 1998, López-Fuster 2002); já o ratodas-hortas utiliza espaços abertos, com vegetação herbácea, condições encontradas nas zonas de cultivo (Palomo 2002). O rato-do-campo é uma das espécies de pequenos mamíferos mais consumida por predadores, pelo que, prefere. zonas. com. boa. cobertura. arbustiva,. onde. encontra. maior. disponibilidade de refúgio (Jubete 2002). É evidente a diferença entre as espécies do género Rattus e as restantes; enquanto que as primeiras são mais abundantes nas galerias ripícolas, as últimas estão mais presentes nas hortas/olivais. O rato-preto ocupa habitats muito diversos, inclusive pequenos núcleos urbanos desde que tenha cobertura vegetal suficiente (Blanco 1998, Zamorano & Palomo 2002). Já a ratazana é fortemente comensal, habitando meios urbanos e rurais e está também muito ligada à presença de água (Blanco 1998, Rojas & Palomo 2002). A. média. da. abundância. de. pequenos. mamíferos. não. difere. significativamente entre os três habitats pois, quando em reduzida densidade, a distribuição dos indivíduos é aleatória e, portanto, não é afectada pela qualidade do habitat (Corominas 2004). A reduzida abundância de pequenos mamíferos no montado pode deverse ao facto da maior parte destas áreas se encontrar praticamente desprovida de subcoberto, devido ao pastoreio e de forma a facilitar a extracção da cortiça, não proporcionando assim o refúgio essencial para os pequenos mamíferos. Os locais com maior abundância de pequenos mamíferos são olivais situados perto de zonas urbanas, o que é justificado pelo carácter generalista e oportunista destas espécies (Blanco 1998), principalmente o rato-preto e a a ratazana. Outro factor que contribui para as preferências por parte destes animais é a existência de coberto vegetal denso (que se verifica frequentemente em olivais abandonados e galerias ripícolas) pois promove a alimentação e o refúgio (Gonnet & Ojeda 1998, Corominas 2004, Converse et al. 2006). O contexto multivariado mostra uma clara associação entre rato-preto e ratazana e musaranho-de-dentes-brancos e rato-das-hortas. As espécies do género Rattus encontram-se associadas a galerias ripícolas e zonas urbanizadas, que são os habitats geralmente mais utilizados por estes mamíferos (Blanco 1998, Rojas & Palomo 2002, Zamorano & Palomo 2002). 34.
(35) Neste caso, as galerias ripícolas estão associadas a ribeiras permanentes e as culturas são olivais de grandes dimensões. O rato-do-campo está associado a olivais abandonados, o que é explicado, além da disponibilidade de alimento, pela densidade de coberto vegetal que proporciona melhor refúgio contra os predadores, necessidade que é maior nesta espécie do que nos restantes insectívoros devido às suas dimensões superiores. Quanto ao rato-das-hortas e ao musaranho-de-dentes-brancos, estão predominantemente ligados a dois tipos de habitat, cultura e montado; o primeiro está associado a ambientes tipicamente mediterrânicos e espaços abertos, como o montado, e, adicionalmente, tem escassos requerimentos hídricos (Blanco 1998, Palomo 2002); enquanto que o segundo prefere habitats abertos e campos de cultivo e frequenta as proximidades de núcleos urbanos (Blanco 1998, López-Fuster 2002). Os dados indicam que as características dos fragmentos de habitat nãomatriz que mais afectam a abundância de pequenos mamíferos são o perímetro e a distância à galeria ripícola mais próxima, ambas relacionadas negativamente com a variável resposta. Assim, a abundância de pequenos mamíferos é menor em fragmentos com maior perímetro, o que se deve ao efeito de margem, pois está documentado que os predadores terrestres caçam com maior eficácia ao longo das orlas dos habitats (Lidicker 1999); estudos com roedores demonstram que estas espécies utilizam menos as margens do terreno do que o seu interior, uma vez que na margem se encontram mais vulneráveis aos predadores (Bowers & Dooley 1999). Tal como esperado, a abundância de pequenos mamíferos é tanto mais elevada quanto menor for a distância a galerias ripícolas. A distância à horta mais próxima tem uma relação positiva pois influencia directamente a abundância de pequenos mamíferos, o que está relacionado com o facto de estes animais apenas terem capacidade para percorrer distâncias pequenas, a dispersão média daria não excede os 100m (Blanco 1998), e assim conseguirem uma maior variedade de fontes de alimento a uma curta distância. A dimensão do fragmento constitui o factor menos importante para a ocorrência de pequenos mamíferos; tal como verificado em trabalhos 35.
(36) anteriores, a densidade de pequenos mamíferos não é mais elevada em fragmentos de dimensão superior (Arvicola ssp., Microtus ssp., Foster & Gaines 1991). Está também demonstrada a capacidade dos pequenos mamíferos de viver em manchas pequenas, quando os habitats são heterogéneos (Andrén 1994, Burel et al. 1998).. Carnívoros. Visto. que. os. carnívoros. ocupam. vastos. territórios,. excedendo. 2. geralmente 3km (Elmeros et al. 2005, Rountree 2004) era já esperado que utilizassem regularmente diversos fragmentos dispersos na matriz (Paniagua et al. 2004; Virgós et al. 2002), não se verificando diferenças significativas na abundância média de carnívoros nos três habitats em estudo. Ainda assim, o habitat com abundância média mais alta é a galeria ripícola, pois os carnívoros utilizam frequentemente este habitat como refúgio e/ou fonte de alimento, tal como detectado em estudos anteriores (Naiman et al. 1993, Virgós 2001, Martin 2002, Macdonald 2005), provavelmente devido à contribuição para amenizar o clima localmente (Virgós 2001); adicionalmente, e em particular na área de estudo, está documentado que os carnívoros predam lagostim americano, Procambarus clarkii, que é muito comum nos cursos de água da região (Santos et al. 2007). As espécies generalistas (sacarrabos, fuinha, geneta, raposa e texugo) estão associadas a culturas e montado, onde estas estão inseridas, proporcionando uma enorme variedade de fontes de alimento; pelo contrário, as espécies especialistas estão claramente afastadas deste habitat mais humanizado. Tal como esperado, verifica-se uma forte associação entre a lontra e as galerias ripícolas; assim como, entre as restantes espécies (à excepção da doninha) e as manchas cultivadas ou de montado. A doninha está associada a uma ribeira (GR7) e a uma cultura numa zona urbanizada (C8), locais onde ocorrem frequentemente pequenos mamíferos, principal fonte de alimento deste carnívoro (Gisbert & Santos-Reis 2002, Paniagua et al. 2004).. 36.
(37) A mediana da distância às hortas mais próximas é o descritor que mais afecta a abundância de carnívoros, seguido da distância à galeria mais próxima, ambos de forma negativa. Estes resultados são contrários ao que era esperado face às extensas áreas vitais que caracterizam este grupo de mamíferos; seria de esperar que a abundância aumentasse quando a distância a outras hortas é reduzida, ou seja quando o seu território inclui várias hortas próximas; mais ainda, como já foi referido, era esperado que a abundância estivesse também relacionada com as galerias ripícolas, o que não se verifica. Talvez estes resultados se devam ao facto do método utilizado permitir apenas distinguir espécies e não indivíduos, o que pode fazer com que a abundância detectada num determinado local seja na verdade superior à real, uma vez que o mesmo indivíduo pode ser responsável por vários indícios. O factor que menos importância tem para a abundância de carnívoros é a dimensão da mancha de habitat não-matriz o que é coerente uma vez que um animal da dimensão destas espécies não está restrito a uma única mancha, incluído várias no seu território. Finalmente, os resultados obtidos permitem corroborar a hipótese em análise, uma vez que se confirma o papel preponderante das manchas de habitat não-matriz na biodiversidade de mamíferos encontrada na paisagem típica do montado.. 37.
(38) CONSIDERAÇÕES FINAIS. O presente trabalho padece de algumas limitações, sendo a principal o reduzido intervalo de tempo disponível para a realização da amostragem, razão pela qual os dados dos diferentes locais, apesar de recolhidos num intervalo de tempo heterogéneo, terem sido tratados e comparados como se reportassem à mesma época, o que implica erros pois a utilização do espaço por parte dos animais. difere,. nomeadamente,. em. época. seca/húmida,. época. de. reprodução/não reprodução. Seria mais exacto proceder às amostragens na mesma época em anos consecutivos. Outra das limitações evidentes é a não detecção de algumas espécies de mamíferos dadas como presentes na área de estudo, nomeadamente o musaranho-anão-de-dentes-brancos, a rata-de-água, o rato de Cabrera, o ratocego-mediterrânico e o toirão; mais uma vez devido aos constrangimentos de tempo, optou-se por utilizar apenas métodos não selectivos de modo a detectar o maior número de espécies possível, no entanto, estes métodos têm alguns problemas conhecidos. O método dos transectos apresenta vários inconvenientes que podem mascarar os resultados, tais como, a imprecisão que confere ao cálculo de abundâncias, a dependência das condições atmosféricas e do substrato, a influência da dieta e do comportamento social na produção de dejectos (Paniagua et al. 2004, Sadlier et al. 2004), além do que, o padrão encontrado numa única visita por fragmento pode não reflectir a utilização regular do mesmo (Virgós 2001). Entre. as. limitações. da. armadilhagem,. destaca-se. o. possível. enviesamento nos dados obtidos devido ao clima, ao tipo de armadilha e isco utilizado (Corominas 2004). Uma vez que o tamanho da amostra é relativamente reduzido, as análises estatísticas efectuadas devem ser interpretadas com precaução, tendo em conta os possíveis factores causadores de erros, tais como os referidos anteriormente. Apesar de ser reconhecido que os recursos se distribuem no ecossistema de forma heterogénea, o grau de heterogeneidade e a dimensão e 38.
(39) forma como as diferentes manchas (fragmentos) se distribuem no espaço pode influenciar os resultados obtidos. Em particular na área de estudo, as manchas de habitat não-matriz são de tamanho médio reduzido e encontram-se muito dispersas na paisagem, pelo que, era de esperar que os mamíferos, sendo espécies com elevada mobilidade, frequentem vários habitats, mascarando assim as diferenças de abundância e diversidade. Adicionalmente, em locais altamente heterogéneos verifica-se uma elevada dispersão diária por parte dos animais (Jager et al. 2006), neste tipo de ambiente e tratando-se maioritariamente de mamíferos generalistas, os animais adoptam um comportamento oportunista, frequentando os locais com maior vantagem em determinado momento (Law & Dickman 1998). Quanto aos pequenos mamíferos presume-se que, tendo menos capacidade de dispersão do que os carnívoros, em épocas de abundância reduzida, a sua distribuição é menos afectada pelo habitat, podendo mesmo ser aleatória (Corominas 2004). Em geral, pode-se concluir que o montado de sobro por si só não garante a conservação de mamíferos em ecossistemas mediterrânicos, são os fragmentos de habitat não-matriz que asseguram a abundância e diversidade das espécies. Constatou-se que as galerias ripícolas dão um importante contributo para a riqueza específica e diversidade de mamíferos, pelo que, é fundamental contemplar este tipo de habitat nos planos de conservação em regiões mediterrânicas. Em particular, a abundância de pequenos mamíferos está muito dependente das áreas cultivadas, assim, a preservação destes fragmentos é essencial para assegurar ecossistemas equilibrados e saudáveis, não sujeitos às consequências duma diminuição da abundância de presas. No sentido de conservar a diversidade de mamíferos típica do montado é ainda necessário considerar a dimensão e distância entre as manchas de habitat não-matriz, uma vez que é fundamental maximizar a área (em especial das manchas cultivadas) e minimizar a distância aos restantes fragmentos. Assim, é necessário um plano de gestão da paisagem típica do montado que tenha em conta a manutenção de um grau moderado de humanização da paisagem, de forma a favorecer uma comunidade diversificada de mamíferos.. 39.
(40) BIBLIOGRAFIA. Andrén, H. (1994). Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: A Review. Oikos 71: 355-366.. Antongiovanni, M. & J. P. Metzger (2004). Influence of matrix habitats on the occurrence of insectivorous bird species in Amazonian forest fragments. Biological Conservation 122: 441–451.. August, P. V. (1983). The role of habitat complexity and heterogeneity in structuring tropical mammal communities." Ecology 64: 1495-1507.. Bazzaz, F. A. (1975). Plant species diversity in old-field successional ecosystems in southern Illinois. Ecology 56: 485-488.. Bell, W. J. (1990). Searching behavior patterns in insects. Annual Review of Entomology 35: 447–467.. Bell, W. J. (1991). Searching behaviour: the behavioural ecology of finding resources. London, Chapman and Hall.. Blanco, J. C. (1998). Mamíferos de España. Barcelona, Planeta.. Blondel, J. and J. Aronson (1999). Biology and wildlife of the Mediterranean Region. London, Oxford University Press.. Borralho, R., F. Rego, et al. (1995). The distribution of the egyptian mongoose Herpestes ichneumon (L.) in Portugal. Mammal Review 25: 229-236.. Bove, E. D. & R. Isotti (2001). The european badger (Meles meles) diet in a mediterranean area." Hystrix Italian Journal of Mammalogy 12: 19-25. 40.
(41) Bowers, M. A. & J. L. Dooley (1999). "A controlled, hierarchical study of habitat fragmentation: responses at the individual, patch, and landscape scale." Landscape Ecology 14: 381-389.. Bright, P. W. (1993). Habitat fragmentation: problems and predictions for British mammals. Mammal Review 23: 101-111.. Burel, F., J. Baudry, et al. (1998). Comparative biodiversity along a gradient of agricultural landscapes. Acta Oecologica 19 (47-60).. Cabral MJ (coord.), Almeida J, Almeida PR, Dellinjer T, Ferrand de Almeida N, Oliveira ME, Palmeirim JM, Queiroz AI, Rogado L & Santos-Reis M (eds) (2005). Livro Vermelho dos Vertebrados de Portugal. Instituto da Conservação da Natureza. Lisboa. 660 pp.. Calzada, J. (2002). Genetta genetta. Atlas de los Mamíferos Terrestres de España. J. L. Palomo and J. Gisbert. Madrid, Dirección General de Conservación de la Naturaleza-SECEM-SECEMU.. Cartar, R. V. & L. A. Real (1997). Habitat structure and animal movement: the behaviour of bumble bees in uniform and random spatial resource distributions. Oecologia 112: 430–434.. Cavallini, P. (1994). Feaces count as an index of fox abundance. Acta Theriologica 39: 417–424.. Christophersen, R. G., R. C. Kuntz, et al. (2005). A survey of forest carnivore species composition and distribution in north cascades national park service complex, Washington. Sedro Woolley, USDI National Park Service.. Converse, S. J., W. M. Block, et al. (2006). Small mammal population and habitat responses to forest thinning and prescribed fire. Forest Ecology and Management 228: 263–273. 41.
(42) Corbalán, V. (2005). Microhabitat selection by murid rodents in the Monte desert of Argentina. Journal of Arid Environments 65: 102-110.. Corbalán, V., S. Tabeni, et al. (2006). Assessment of habitat quality for four small mammal species of the Monte Desert, Argentina. Mammalian Biology 71(227-237).. Corominas, I. T. (2004). Distribution, population dynamics and habitat selection of small mammals in Mediterranean environments: the role of climate, vegetation structure, and predation risk. Departament de Biologia. Barcelona, Universitat Barcelona. PhD thesis.. Correia, A. I. & S. Nisa (1999). Flora Vascular. Caracterização da flora e da fauna do montado da herdade da Ribeira Abaixo (Grândola - Baixo Alentejo). M. Santos-Reis & A. I. Correia. Lisboa, Centro de Biologia Ambiental: 47-60.. Correia, A. I. & M. Santos-Reis (1999). Área de Estudo. Caracterização da flora e da fauna do montado da Herdade da Ribeira Abaixo (Grândola - Baixo Alentejo). M. Santos-Reis & A. I. Correia. Lisboa, Centro de Biologia Ambiental: 5-8.. Cresswell, J. E. (1997). Spatial heterogeneity, pollinator behaviour and pollinator-mediated gene flow: bumblebee movements in variously aggregated rows of oil-seed rape. Oikos 78: 546–556.. Dauber, J., M. Hirsch, et al. (2003). Landscape structure as an indicator of biodiversity: matrix effects on species richness. Agriculture, Ecosystems and Environment 98: 321-329.. Díaz, M. (1992). Rodent seed predation in cereal crop areas of Central Spain: effects of physiognomy food availability and predation risk. Ecography 15: 7785.. 42.
(43) Díaz, M., F. J. Pulido, et al. (2003). Diversidad biológica y sostenibilidad ecológica y económica de los sistemas adehesados. Ecosistemas.. Elmeros, M., A. B. Madsen, et al. (2005). Home range of the badger (Meles meles) in a heterogeneous landscape in Denmark. Lutra 48 (1): 35-44.. Emlen, J. M. (1966). The role of time and energy on food preference. American Naturalist 100: 611-617.. Espírito-Santo, C., L. M. Rosalino, et al. (2007). Factors afecting the placement of common genet latrine sites in a mediterranean landscape in Portugal. Journal of Mammalogy 88: 201–207.. Foster, J. & M. S. Gaines (1991). The effects of a successional habitat mosaic on a small mammal community." Ecology 72(72): 1358-1373.. Fretwell, S. & H. Lucas (1970). On territorial behavior and other factors influencing habitat distribution in birds I: theoretical development. Acta Biotechnologica 19: 16–36.. Fromm, J. E. & W. J. Bell (1987). Search orientation of Musca domestica in patches of sucrose drops. Physiological Entomology 12: 297–307.. Gisbert, J. & M. Santos-Reis (2002). Mustela nivalis. Atlas de los Mamíferos Terrestres de España. J. L. Palomo and J. Gisbert. Madrid, Dirección General de Conservación de la Naturaleza-SECEM-SECEMU.. Gittleman, J. L., S. M. Funk, et al. (2001). Why carnivore conservation? Carnivore conservation. J. L. Gittleman, S. M. Funk, D. W. Macdonald and R. K. Wayne. Cambridge, Cambridge University Press: 1-7.. Gonnet, J. M. (1998). Influencia del pastoreo sobre poblaciones de aves y mamıferos herbıvoros en la region de la reserva de la biosfera ‘Nacunan’, Mendoza, Argentina. Cordoba, Universidad Nacional de Cordoba. 43.
(44) Gonnet, J. M. & R. A. Ojeda (1998). Habitat use by small mammals in the arid andean foothills of the Monte Desert of Mendoza, Argentina." Journal of Arid Environments 38: 349-357.. Hassell, M. P. & T. R. E. Southwood (1978). Foraging strategies of insects. Ecology 9: 75–98.. Hellegers, P. The role of agricultural policy in maintaining high nature value farming systems in Europe. A. E. R. Institute.. Jager, H. I., E. A. Carr, et al. (2006). Simulated effects of habitat loss and fragmentation on a solitary mustelid predator. Ecological Modelling 191: 416430.. Jones, C., W. J. McShea, et al. (1996). Capturing mammals. Measuring and monitoring biological diversity - standard methods for mammals. D. E. Wilson, F. R. Cole, J. D. Nichols, R. Rudran and M. S. Foster. Washington, Smithsonian Institution Press: 115-156.. Jubete, F. (2002). Apodemus sylvaticus. Atlas de los Mamíferos Terrestres de España. J. L. Palomo and J. Gisbert. Madrid, Dirección General de Conservación de la Naturaleza-SECEM-SECEMU.. Kareiva, P. M. (1985). Finding and losing host plants by Phyllotreta: patch size and surrounding habitat. Ecology 66: 1809–1816.. Kendall, K. C., L. H. Metzgar, et al. (1992). Power of sign surveys to monitor population trends." Ecological Applications 2(4): 422-430.. Klein, B. C. (1989). Effects of forest fragmentation on dung and carrion beetle communities in central Amazonia. Ecology 70: 1715–1725.. 44.
(45) Kotler, B. P., Y. Ayal, et al. (1994). Effects of predatory risk and resource renewal on the timing of foraging activity in a gerbil community. Oecologia 100: 391-396.. Krebs, C. J. (1989). Species diversity measures. Ecological methodology, HarperCollins: 328-370.. Lack, D. (1969). The numbers of bird species on islands. Bird Study 16 (193209).. Lacy, R. C. (1993). Impacts of inbreeding in natural and captive populations of vertebrates: implications for conservation. Perspectives in Biology and Medicine 36: 480–496.. Lacy, R. C. (2000). Considering threats to the viability of small populations using individual-based models. Ecological Bulletins 48: 39– 51.. Lande, R. (1993). Risks of population extinction from demographic and environmental stochasticity and random catastrophes. The American Naturalist 142: 911–927.. Law, B. S. & C. R. Dickman (1998). The use of habitat mosaics by terrestrial vertebrate fauna: implications for conservation and management. Biodiversity and Conservation 7: 323-333.. Lawton, J. H. (1993). Range, population abundance and conservation. Trends in Ecology and Evolution 8: 409-413.. Lidicker, W. Z. (1999). Responses of mammals to habitat edges: an overview. Landscape Ecology 14: 333-343.. López-Fuster, M. J. (2002). Crocidura russula. Atlas de los Mamíferos Terrestres de España. J. L. Palomo and J. Gisbert. Madrid, Dirección General de Conservación de la Naturaleza-SECEM-SECEMU. 45.
(46) MacArthur, R. H. & E. R. Pianka (1966). On optimal use of a patchy environment 100: 603-609.. MacArthur, R. H. & E. O. Wilson (1967). The theory of island biogeography. Princeton, Princeton University Press.. Macdonald, D. W. (2005). European mammals. Evolution and behaviour. London, HarperCollins Publishers.. Marshall, E. J. P., T. M. West, et al. (2006). Impacts of an agri-environment field margin prescription on the flora and fauna of arable farmland in different landscapes. Agriculture, Ecosystems and Environment 113: 36-34.. Martin, S. O. (2002). Riparian habitat management for mammals on corps of engineers projects. U. S. Army Engineer Research and Development Center. Vicksburg.. Mathias, M. L. & M. G. Ramalhinho (1999). Insectívoros e Roedores. Caracterização da flora e da fauna do montado da herdade da Ribeira Abaixo (Grândola - Baixo Alentejo). M. Santos-Reis & A. I. Correia. Lisboa, Centro de Biologia Ambiental: 241-248.. McCarthy, M. A., D. C. Franklin, et al. (1994). The importance of demographic uncertainty: an example from the helmeted honeyeater. Biological Conservation 67: 135–142.. McItyre, N. E. & J. A. Wiens (1999). Interactions between landscape structure and animal behavior: the roles of heterogeneously distributed resources and food deprivation on movement patterns. Landscape Ecology 14: 437–447.. Murray, D. L., S. Boutin, et al.. (1994) "Winter habitat selection by lynx and coyote in relation to snowshoe hare abundance." Canadian Journal of Zoology 72: 1444-1451 46.
Imagem
+7
Documentos relacionados
Avaliação do impacto do processo de envelhecimento sobre a capacidade funcional de adultos mais velhos fisicamente ativos.. ConScientiae
forficata recém-colhidas foram tratadas com escarificação mecânica, imersão em ácido sulfúrico concentrado durante 5 e 10 minutos, sementes armazenadas na geladeira (3 ± 1
Water and wastewater treatment produces a signi ficant amount of methane and nitrous oxide, so reducing these emissions is one of the principal challenges for sanitation companies
Foram analisados a relação peso-comprimento e o fator de condição de Brycon opalinus, em três rios do Parque Estadual da Serra do Mar-Núcleo Santa Virgínia, Estado de São
For additional support to design options the structural analysis of the Vila Fria bridge was carried out using a 3D structural numerical model using the finite element method by
ed è una delle cause della permanente ostilità contro il potere da parte dell’opinione pubblica. 2) Oggi non basta più il semplice decentramento amministrativo.
O objetivo do curso foi oportunizar aos participantes, um contato direto com as plantas nativas do Cerrado para identificação de espécies com potencial
libras ou pedagogia com especialização e proficiência em libras 40h 3 Imediato 0821FLET03 FLET Curso de Letras - Língua e Literatura Portuguesa. Estudos literários
