Eletrofisiologia em Neurologia
de Animais de Companhia
Dissertação de Mestrado em Medicina Veterinária
Cláudia Raquel Esteves Soares
Orientador:
Professor Doutor Artur Severo Proença Varejão
II
Eletrofisiologia em Neurologia
de Animais de Companhia
Dissertação de Mestrado em Medicina Veterinária
Cláudia Raquel Esteves Soares
Orientador:
Professor Doutor Artur Severo Proença Varejão
Composição do Júri:
Professora Doutora Felisbina Luísa Pereira Guedes Queiroga
Professora Doutora Justina Maria Prada Oliveira
Professor Doutor Artur Severo Proença Varejão
membrana das suas células. A gravação destas alterações de potencial através de equipamento especializado constitui a base fisiológica dos exames de eletrodiagnóstico. Estes permitem obter informação imediata e objetiva para localizar e caraterizar disfunções do sistema nervoso periférico. Atualmente, a avaliação eletrofisiológica é considerada elemento fundamental no exame diagnóstico de animais com alterações dos nervos periféricos, músculos ou junção neuromuscular.
Foram estudados 8 casos clínicos selecionados com base na história, sinais clínicos e aproximação diagnóstica, incluindo-se casos em que se recorreu a exames eletrofisiológicos. O recurso a estes exames revelou-se bastante útil em todos os casos descritos. Foi possível observar atividade elétrica espontânea em todos eles, a qual é, regra geral, indicadora de uma neuropatia ou miopatia. Por outro lado, a análise dos resultados com o intuito de localizar a lesão, permitiu diferenciar doenças focais (3 casos de tumor) de uma doença generalizada (polineuropatia ou polimiopatia). Foi possível detetar um decréscimo na velocidade de condução num dos 6 casos em que se realizou este tipo de estudo e ausência completa de condução noutros 2, situações compatíveis com uma alteração na mielina. A análise da amplitude dos potenciais obtidos nestes casos permitiu identificar 2 situações de axonopatia associada a desmielinização. No que concerne a avaliação da junção neuromuscular, aplicou-se a técnica de estimulação nervosa repetitiva em 2 casos, a qual, por não revelar alterações, permitiu descartar hipóteses diagnósticas como miastenia gravis ou botulismo. Por último, foi possível testemunhar a aplicabilidade das ondas F na identificação de desmielinização do nervo motor proximal e raiz nervosa dorsal, assim como na localização da lesão.
iv
The nervous system is specialized for conducting changes in the membrane potentials of its cells. The recording of these changes in potential utilizing specialized equipment is the physiological basis of electrodiagnosis. These tests give immediate and objective information to locate and characterize disorders of the peripheral nervous system. Currently, electrophysiological evaluation is a key element in the diagnostic examination of animals with changes in peripheral nerves, muscles and neuromuscular junction.
Eight clinical cases were studied. They were selected on the basis of history, clinical signs and diagnostic approach, including cases in which electrophysiological tests were used. The application of these tests has proved very useful in all the cases described. We observed spontaneous electrical activity in all of them, which is generally indicative of a neuropathy or myopathy. Moreover, the analysis of the results in order to locate the lesion, allowed to differentiate between focal diseases (3 cases of tumors) and generalized disease (polyneuropathies and polymiopathies). It was possible to detect a decrease in conduction velocity in 1 of the 6 cases held in this type of study, and complete absence of conduction was detected in other 2 cases. The amplitude analysis of the potentials obtained in these 2 cases revealed axonopathy associated with demyelination. Regarding the evaluation of the neuromuscular junction, the technique of repetitive nerve stimulation was applied in 2 cases, which, by not disclosing changes, allowed to discard some diagnostic hypothesis as myasthenia gravis or botulism. Finally, it was possible to witness the applicability of F waves to identify demyelination of motor nerve and proximal dorsal nerve root, as well as the location of the lesion.
v
CAPÍTULO I – REVISÃO BIBLIOGRÁFICA ... 1
1. MÉTODOSDEPESQUISAESELEÇÃO ... 1
2. BASEFISIOLÓGICAEPRINCÍPIOSGERAIS... 1
2.1. Potenciais de Membrana em Repouso ... 1
2.2. Potenciais de Ação ... 1
2.3. Potenciais Pós-sináticos ... 2
2.4. O Sistema Nervoso Periférico e as suas Disfunções ... 2
2.5. Aspetos técnicos da gravação da atividade elétrica ... 3
2.5.1. Amplificador Diferencial ... 4
2.5.2. Tipos de Elétrodos ... 5
2.5.3. Atividade Voluntária, Espontânea e Evocada ... 6
2.5.4. Sinais desencadeados e não desencadeados ... 6
2.5.5. Média dos Sinais ... 6
2.5.6. Calibragem ... 7
2.5.7. Monitorização Áudio ... 7
3. OSTESTESDEELETRODIAGNÓSTICOEMMEDICINAVETERINÁRIA ... 7
4. ATIVIDADEELÉTRICANOMÚSCULO... 9
4.1. Eletromiografia ... 9
4.2. Músculo em Repouso ... 10
4.2.1. Atividade de Inserção ... 10
4.2.2. Potenciais Miniatura de Placa Terminal ... 11
4.2.3. Potenciais da Unidade Motora ... 12
4.3. Atividade Espontânea Anormal... 12
4.3.1. Fasciculação... 14
4.3.2. Potenciais de Fibrilhação ... 14
4.3.3. Ondas Agudas Positivas ... 15
4.3.4. Descargas Repetitivas Complexas ... 15
4.3.5. Potenciais Miotónicos ... 16
4.4. Atividade Evocada no Músculo ... 16
4.4.1. Atividade Evocada Eletricamente ... 17
4.4.2. Atividade Evocada por Estimulação Magnética ... 17
4.4.3. Onda M ... 18
4.4.4. Estimulação Nervosa Repetitiva ... 19
4.4.5. Velocidade de Condução Nervosa ... 21
vi
5. ATIVIDADEELÉTRICANOSNERVOSPERIFÉRICOS ... 31
5.1. Atividade Espontânea ... 31
5.1.1. Eletroneurografia ... 31
5.2. Atividade Evocada ... 31
CAPÍTULO II – APRESENTAÇÃO DOS CASOS CLÍNICOS ... 34
1. MATERIALEMÉTODOS ... 34
2. CASOSCLÍNICOS ... 37
CAPÍTULO III – DISCUSSÃO ... 57
vii
Figura 2 – Posicionamento do animal em decúbito lateral e numa superfície acolchoada Figura 3 (A – E) – Exame eletromiográfico. Elétrodo de EMG em agulha concêntrico e fio-terra Figura 4 (A e B) – Estudo dos potenciais elétricos através do seu aspeto no monitor e do áudio
do altifalante
Figura 5 (A e B) – Posicionamento dos elétrodos para estudos de condução nervosa no
membro pélvico
Figura 6 – Ondas F obtidas a partir da estimulação do n. tibial esquerdo ao nível da articulação
tibiotársica. Latência onda F = 35,8 ms. Observa-se polifasia e dispersão temporal
Figura 7 – Ondas F obtidas a partir da estimulação do n. tibial direito ao nível da articulação
tibiotársica. Latência onda F = 21,4 ms
Figura 8 – Fibrilhação de densidade moderada detetada no exame eletromiográfico do m.
extensor carpo-radial esquerdo
Figura 9 – Fibrilhação grave detetada no exame eletromiográfico do m. bíceps braquial
esquerdo
Figura 10 – Estudo de condução nervosa do MPE. Potenciais obtidos a partir do m. interósseo
plantar, após estimulação do nervo ciático ao nível do grande trocânter (A1) e inserção do tendão gastrocnémio – tuberosidade calcânea (A2). VCN = 78 m/s
Figura 11 – Fibrilhação moderada detetada no exame eletromiográfico do m. temporal
esquerdo
Figura 12 – Estudo de condução nervosa motora do MPE. Potenciais obtidos a partir do m.
interósseo plantar após estimulação do nervo tibial ao nível da articulação tibiotársica (A1), inserção do tendão gastrocnémio – tuberosidade calcânea (A2) e grande trocânter (A3).
VCN = 42, 50 e 46 m/s, respetivamente.
Figura 13 – Estudo de condução nervosa motora do MAE. Potenciais obtidos a partir do m.
extensor digital comum, após estimulação do nervo radial ao nível do antebraço (A1) – ausência de potenciais –zona lateral do úmero (A2) e axila (A3). VCN = 103 m/s
Figura 14 – Estudo de condução nervosa sensorial do MAE. Potenciais obtidos ao nível medial
do cotovelo após estimulação do nervo ulnar
Figura 15 – Fibrilhação suave ou de densidade baixa detetada no exame eletromiográfico do
viii
Figura 17 – Estudo de condução nervosa do MAD com estimulação do nervo radial ao nível do
antebraço (A1) – ausência de potenciais, zona distal lateral do úmero (A2) – potenciais de baixa amplitude (0,7 µ), e plexo braquial (A3) – ausência de potenciais
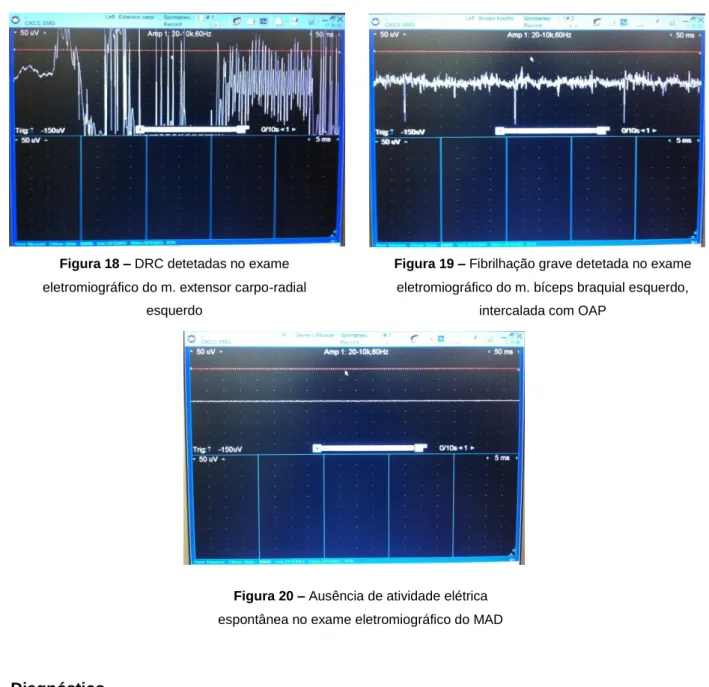
Figura 18 – DRC detetadas no exame eletromiográfico do m. extensor carpo-radial esquerdo Figura 19 – Fibrilhação grave detetada no exame eletromiográfico do m. bíceps braquial
esquerdo, intercalada com OAP
Figura 20 – Ausência de atividade elétrica espontânea no exame eletromiográfico do MAD Figura 21 – Atividade de inserção detetada no exame eletromiográfico do m. tibial anterior
direito
Figura 22 – Fibrilhação grave detetada no exame eletromiográfico do m. gastrocnémio direito,
intercalada com OAP
Figura 23 – Estudo de condução nervosa do MP. Potenciais obtidos a partir do m. tibial cranial
após estimulação do n. peroneal ao nível da inserção do tendão gastrocnémio – tuberosidade calcânea (A1) e grande trocânter (A2). VCN = 69 m/s
Figura 24 – Fibrilhação de densidade moderada detetada no exame eletromiográfico do m.
flexor carpo-radial esquerdo, intercalada com OAP
Figura 25 – Estudo de condução nervosa motora do MAD. Potenciais obtidos a partir do m.
interósseo palmar após estimulação do nervo ulnar ao nível da articulação carpo-radioulnar
(A1) e úmero-radioulnar (A2). VCN = 91 m/s
Figura 26 – Estudo de condução nervosa sensorial do MPD. Potenciais obtidos a partir da
ix Ac - Anticorpos
ACh – Acetilcolina
ALT – Alanina aminotransferase AST – Aspartato aminotransferase BID – Duas vezes por dia
CI – Cavilha intramedular CK – Creatinina quinase cm - Centímetro
DRC – Descargas repetitivas complexas EMG - Eletromiografia
EMGFU – Eletromiografia de fibra única FA – Fosfatase alcalina
FeCoV – Coronavírus felino FelV – Vírus da leucemia felina FIV – Vírus da imunodeficiência felina G – Gauge
H-E – Hematoxilina & eosina Hz - Hertz i.e. – isto é IM – Intramuscular IV – Intravenoso K+ - Potássio kg – Quilograma L – Litro LCR – Líquido cefalorraquidiano MAD – Membro anterior direito MAE – Membro anterior esquerdo ME – Medula espinhal mg - Miligrama min - Minuto mm - Milímetro m – Músculo mmol - Milimole
MPD – Membro posterior direito
MPE – Membro posterior esquerdo ms – Milissegundo
mV – MiliVolt n - Nervo Na+ - Sódio
OAP – Onda aguda positiva p.e. – Por exemplo
PA – Potencial de ação
PAC – Potencial de ação composto PANS – Potencial de ação nervosa sensorial
PF – Potencial de fibrilhação
PMAC – Potencial muscular de ação composto
PME – Potencial muscular evocado PMPT – Potencial miniatura de placa terminal
PO – per os
PPS – Potencial pós-sinático PPSE – Potenciais pós-sináticos excitatórios
PPSI – Potenciais pós-sináticos inibitórios PPT – Pico de placa terminal
PUM – Potencial da unidade motora QOD – A cada dois dias
ref - Referência
RM – Ressonância magnética
RMDC - Radiculomielopatia degenerativa crónica
RPL – Reflexo pupilar à luz s - Segundo
SID – Uma vez por dia
SNA – Sistema nervoso autónomo SNC – Sistema nervoso central
x T4 – Tirosina
TC – Tempo de condução TC – Tomografia computorizada TSH – Hormona estimulante da tiróide UI – Unidades internacionais
UM – Unidade motora
VCN – Velocidade de condução nervosa
VCNS – Velocidade de condução nervosa sensorial
% - Percentagem µg – Micrograma ºC – Graus Celsius ™ - marca registada
xi
acompanhamento prestado e pelo voto de confiança na escolha do tema.
Ao Hospital Veterinário Montenegro e a toda a equipa pela disponibilidade e amabilidade com que me receberam e pelos conhecimentos transmitidos. A todos os colegas estagiários, companheiros de percurso, pelo apoio e amizade.
Ao Queen’s Veterinary School Hospital e à Universidade de Cambridge pelo simpático acolhimento, confiança e incentivo por parte de todos. Um agradecimento especial à Heidi, Susana, Filipe e Ricardo que me fizeram sentir em casa. À An, Ed e Paul, referências como profissionais, pela disponibilidade e generosidade com que auxiliaram na realização deste trabalho.
Aos colegas de turma e de curso que tiveram um papel importante durante o percurso académico. Agradeço o carinho e a troca de experiências.
À “Família Araucária” que tornou a casa longe de casa num lar e terá sempre um lugar especial na minha vida.
Aos amigos de sempre pela presença constante e indispensável companhia e apoio. Ao meu pai pela confiança e motivação. À minha mãe, amiga, companheira e exemplo. Ao meu avô Delfim que sempre acreditou e incentivou. À minha avó Carolina, a responsável por tudo e “culpada” do sonho.
1
CAPÍTULO I – REVISÃO BIBLIOGRÁFICA 1. MÉTODOS DE PESQUISA E SELEÇÃO
As publicações clínicas relacionadas com o tema “Eletrofisiologia em neurologia de animais de companhia” foram pesquisadas na “Medline”, com “palavras-chave” adequadas como “eletrofisiologia”, “estudo eletrofisiológico”, “eletromiografia”, “estudo de condução nervosa”, “doença neuromuscular”, “neuropatia”, “radiculopatia”, “miopatia”, “cão” e “gato”, com maior incidência na última década.
Os títulos e resumos foram examinados, e foi feita uma seleção adequada dos respetivos conteúdos. A pesquisa nas ciências básicas foi restrita aos artigos que se aproximassem das aplicações clínicas.
2. BASE FISIOLÓGICA E PRINCÍPIOS GERAIS
O sistema nervoso é constituído por 2 tipos principais de células: os neurónios e as células da glia (Kay, 1998; Reece, 2005), sendo os primeiros considerados a “força de trabalho” do sistema. Geram e conduzem potenciais de ação (PA) que são o meio através do qual a informação é transmitida no sistema nervoso (Kay, 1998; Srenk et al., 2010).
2.1. Potenciais de Membrana em Repouso
As membranas fosfolipídicas das células musculares e nervosas separam fluidos intra e extracelulares que diferem na sua composição. Separam também moléculas carregadas, de forma que a voltagem no interior das células é 70 a 90 milivolt (mV) negativa em relação ao exterior. Este gradiente elétrico designa-se “potencial de membrana em repouso” (Reece, 2005).
2.2. Potenciais de Ação
Quando estímulos de intensidade suficiente são aplicados a células excitatórias, o potencial de membrana é invertido – despolarização - e recupera espontaneamente - repolarização. Estas alterações são provocadas por um influxo de sódio (Na+) – despolarização - seguido por um efluxo de potássio (K+) - repolarização. Os fluxos de iões são iniciados por alterações na permeabilidade da membrana associadas a mudanças em canais iónicos específicos localizados no interior da membrana (Reece, 2005). A tendência do Na+ e K+ para se deslocarem contra o seu gradiente eletroquímico através de canais abertos é a base do PA que é propagado ao longo do axolema (por condução contínua ou saltatória) ou do sarcolema (Steiss, 2003).
2
2.3. Potenciais Pós-sináticos (PPS)
A sinapse é o meio de comunicação usual entre células excitatórias (Reece, 2005; Srenk et
al., 2010). A sinapse entre um neurónio e o músculo esquelético designa-se “junção
mioneuronal” ou “neuromuscular”. Em resposta a neurotransmissores, a membrana pós-juncional dos neurónios pode produzir potenciais pós-sináticos excitatórios (PPSE) ou potenciais pós-sináticos inibitórios (PPSI) (Srenk et al., 2010). A excitação pós-juncional no músculo esquelético como resposta à libertação de acetilcolina (ACh) pelo terminal motor do axónio, é designada como “potencial de placa terminal”. Trata-se de um potencial local e não propagável (Steiss, 2003).
2.4. O Sistema Nervoso Periférico e as suas Disfunções
O sistema nervoso periférico (SNP) engloba o sistema neuromuscular, o sistema nervoso sensorial (SNS) e o sistema nervoso autónomo (SNA). O sistema neuromuscular é responsável pela condução do impulso do sistema nervoso central (SNC) para o sistema músculo-esquelético, e é composto por unidades motoras (UM) (Steiss, 2003; Anor, 2007). Os componentes essenciais da UM incluem: um corpo celular do neurónio (localizado no SNC; também designado motoneurónio inferior) e o respetivo axónio periférico, apoiado por células de Schwann, a junção neuromuscular e as fibras musculares inervadas pelo neurónio motor (miofibras) (LeCouteur, 2005; Srenk et al., 2010).
Uma alteração em qualquer parte da UM pode resultar em sinais de doença neuromuscular (LeCouteur, 2005). Nas disfunções da UM incluem-se aquelas que afetam corpos celulares de neurónios localizados no corno ventral da medula espinhal (ME) (neuronopatias), as raízes nervosas (radiculopatias), os nervos periféricos (neuropatias), a junção neuromuscular, e músculos esqueléticos (miopatias) (Jeffery, 1999). Quando as alterações incluem tanto os neurónios como as fibras musculares designam-se neuromiopatias (LeCouteur, 2005).
O SNS é constituído por neurónios sensoriais com uma zona dendrítica modificada para formar recetores. Estes recetores recebem e transformam várias modalidades sensoriais em atividade elétrica, na forma de um PA que viaja a partir da zona dendrítica, pelo axónio, através do gânglio segmentar da coluna vertebral (corpo celular neuronal) na raiz dorsal, e, finalmente entram na ME. As modalidades sensoriais incluem nociceção, proprioceção, sensibilidade, temperatura, e modalidades especiais, tais como a visão, paladar e audição. Tal como ocorre com a UM, lesões em qualquer localização ao longo das vias sensoriais podem resultar em deficiências sensoriais (Anor, 2007).
Os nervos periféricos são compostos por axónios motores e sensoriais (mielinizados e não mielinizados) e são essenciais no funcionamento do SNA e voluntário (Glass e Kent, 2002, Srenk et al., 2010). Algumas neuropatias caraterizam-se por uma disfunção predominante ou
3
exclusivamente motora, outras por uma disfunção sensorial, e outras por uma combinação de sinais motores e sensoriais. Apesar de existirem neuropatias puramente sensoriais, a maioria das neuropatias periféricas originam sinais clínicos que refletem alterações das UM inferiores, como tónus muscular diminuído, diminuição dos reflexos miotáticos e flexores ou de retirada (Glass e Kent, 2002). É comum a paresia manifestada sob a forma de fraqueza. A disfunção de motoneurónio inferior resulta, geralmente, numa marcha curta com flexão exagerada das articulações, e perda da capacidade de sustentação de peso (Olby, 2004). Gatos com diversas disfunções neuromusculares apresentam, com frequência, uma postura alterada com flexão da cabeça e pescoço (Olby, 2004; LeCouteur, 2005).
Em doenças neuromusculares que afetam apenas UM inferiores e poupam os nervos sensoriais, as reações posturais permanecem normais, contanto que o animal possua força para suportar o próprio peso. Assim que a fraqueza de motoneurónio inferior se torna marcada, a paresia pode afetar a capacidade do clínico para avaliar as reações posturais porque o animal está demasiado fraco para efetuar reações normais (Anor, 2007). Disfunções no nervo laríngeo recorrente provocam alterações da voz ou perda do ladrar e aumento dos sons respiratórios (estridores respiratórios). Alterações concomitantes dos músculos da faringe e laringe resultam, habitualmente, no aparecimento de megaesófago e, subsequente pneumonia por aspiração (Olby, 2004). Doenças que afetam o SNP sensorial causam ataxia propriocetiva e diminuição da perceção da dor ou sensação (Anor, 2007).
Apesar de as manifestações clínicas de neuropatias periféricas estarem, geralmente, relacionadas com alterações dos nervos periféricos, os nervos craniais podem também ser afetados, surgindo sinais como paresia facial ou de doença vestibular (Platt e Garosi, 2004). Desenvolve-se rapidamente atrofia muscular grave e generalizada. Contudo, algumas neuropatias e miopatias podem originar hipertrofia muscular (Platt e Garosi, 2004; LeCouteur, 2005).
As miopatias podem distinguir-se das neuropatias porque, habitualmente, não estão associadas a défices nas reações posturais ou diminuição dos reflexos miotáticos. A intolerância ao exercício pode ser o único sinal clínico observado em algumas miopatias e disfunções da junção neuromuscular, como miastenia gravis. Contudo, outras alterações, como o botulismo, causam muitas vezes fraqueza ascendente de início agudo, que progride dos membros pélvicos para os torácicos (Platt e Shelton, 2004).
2.5. Aspetos técnicos da gravação da atividade elétrica
Os PA caraterizam-se por parâmetros como amplitude, duração, número de fases e polaridade, frequência, repetição e velocidade de condução (LeCouteur, 2005). A amplitude de um PA a partir de uma única célula é medida em mV, e pode perder-se completamente se o
4
elétrodo de gravação não se encontrar perto da célula ativa e/ou se o ganho do amplificador for demasiado baixo. Desta forma, a maioria dos procedimentos clínicos de eletrodiagnóstico baseia-se em gravações de alterações transmembranares de um grande número de células (Steiss, 2003).
Quando as células excitatórias são ativadas simultaneamente, pode ser gravado um potencial de ação composto (PAC). A amplitude de um PAC é dependente do número de células participantes e respetivas amplitudes. A duração do PAC é um reflexo de sincronia. Células semelhantes com descargas em simultâneo produzem uma breve duração, enquanto uma longa duração será causada por células diferentes que despolarizam de forma sincrónica ou células semelhantes a despolarizar em assincronia (Steiss, 2003).
Em gravações eletrodiagnósticas, os termos “potencial” ou “resposta” podem ser utilizados em referência a um PA único ou composto, ou uma combinação de PA e PPS (Steiss, 2003).
Quando várias células nos tecidos estão ativas, as respostas podem ser detetadas por dispositivos de gravação muito sensíveis. Regra geral, recorre-se a um sistema informático em interface com amplificadores diferenciais, filtros, estimulador e um dispositivo que calcula a média dos sinais (Sims, 1996).
2.5.1. Amplificador Diferencial
Devido ao pequeno tamanho dos sinais bioelétricos, torna-se necessário recorrer a amplificação (Dumitru e Zwartz, 2001). A amplificação de sinais biológicos é, geralmente, conseguida através de um amplificador diferencial que possui duas entradas, sendo a magnitude de saída representa a diferença entre as duas. Para muitos procedimentos de eletrodiagnóstico, as entradas são obtidas a partir de um elétrodo ativo e outro de referência. O primeiro é posicionado na área onde decorre a atividade que se pretende gravar e o segundo numa área distante e inativa. Uma vez que o amplificador diferencial atenua a atividade que é comum às duas entradas, ajuda a eliminar atividade elétrica de fundo indesejada ou “ruído”, a qual representa uma fonte de artefactos (Steiss, 2003).
O alcance de frequência no qual um amplificador pode atuar sem distorção designa-se “resposta de frequência”. Filtros de baixa e alta frequência permitem que a resposta de frequência possa ser ajustada para uma gravação ideal da resposta. Trata-se de dispositivos eletrónicos que removem componentes da onda a frequências específicas (Dumitru e Walsh, 1988, Dumitru e Zwartz, 2001; Oh, 2002). Para cada tipo de procedimento eletrodiagnóstico, devem utilizar-se, de forma consistente, as definições de filtro adequadas. Definições incorretas podem alterar a amplitude, forma e/ou latência das respostas (Koo et al., 2012).
5 2.5.2. Tipos de Elétrodos
Estão disponíveis 4 tipos principais de elétrodos de gravação para EMG. Os elétrodos de agulha concêntricos e os monopolares são os 2 tipos mais comummente utilizados em Medicina Veterinária (Bowen, 1987). Apesar da sua ampla utilização em Medicina Humana, os elétrodos de superfície não são adequados para uso em Veterinária devido às dificuldades técnicas associadas ao fraco contacto entre a pele e o elétrodo, como resultado da presença de pelo (Cuddon, 2002). Assim, recorre-se geralmente a elétrodos de agulha (Sims, 1996), apesar de também serem usados elétrodos adesivos, discos de metal e pinças crocodilo (Feitosa et al., 2002). Pequenos elétrodos (27G) podem ser aplicados subcutaneamente e agulhas isoladas com pontas simples podem ser inseridas nos músculos ou colocadas junto aos nervos (Sims, 1996).
O elétrodo concêntrico (coaxial) mede a diferença de potencial entre um fio de nicromo, prata ou platina e o eixo de aço-inoxidável que o rodeia (Bowen, 1987). O fio central do elétrodo (elétrodo ativo ou de exploração) é isolado da cânula que o rodeia (elétrodo de referência), de modo que a área que efetivamente é testada (entre os 2) seja bastante pequena (Cuddon, 2002; LeCouteur e Williams, 2012). Os elétrodos monopolares são compostos de isolamento de aço inoxidável, exceto na ponta e são referenciados para um segundo elétrodo noutro local (Bowen, 1987). Os elétrodos bipolares têm, tanto o elétrodo ativo como o de referência, incorporados no centro da agulha (Steiss, 2003). Este último tipo é usado raramente em Medicina Veterinária (Cuddon, 2002).
Cada um destes tipos de elétrodos apresenta vantagens e desvantagens. O elétrodo concêntrico deteta menos ruído de fundo que os elétrodos monopolares, e identifica potenciais de fibrilhação mais frequentemente, uma vez que estes potenciais são induzidos mais comummente pela inserção da agulha e não por ocorrência espontânea. Pelo facto de estes elétrodos provocarem um maior dano nos tecidos que os elétrodos monopolares, a deteção dos potenciais de fibrilhação é facilitada. Os elétrodos monopolares são menos estáveis eletricamente e mais ruidosos, embora produzam menos dor e gravem um potencial consideravelmente maior a partir da mesma fonte, comparativamente a um elétrodo concêntrico (Bowen, 1987; Sherman, 1990). São também capazes de detetar potenciais distantes, como resultado da sua substancial distância entre elétrodos (Sherman, 1990).
O tipo e a localização do elétrodo são críticos para o tipo específico de gravação (Cuddon, 2002). Os elétrodos de superfície são geralmente considerados melhores que os elétrodos de agulha para a gravação de PMAC, uma vez que avaliam as contribuições de todas as unidades em descarga (Kimura, 1981c). Os elétrodos de agulha registam apenas uma pequena percentagem dos PMAC potencialmente graváveis. Para além disso, o posicionamento das agulhas torna-se geralmente mais crítico, já que um menor deslocamento pode resultar numa
6
substancial alteração do tamanho e forma do PMAC (Kimura, 1981c). Apesar disso, os elétrodos de agulha são considerados preferíveis uma vez que são mais fáceis de usar, especialmente no que respeita à colocação e ajuste num músculo em particular. Por outro lado, melhoram a gravação a partir de músculos pequenos e atróficos (i.e., músculos interósseos), que são os músculos usados por rotina em estudos de condução nervosa motora (Cuddon, 2002). Os elétrodos de agulha são considerados melhores que os de superfície em estudos de condução nervosa sensorial, já que melhoram significativamente a razão sinal-ruído (Kimura, 1981c).
2.5.3. Atividade Voluntária, Espontânea e Evocada
A atividade elétrica gravada a partir de tecido excitável pode ser voluntária, espontânea ou evocada. A atividade voluntária ocorre quando o animal conscientemente realiza alguma atividade, como mover um membro. A atividade espontânea pode ser gravada sem participação voluntária ou o uso de um estímulo externo, p.e., num eletroencefalograma. As respostas evocadas ou potenciais evocados representam a resposta elétrica a um estímulo externo, normalmente recorrendo-se a uma intensidade e taxa específicas (frequência). Com a exceção dos testes dos sistemas olfativo, visual e auditivo, a maioria das respostas evocadas é obtida através de pulsos curtos de corrente elétrica (Steiss, 2003).
2.5.4. Sinais desencadeados e não desencadeados
As respostas elétricas que ocorrem através das membranas podem ser gravadas a partir de um tecido espontaneamente ativo ou evocadas através da aplicação adequada de estímulos. A aplicação de um estímulo pode ser usada para desencadear o início de gravações repetitivas de atividade fisiológica para janelas pré-selecionadas de tempo. Uma vez que as respostas evocadas estão geralmente associadas no tempo ao estímulo, a visualização da resposta é amplamente melhorada pelo seu desencadeamento (Sims, 1996).
Nos potenciais evocados, os amplificadores são ativados pelo estímulo a iniciar a gravação, durante um periodo de tempo pré-determinado, que se designa “tempo de análise”. Em alguns protocolos utiliza-se um circuito de atraso por forma a iniciar a gravação num tempo específico, após a aplicação do estímulo. O tempo em que o amplificador grava a atividade denomina-se “janela”. Uma gravação da atividade espontânea normalmente requer uma janela de gravação mais ampla (Steiss, 2003).
2.5.5. Média dos Sinais
Muitas respostas evocadas são extremamente pequenas, com uma relação sinal-ruído pobre. A “média dos sinais” é uma técnica amplamente utilizada para aumentar o biopotencial e
7
reduzir o ruído (Holliday et al., 1979; Farnbach, 1981; Redding et al., 1982; Bowen, 1986). Com a obtenção eletrónica da média de múltiplas respostas, a amplitude da resposta evocada aumenta em proporção direta ao número de amostras. O ruído de fundo, que é aleatório, diminui pela raiz quadrada do número de respostas. O número de repetições que precisam de ser incluídas na média é determinado por fatores tais como a amplitude de resposta e a quantidade de ruído. Embora se possa utilizar um número definido de repetições num protocolo, fica a cargo do examinador a determinação do momento em que uma resposta ótima foi registada. Um maior número de repetições não iguala maior qualidade de gravação. Em alguns casos, a utilização de um grande número de repetições pode diminuir a qualidade da gravação (Steiss, 2003). Com a média dos sinais, a voltagem do ruído que muda aleatoriamente a sua relação no tempo com o estímulo (i.e, ruído de fundo), é arredondada a zero em cada ponto no tempo após o estímulo. Pelo contrário, os sinais que estão associados no tempo ao estímulo somam a uma latência constante e aparecem como um potencial evocado (Holliday et al., 1979; Farnbach, 1981; Redding et al., 1982; Bowen, 1986).
2.5.6. Calibragem
Os biopotenciais aqui discutidos são principalmente de ocorrência rápida e com sinais de baixa amplitude. Uma vez que muitos estão longe do campo de gravação, são bastante menores quando comparados aos potenciais registados diretamente a partir de células individuais com microelétrodos. A maioria do equipamento de eletrodiagnóstico é desenhado para captar potenciais na gama dos mV e microvolts (μV). As calibragens são realizadas, geralmente, em milissegundos (ms) e usam-se programas informáticos que permitem medições precisas (Steiss, 2003). Mesmo após o cálculo do sinal médio e da filtragem, a maioria das respostas é ainda bastante pequena (na ordem dos mV ou µV) e exige um amplificador de alto ganho para a sua gravação (Sims, 1996), como foi já referido.
2.5.7. Monitorização Áudio
Em eletromiografia (EMG) recorre-se à monitorização áudio dos sinais. Os biopotenciais são recolhidos do amplificador do sistema e conduzidos a um amplificador de áudio e altifalante (LeCouteur, 2005; LeCouteur e Williams, 2012). Isto permite ao operador identificar potenciais pelo seu som caraterístico, além do seu aspeto no monitor (Srenk et al., 2010; LeCouteur e Williams, 2012).
3. OS TESTES DE ELETRODIAGNÓSTICO EM MEDICINA VETERINÁRIA
Os testes de eletrodiagnóstico foram introduzidos em Medicina Veterinária em 1949 (Allam,
8
gravação da atividade elétrica dos músculos, a EMG não se desenvolveu como um campo independente até 1967 (Botello et al. 1967).
A avaliação eletrofisiológica tornou-se a pedra angular no exame diagnóstico de animais com alterações dos nervos periféricos, músculos ou junção neuromuscular (Cuddon, 2002; Koo et al. 2012; Krarup e Crone, 2002). As correntes elétricas produzidas pelas células excitatórias no sistema nervoso, quando ativas, fornecem a base fisiológica para estes estudos (Sims, 1996) e permitem obter informação imediata e objetiva (Poncelet, 2004), não só para localizar, mas também para caraterizar disfunções do SNP (Srenk et al., 2010; Koo et al. 2012).
A condução dos impulsos nervosos depende da integridade do corpo celular neuronal, axónio, bainha de mielina e junção entre o neurónio e o seu axónio e o alvo (músculo ou órgão inervado). A velocidade de condução do impulso nervoso está relacionada, em grande medida, com o diâmetro do axónio (quanto maior o axónio, mais rápida é a condução). A mielina produzida pelas células de Schwann no SNP isola os axónios, aumentando a resistência da sua membrana e originando uma condução do impulso de nó em nó (regiões de membrana exposta), o que aumenta de forma dramática a velocidade de condução (Olby, 2004; Srenk et
al., 2010). Os neurónios do SNP são mais resistentes à lesão que os seus homólogos do SNC
e são capazes de regenerar axónios a uma velocidade surpreendentemente elevada (1-4 mm/dia), se a regeneração ocorrer num tubo de células do endonervo e de Schwann intactas (Olby, 2004).
Apesar de se aplicarem aos nervos periféricos os principais tipos de lesão que afetam o SNC, as alterações metabólicas, hereditárias, degenerativas e tóxicas apresentam uma maior importância clínica no SNP. Na maioria das neuropatias periféricas, independentemente da causa, está presente uma mistura de desmielinização e degenerescência axonal. As extremidades distais dos axónios são particularmente suscetíveis à degenerescência, como resultado da sua distância ao corpo celular neuronal. Os nervos mais longos (laríngeo recorrente, ciático) são frequentemente os primeiros afetados em alterações tóxicas, degenerativas e metabólicas (Olby, 2004, Anor, 2007).
Uma avaliação eletrofisiológica cuidada consiste em EMG, estudos de condução motora e sensorial de nervos periféricos, avaliação da junção neuromuscular via estimulação supramaximal repetitiva ou eletromiografia de fibra única, e ainda avaliação das porções proximais de nervos periféricos e raízes nervosas dorsais e ventrais através dos potenciais evocados da ME e ondas F, respetivamente (Cuddon, 2002).
9
4. ATIVIDADE ELÉTRICA NO MÚSCULO 4.1. Eletromiografia
Por definição, electromiografia (EMG) é a gravação e estudo de atividade elétrica de inserção, espontânea e voluntária no músculo (Bowen, 1978; Farnback, 1980; Kimura, 1989). Devido à dificuldade de cooperação por parte dos doentes, em pequenos animais, por rotina, não se realizam estes estudos em animais acordados (Cuddon, 2002; LeCouteur e Williams, 2012). O recurso a anestesia ou sedação permite eliminar atividade voluntária indesejada e artefactos associados ao movimento (Steiss, 2003). A atividade muscular voluntária não pode ser avaliada no animal anestesiado, mas pode registar-se a presença de atividade elétrica espontânea e o seu tipo e distribuição podem ajudar no estabelecimento de um diagnóstico ou simplesmente identificar os músculos afetados (Cuddon, 2002; Srenk et al., 2010).
Os dados de uma EMG fornecem um diagnóstico específico em doenças em que ocorre contração involuntária do músculo, como a miotonia. Miopatias tão diversas como distrofia muscular ou polimiosite apresentam alterações da EMG menos específicas que facilitam o mapeamento da extensão da doença (Olby, 2005).
A EMG pode também ajudar na seleção de tecido para a colheita de biópsias. Os músculos que revelam sinais moderados são ideais para uma comparação entre miofibras afetadas e não afetadas. As biópsias são geralmente realizadas no lado do animal que não foi submetido a avaliação eletromiográfica. Em situações generalizadas, assume-se que as alterações serão simétricas (LeCouteur e Williams, 2012). Em humanos, a contribuição diagnóstica da EMG de agulha fina em casos de miastenia gravis é superior à da determinação de anticorpos antirrecetores ACh no soro ou estimulação nervosa repetitiva supramaximal (Sanders e Howard, 1986).
Todas as avaliações eletromiográficas estão sujeitas a dificuldades técnicas associadas a ruído externo de outros equipamentos elétricos ou de anestesia na mesma sala e, idealmente, devem ser realizadas numa sala Faraday. O animal deve ser colocado numa superfície acolchoada, conetado à máquina com um fio-terra, e num ambiente quente para evitar alterações artefactuais nos resultados (Braund, 1994).
A sensibilidade do exame eletromiográfico aumenta dramaticamente com o aumento do número de locais examinados em cada músculo, e também com o número de músculos analisados. A atividade muscular espontânea anormal é, geralmente, um fenómeno desigual que varia consideravelmente com a localização (Cuddon, 2002). Variando a profundidade e o ângulo do elétrodo, podem examinar-se diferentes locais no músculo com apenas uma inserção da agulha através da pele. O tamanho e espessura de cada músculo são determinantes para o número de locais que podem ser avaliados (LeCouteur e Williams, 2012).
10
A seleção dos músculos para avaliação eletromiográfica varia de acordo com os sinais clínicos do animal em questão e a lista de diagnósticos provisórios. Para uma situação generalizada, analisam-se diversos músculos, sendo, por rotina, avaliados os músculos apendiculares proximais e distais, bem como diversos músculos axiais. Nestes casos, os testes são normalmente reservados a um dos lados do animal, sendo o outro preservado para avaliação histopatológica. Em casos selecionados, os músculos da faringe, laringe, língua e esfíncter anal podem também ser sujeitos a avaliação eletromiográfica (LeCouteur e Williams, 2012).
4.2. Músculo em Repouso
A atividade espontânea no músculo, quando ocorre, pode ser iniciada no motoneurónio inferior, na raiz nervosa, no axónio periférico ou na própria membrana muscular (Steiss, 2003). Se o músculo esquelético não for ativado de forma voluntária ou reflexa, é eletricamente quiescente (Cuddon, 2002; LeCouteur, 2005). A maioria da atividade espontânea é indicativa de uma alteração neuromuscular (Steiss, 2003).
4.2.1. Atividade de Inserção
A inserção ou manipulação de um elétrodo de agulha no músculo saudável é acompanhada por atividade elétrica designada “potenciais de inserção”. Estes potenciais têm origem na estimulação mecânica das fibras musculares e, geralmente param assim que o movimento da agulha termina (Cuddon, 2002; LeCouteur, 2005; LeCouteur e Williams, 2012).
A atividade de inserção pode ser usada para determinar a excitabilidade muscular (Cuddon, 2002). Consiste em picos positivos e negativos de alta frequência que ocorrem em aglomerado, produzindo um som nítido e estático (Kimura, 1981). No músculo normal, esta atividade encontra-se na ordem das poucas centenas de ms (5 - 300 ms), dependendo da velocidade e magnitude do movimento do elétrodo (Kimura, 1981a; Srenk et al., 2010), e apresenta uma amplitude de 100 µV (Srenk et al., 2010). A atividade de inserção pode estar aumentada em alterações neuropáticas e miopáticas e surgir em conjunto com outros potenciais anormais (Steiss, 2003; LeCouteur, 2005), particularmente em doenças que causem instabilidade da membrana muscular, como desinervação ou inflamação (Kimura, 1981a; Kimura, 1981b; Bowen, 1987; Srenk et al., 2010). As fibras musculares afetadas estão hiperexcitáveis, tendo um limiar mais baixo, o qual se deve a potenciais de membrana diminuídos. Uma atividade de inserção prolongada pode ser a única alteração observada em estadios iniciais de doença neuromuscular (LeCouteur e Williams, 2012).
Pelo contrário, em músculos com atrofia grave ou fibrose, a atividade de inserção pode estar reduzida, devido à diminuição no número de miofibras disponíveis (Kimura, 1981a;
11
Kimura, 1981b; Bowen, 1987; Srenk et al., 2010). Este pode ser o único dado retirado de uma EMG em músculos no estadio terminal de doença (contudo, é frequentemente subtil e facilmente ignorado) (LeCouteur e Williams, 2012). A falta de potenciais de inserção é dos dados típicos de uma EMG de lesão muscular isquémica (Srenk et al., 2010).
4.2.2. Potenciais Miniatura de Placa Terminal
Cada célula muscular é inervada por um único ramo do axónio do motoneurónio inferior numa sinapse que se designa “placa terminal” (Steiss, 2003). Na ausência da ativação do motoneurónio inferior, a atividade espontânea no músculo só é gravada em localizações no músculo com elevada concentração de placas terminais (Steiss, 2003; LeCouteur e Williams, 2012). Estas descargas elétricas podem ser gravadas por elétrodos de agulha como ruído de placa terminal, o qual consiste num grande número de potenciais de placa terminal (Steiss, 2003; Srenk et al., 2010).
Trata-se de atividade espontânea de baixa amplitude considerada normal (Bowen, 1974; Kimura, 1981a). Tem na sua origem a libertação intermitente e aleatória de pequenas quantidades de transmissores na junção neuromuscular, resultando em despolarização na membrana pós-sinática (Bowen, 1974; Kimura, 1981a; Cuddon, 2002; LeCouteur e Williams, 2012). Os Potenciais Miniatura de Placa Terminal (PMPT) produzem um som semelhante ao som proveniente de uma concha do mar ou um silvo moderado. Geralmente têm uma amplitude entre os 5 e os 15 µV, embora possam atingir os 50 µV (Bowen, 1974; Kimura, 1981a). Duram 1 a 2 ms e, uma vez que não resultam da completa despolarização da miofibra, apenas são observados se o elétrodo for colocado em estreita proximidade com uma placa terminal.
Doenças que reduzem a libertação de ACh na membrana pré-sinática, como o botulismo, produzem PMPT com amplitude normal mas frequência reduzida. Contrariamente, doenças como miastenia gravis, que diminuem a ligação da ACh pós-sinaticamente, causam uma redução da amplitude com frequência normal (Farnbach, 1980; Cuddon, 2002; Srenk et
al., 2010). A desinervação grave resulta numa ausência completa de PMPT (Farnbach, 1980;
Cuddon, 2002).
O Pico de Placa Terminal (PPT) é uma forma de onda comummente associada aos PMPT, e que resulta da descarga de uma única fibra muscular que é excitada pela atividade nos terminais nervosos (Kimura, 1981a; LeCouteur e Williams, 2012). Estes picos ocorrem de forma intermitente, com um ritmo de disparo de 5 a 50 Hz e uma amplitude que varia entre 100 e 300 µV. Têm um desvio inicial negativo (ascendente) relativamente à linha de base, e são, geralmente, bifásicos (Cuddon, 2002). Ao contrário de outros paradigmas elétricos de gravação, um potencial positivo provoca um desvio descendente no eletromiograma (Sims,
12
1996). A aparência dos PPT assemelha-se à de potenciais de fibrilhação (Cuddon, 2002; LeCouteur e Williams, 2012).
4.2.3. Potenciais da Unidade Motora
Os Potenciais da Unidade Motora (PUM) são observados apenas na presença de contração voluntária ligeira e representam descargas isoladas de uma ou mais unidades motoras (Kimura, 1981a; Bowen, 1987; Cuddon, 2002). Trata-se de um PAC de miofibras na área de gravação do elétrodo (Kimura, 1981a; LeCouteur e Williams, 2012). Estas ondas são, geralmente, bifásicas ou trifásicas, com um pico inicial negativo. As amplitudes são variáveis mas, geralmente, situam-se na ordem dos 100 µV a 3 mV, e disparam a uma taxa de 5 a 7 Hz (Kimura, 1981ª; Bowen, 1987). A sua frequência aumenta com a ativação das mesmas ou novas UM, isto é, com uma maior força de contração (geralmente designada “recrutamento”) (Chrisman et al., 1972; Kimura, 1981a; LeCouteur e Williams, 2012). Com contração muscular máxima, há descarga simultânea de vários PUM, impedindo o reconhecimento de um PUM individual (“padrão de interferência”) (Chrisman et al., 1972; Kimura, 1981a; Bowen, 1987).
Os padrões de PUM, durante a contração máxima, são afetados de diferente forma em diversas doenças (Kimura, 1981a). A miopatia produz um decréscimo nas amplitudes dos PUM, mas com uma densidade normal, enquanto uma desinervação parcial resulta numa densidade reduzida (Bowen, 1987). A presença de vários PUM gigantes (500 µV – 5 mV) pode indicar reinervação (Griffiths e Duncan, 1978; Kimura, 1981a; Bowen, 1987; Childers et al., 1998) e é observada com frequência em algumas neuropatias (Childers et al., 1998; LeCouteur e Williams, 2012). Estes PUM gigantes são, geralmente, polifásicos uma vez que os novos ramos nervosos colaterais têm bainhas de mielina mais finas que as dos axónios estabelecidos e, assim, velocidades de condução menores (Griffiths e Duncan, 1978; Kimura, 1981a; Bowen, 1987).
Como resultado da falta de cooperação por parte dos animais, o estudo dos PUM e padrões de recrutamento não é muito usado em Medicina Veterinária. No entanto, uma área de aplicação consiste na avaliação da gravidade e extensão da lesão em nervos periféricos após traumatismo. A ausência de qualquer PUM induzível num músculo específico, com apoio de peso no membro (músculo extensor) ou flexão (músculo flexor) pode indicar uma lesão nervosa grave (Bowen, 1987; Cuddon, 2002).
4.3. Atividade Espontânea Anormal
A atividade espontânea anormal pode ser observada quando um elétrodo imóvel é posicionado em proximidade a uma miofibra ou grupo de miofibras alteradas (Srenk et al., 2010; LeCouteur e Williams, 2012). A ocorrência de potenciais espontâneos não relacionados
13
com estimulação mecânica não deve ser observada no músculo normal. Estes potenciais podem ser gerados por fibras individuais (potenciais de fibrilhação - PF - e ondas agudas positivas - OAP) ou pelas fibras de uma UM (potenciais de fasciculação) (LeCouteur, 2005). Recorre-se a um sistema de classificação estandardizado para caraterizar a atividade espontânea em cada músculo testado. A atividade muscular é classificada de normal (0) a grave (+4) (LeCouteur e Williams, 2012).
Os PF e as OAP são tipicamente observados em miopatias ou neuropatias com desinervação. No entanto, estes tipos de onda são também relatados em indivíduos normais, especialmente nos músculos da região paraespinhal lombossagrada (Date et al., 1996; Nardin
et al., 1998) e cervical (Date et al., 2006). Deve, contudo, considerar-se a possibilidade de
ocorrência de PPT de aparência atípica que imitam a configuração dos PF e OAP (Dumitru, 2000; Dumitru et al., 2001). Uma taxa de disparo mais lenta e descargas irregulares são caraterísticas que permitem distinguir PPT atípicos de potenciais espontâneos anormais. Apesar disto, confirmou-se que podem estar presentes potenciais de desinervação em indivíduos normais, mesmo após examinar cuidadosamente os tipos de onda, por forma a excluir PPT (Date et al., 2006). Pensa-se que estes potenciais espontâneos sejam causados por herniação de discos ou estiramento e aprisionamento dos ramos primários posteriores por movimento excessivo (Haig, 2002). Não é incomum observar atividade espontânea ligeira nos músculos das extremidades distais em cães saudáveis (Cork et al., 1983; Chrisman et al., 1984).
Acredita-se que a atividade espontânea anormal após a lesão surge na EMG a tempos específicos, os quais seguem a regra do envolvimento muscular dependente do comprimento do nervo em questão. Alguns autores alertaram para o facto da duração dos sinais clínicos não parecer estar relacionada com a atividade espontânea (Dillingham et al., 1998a; Dillingham et
al., 1998b; Pezzin et al., 1999; Dillingham et al., 2000). Embora estes autores aconselhem
precaução ao interpretar resultados de exames de eletrodiagnóstico com base na duração dos sinais clínicos, a evolução dos dados de EMG pode ser relacionada com o tempo passado após a lesão e a gravidade do dano axonal (Date e Kim, 2006). A frequência destes potenciais está, geralmente, relacionada com o número de neurónios motores afetados (Thieman et al., 2010).
O facto de não se detetarem alterações na EMG não significa necessariamente que não exista doença. Algumas miopatias (p.e, dermatomiosite) podem apresentar uma distribuição desigual. Para além disso, doenças nas quais a desmielinização é a lesão inicial (p.e, a doença de Niemann-Pick em gatos) podem originar um eletromiograma normal em estadios iniciais e até que se desenvolva uma axonopatia secundária. É necessário algum tempo (dias a semanas) para que a atividade espontânea seja detetável num músculo que sofreu uma lesão
14
aguda dos seus neurónios motores. Este atraso varia com a proximidade da lesão ao local de gravação. A EMG é também incapaz de detetar disfunções restritas apenas a neurónios sensoriais (LeCouteur e Williams, 2012).
4.3.1. Fasciculação
A contração espontânea que ocorre quando UM ou partes das UM vizinhas descarregam denomina-se “fasciculação”. São provavelmente causadas pela ativação enfática de fibras musculares como resultado da descarga em pacemaker das fibras do nervo ou músculo. Por vezes podem observar-se fasciculações superficiais através da pele (Steiss, 2003), a olho nu, ao contrário dos PF. As fasciculações profundas são mais comuns e são detetadas apenas por EMG (Kimura, 2001; Cuddon, 2002). Em termos elétricos, as fasciculações têm durações, amplitudes e outras caraterísticas similares aos PUM (Steiss, 2003). Produzem um som agudo sem qualquer relação com o esforço (Kimura, 2001) e podem surgir numa variedade de doenças que afetam os nervos e o músculo e, menos frequentemente, em doenças degenerativas da substância cinzenta da ME (Kimura, 2001; Steiss, 2003).
4.3.2. Potenciais de Fibrilhação
Potenciais de Fibrilhação (PF) são potenciais espontâneos bi ou trifásicos que representam a descarga de fibras musculares individuais (Sims e Redding, 1978; Cuddon, 2002), como resultado de uma desestabilização do seu sarcolema (Kimura, 2001). Apresentam um desvio inicial da linha de base, geralmente, no sentido positivo, exceto se forem gravados numa região de placa terminal (Cuddon, 2002). Também denominados “potenciais de desinervação”, são uma das principais caraterísticas elétricas do músculo desinervado, sendo observados comummente em alterações neurogénicas e musculares primárias (Sims, 1996).
O seu som a partir de um altifalante é semelhante a “fritar ovos”, “chuva a cair num telhado de zinco” ou “amarrotar de papel” (Cuddon, 2002; Steiss, 2003). A amplitude dos PF varia de 50-350 µV com durações de 1 - 2 ms. Os PF não ocorrem imediatamente após a desinervação. Surgem no músculo desinervado após um período latente que é proporcional ao comprimento do axónio restante, distalmente ao local da lesão no nervo (Sims e Redding, 1978; Steiss, 2003). O seu início pode ser precedido por um aumento na atividade de inserção. Após o aparecimento destes potenciais no músculo desinervado, a sua taxa de ocorrência aumenta ao longo de um período de várias semanas, e eles persistem até que o músculo seja reinervado ou até que não existam mais fibras musculares viáveis (Sims, 1996; Steiss, 2003).
Os PF também são vistos em disfunções miopáticas, como distrofias musculares, polimiosite e dermatomiosite. A sua origem está relacionada com alterações da membrana
15
oscilante ou pré-potenciais irregulares causados por instabilidade na membrana (Sims, 1996; Cuddon, 2002; Steiss, 2003). Em neuropatias caraterizadas pela desmielinização em vez de degenerescência axonal (degenerescência Walleriana), os PF tendem a estar ausentes (Sims, 1996; Steiss, 2003).
4.3.3. Ondas Agudas Positivas
Ondas Agudas Positivas (OAP) são potenciais espontâneos anormais que surgem no músculo desinervado e numa variedade de alterações miopáticas (Sims, 1996). Caraterizam-se por uma fase inicial positiva seguida por uma fase gradualmente negativa (Sims, 1996; Steiss, 2003). Estas ondas apresentam amplitudes variáveis, entre 50 µV e 4 mV (Cuddon, 2002) e, embora produzam um som menos agudo que os PF e, geralmente tenham uma menor taxa de descarga, considera-se atualmente que representam um tipo de PF (Steiss, 2003). Representam também membranas de miofibras irritadas, apesar de, ao contrário dos PF, o potencial parar numa área imediatamente adjacente ao elétrodo de gravação (Cuddon, 2002).
Embora, na maioria dos casos as OAP e os PF tenham o mesmo significado, há situações em que não são vistos em conjunto (Kraft, 1996). As OAP podem ser gravadas mais precocemente que os PF após uma lesão no nervo periférico que cause axonotemese (Sims, 1996; Kraft, 1996).
4.3.4. Descargas Repetitivas Complexas
As Descargas Repetitivas Complexas (DRC), anteriormente denominadas “descargas bizarras de alta frequência” ou “descargas pseudomiotónicas”, consistem em potenciais polifásicos que descarregam espontaneamente a alta frequência. Dentro do comboio das descargas, cada potencial apresenta a mesma morfologia (Sims, 1996; Steiss, 2003; LeCouteur e Williams, 2012). Apresentam frequência, forma e amplitude uniformes e representam a descarga de múltiplas miofibras em próxima sincronia (Farnbach, 1980; Kimura, 1981b; Bowen, 1987). Este comportamento sugere a presença de fibras pacemaker que oscilam (Sims, 1996; Steiss, 2003). O início é frequentemente associado ao movimento da agulha e as descargas começam e param abruptamente (Farnbach, 1980; Kimura, 1981b; Bowen, 1987; Sims, 1996), mas podem estar associadas a outros estímulos mecânicos como a percussão do músculo (LeCouteur e Williams, 2012). A sua gama de amplitudes situa-se, geralmente, entre 100 µV e 1 mV (Farnbach, 1980; Kimura, 1981b; Bowen, 1987). Estes potenciais podem ocorrer numa variedade de alterações neuromusculares e sugerem a presença de uma condição crónica, como hiperadrenocorticismo (Farnbach, 1980; Sims, 1996) ou polimiosite (Sims, 1996).
16
As DRC são frequentemente confundidas com Potenciais Miotónicos (PM) (Cuddon, 2002). Embora, a certa altura, se designassem “potenciais pseudomiotónicos”, estes potenciais não aumentam e diminuem em amplitude e frequência como os verdadeiros PM (Sims, 1996; Steiss, 2003). A partir do altifalante do eletromiógrafo, estas descargas contínuas têm sons agudos como um motor (Steiss, 2003) ou uma espingarda (Kimura, 2001).
4.3.5. Potenciais Miotónicos
As descargas miotónicas podem ocorrer como resultado de uma permeabilidade anormal nas membranas das fibras musculares. Os músculos continuam a ser eletricamente ativos, mesmo depois do fim da contração voluntária (Sims, 1996; Steiss, 2003). Trata-se de descargas repetitivas e independentes de miofibras simples lesionadas, que ocorrem a uma taxa situada entre os 20 e os 80 Hz (Kimura, 1981b; van Nes, 1986). É possível observar 2 tipos de onda: uma série de ondas positivas idênticas a OAP, ou vários potenciais bifásicos compostos por um pico inicial positivo e pequeno, seguido por um pico maior negativo, assemelhando-se a PF (Kimura, 1981b). Estes potenciais de alta frequência (100 a 200/s) aumentam e diminuem espontaneamente em amplitude e taxa, num padrão de EMG que se tornou a assinatura elétrica da miotonia congénita (Farnbach, 1980; Kimura, 1981b; Sims, 1996). No entanto, e apesar do seu nome, não são patognomónicos de miotonia, podendo ser observados em radiculopatias e polineuropatias (LeCouteur e Williams, 2012).
Para além da atividade nervosa pré-sinática, o início pode ser precipitado mecanicamente através da percussão do músculo ou movimento da agulha de EMG. A atividade espontânea pode durar 1 s ou mais. Nalguns tipos de miotonia, as descargas repetitivas podem ser explicadas por uma alteração na condução do cloro nas membranas musculares, enquanto noutros tipos a doença pode estar relacionada com uma alteração na condução do Na+ (Steiss, 2003). A monitorização áudio dos PM revela um som de EMG caraterístico, conhecido como “mergulho de bombardeiro”, por se assemelhar ao som do mergulho de aviões a hélice (Cuddon, 2002; LeCouteur e Williams, 2012).
4.4. Atividade Evocada no Músculo
A atividade muscular esquelética é geralmente evocada por estimulação elétrica dos nervos motores, com agulhas intramusculares ou elétrodos de superfície usados como elétrodos de gravação. Noutras respostas evocadas, os recetores são estimulados fisiologicamente e os músculos são ativados de forma reflexa (Sims, 1983; Steiss, 2003). A atividade muscular pode também ser produzida por estimulação elétrica transcranial ou estimulação eletromagnética do córtex motor (Sims, 1996).
17 4.4.1. Atividade Evocada Eletricamente
Independentemente do componente do SNP em estudo, existem vários princípios gerais que devem ser considerados ao induzir atividade em procedimentos de eletrodiagnóstico. Em primeiro lugar, os elétrodos de estimulação devem estar sempre posicionados de forma que o cátodo (polo negativo) esteja mais próximo do local de gravação que o ânodo (polo positivo), evitando assim o bloqueio de condução anodal do impulso propagado (Farnbach, 1980b; Kimura, 1981c). Conforme a corrente flui entre o ânodo e o cátodo, são as cargas negativas por baixo do cátodo as responsáveis pela despolarização nervosa. A carga positiva por baixo do ânodo induz hiperpolarização (Kimura, 1981c). O cálculo preciso da velocidade de condução nervosa (VCN) depende de uma medição adequada da distância entre pontos catódicos consecutivos (Kimura, 1981c; Bowen, 1987).
Durante a estimulação, o pulso de voltagem inicial do estimulador cria um artefacto de estímulo, um potencial de campo distante que obscurece a porção inicial de qualquer forma de onda registada (Dumitru et al., 2001). Este artefacto pode dificultar a interpretação dos potenciais quando os elétrodos de estimulação e gravação se encontram demasiado próximos (Koo et al., 2012).
Uma vez que a estimulação com corrente alta ou duração prolongada produz um artefacto de estímulo mais elevado, a força da corrente aplicada e a sua duração devem ser suficientes para obter uma resposta supramaximal (Farnbach, 1980b; Kimura, 1981c; Bowen, 1987; Koo et al., 2012). Uma estimulação supramaximal (50% superior ao estímulo necessário para ativar todos os axónios) garante que todo o conjunto de axónios de um determinado nervo é avaliado independentemente do seu diâmetro e relação espacial com os elétrodos de estimulação (Farnbach, 1980b; Kimura, 1981c; Bowen, 1987). Para calcular o valor de estimulação determina-se o estímulo máximo, i.e, o valor de corrente para o qual a amplitude da onda M é mais elevada e incrementos adicionais não têm efeito, e multiplica-se esse valor por 1,5. Este processo é repetido noutros locais de estimulação, contudo, os elétrodos de gravação não devem ser deslocados após a determinação da sua localização ideal (LeCouteur e Williams, 2012). A colocação do elétrodo-terra entre os elétrodos de estimulação e de gravação é também importante, de forma a diminuir os artefactos associados ao estímulo (Cuddon, 2002; Koo et al., 2012).
4.4.2. Atividade Evocada por Estimulação Magnética
A estimulação magnética apresenta diversas vantagens relativamente à estimulação elétrica: é minimamente invasiva, quase indolor e uma técnica segura que exige menos tempo da parte do operador (Barker et al., 1987, Evans 1991, Barker 1999). Pode ser realizada apenas sob sedação ou com um plano anestético leve (Barker et al., 1987). A técnica de
18
estimulação magnética de nervos periféricos tem-se mostrado útil em Medicina Veterinária (Van Soens et al., 2008; Van Soens et al., 2009). Está descrita em cães e gatos com avulsão do plexo braquial (Van Soens et al., 2009) e disfunção unilateral do nervo ciático, podendo ser usada como uma ferramenta adicional de diagnóstico e prognóstico (Van Soens et al., 2010).
Dependendo do tipo de lesão nervosa, podem observar-se alterações em diferentes parâmetros dos potenciais musculares evocados (PME). De forma semelhante ao que acontece com os potenciais evocados eletricamente, a amplitude do PME reflete o número e tamanho das UM que inervam o músculo. As alterações da amplitude podem ter origem na perda axonal, bloqueio de condução distalmente ao local de estimulação após desmielinização aguda, corte de axónios ou ambos. A latência é um reflexo da condução ao longo do axónio e as alterações resultam da perda dos axónios mais rápidos, bloqueio da condução destes axónios, ou alterações da bainha de mielina (Welch, 1996; Cuddon, 2002). Em casos graves, a estimulação pode mesmo não originar qualquer tipo de potencial (Welch, 1996; Van Soens et
al., 2009).
4.4.3. Onda M
Quando o nervo motor de um músculo é estimulado de forma supramaximal com corrente elétrica, as UM são ativadas assim que as suas fibras nervosas atingem o limiar (Sims, 1996; Steiss, 2003). Um PA numa fibra muscular precede um aumento da tensão, e quando várias fibras estão ativas ao mesmo tempo, um potencial muscular de ação composto (PMAC) pode ser gravado (Sims, 1996).
Este potencial é também designado como resposta muscular, resposta M ou onda M (Sims, 1996; Steiss, 2003). É o resultado da propagação ortodrómica de PA ao longo do nervo, libertação de ACh na junção neuromuscular e despolarização da miofibra (LeCouteur e Williams, 2012). Trata-se de um potencial bi ou trifásico cuja amplitude, dependendo do músculo específico e do tipo de elétrodos de gravação, pode variar desde poucas a várias centenas de mV, e é proporcional ao número e tamanho das fibras a disparar. A latência da onda M é proporcional à distância entre o músculo e a localização do elétrodo de estimulação no nervo periférico. A duração é um reflexo de sincronia, isto é, a que distância os PA permanecem no nervo e a que proximidade as fibras musculares descarregam no tempo (Sims, 1996; Steiss, 2003).
As ondas M que resultam da estimulação em cada local devem apresentar uma configuração semelhante. Se isto não se verificar, é possível que haja envolvimento de outro nervo. Este problema é frequente quando se realiza estimulação na área da anca, onde os segmentos peroneal e tibial do nervo ciático são difíceis de isolar (LeCouteur e Williams, 2012).
19
Para qualquer nervo, quanto mais afastado do músculo o nervo motor é estimulado, maior a duração da onda M e menor a sua amplitude (Steiss, 2003). Alterações no nervo periférico e junção neuromuscular geralmente causam diminuição da amplitude da onda M e aumento da sua duração (Sims, 1996; Cuddon, 1998). A diminuição da amplitude da onda M não é específica, podendo ser observada em neuropatias ou miopatias que afetem o músculo avaliado. Para além disso, a perda de alguns entrenós de mielina consecutivos (que podem ser apenas 2), resulta no bloqueio da condução no neurónio, pelo que a diminuição da amplitude pode também ser vista com desmielinização. O fenómeno de bloqueio da condução pode ser observado em situações tão diversas como lesões metabólicas de um nervo com preservação de axónios (neuropraxia), ou previamente à ocorrência completa de degenerescência Walleriana num axónio recentemente lesionado (axonotemese). Isto torna-se evidente na comparação entre ondas M de diferentes locais de estimulação. Em locais mais distais podem obter-se respostas aparentemente normais que mudam (com frequência de forma dramática) após estimulação em locais proximais (LeCouteur e Williams, 2012).
4.4.4. Estimulação Nervosa Repetitiva
Se um nervo periférico é estimulado repetidamente, o PMAC resultante irá manter a sua amplitude desde que a taxa de estimulação não seja muito alta (Sims, 1996; Cuddon, 1998). A “estimulação nervosa repetitiva supramaximal” é o procedimento eletrofisiológico mais usado para avaliação das disfunções da junção neuromuscular (Keesey, 1989; Kimura, 1989c). O exame é realizado através da estimulação de um nervo periférico com uma série de impulsos supramaximais (Cuddon et al., 1999; LeCouteur e Williams, 2012) com avaliação simultânea dos PMAC evocados. São empregues taxas de estimulação de 1, 3, 5 e 10 Hz (Sims e Selcer, 1981), mas podem variar de 0,5 a 50 Hz (LeCouteur e Williams, 2012). Deve ter-se cuidado na aplicação desta técnica, uma vez que taxas de estimulação superiores a 5 Hz podem resultar em falsos positivos em animais normais (Jeffery, 2004). É importante deixar, pelo menos, 1 min de tempo de recuperação entre estimulações durante estes testes de eletrodiagnóstico (LeCouteur e Williams, 2012).
As alterações mais distintas na amplitude e área dos PMAC são, usualmente, detetadas por comparação do primeiro potencial com as ondas subsequentes. As amplitudes e áreas destes últimos potenciais são medidas e expressas como um decréscimo, em percentagem, em relação ao primeiro PMAC (Sims e Selcer, 1981; LeCouteur e Williams, 2012).
Na determinação de valores de referência normais de padrões decrementais em cães adultos, estímulos de baixas frequências de 3 Hz ou menos não resultaram em qualquer decréscimo acima dos 10% durante a estimulação (Sims e Selcer, 1981; Malik e Ho, 1991; Waxenberger et al., 1992). Ocorrem, no entanto, aumentos significativos no decréscimo