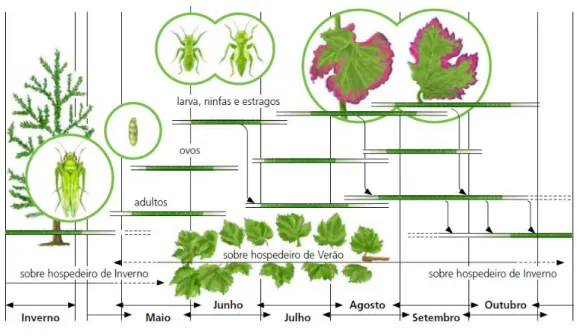
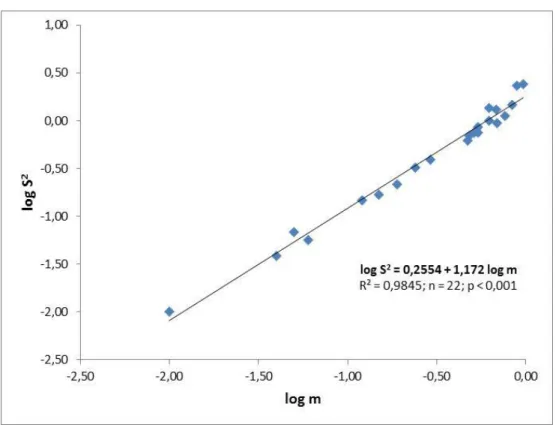
Elaboração de um plano de amostragem para Empoasca vitis Goethe (Homóptera; Cicadellidae) em vinha na sub-região do Lima da Região demarcada dos Vinhos Verdes
Texto
Imagem







Documentos relacionados
nesta nossa modesta obra O sonho e os sonhos analisa- mos o sono e sua importância para o corpo e sobretudo para a alma que, nas horas de repouso da matéria, liberta-se parcialmente
Tabela 1: Unidades de Registo Codificadas na Categoria Nível de Stress Percebido…….25 Tabela 2: Unidades de Registo Codificadas na Categoria Fatores de Stress…………...…27
A hipertensão arterial é um importante factor de risco para várias doenças cardiovasculares, nomeadamente para o acidente vascular cerebral, enfarte agudo do miocárdio
O plano de “culturalização” da marca junto à equipe de vendas é feito inicialmente através da seleção dos colaboradores, que está detalhadamente especificada no Manual
62 uso de uma via metabólica alternativa por Biomphalaria tenagophila e B. glabrata expostas a 20, 40, 60, 80 e 100 mg/L de CaCO
age o& Va&òaZuÁ convexuò and PO&ÁOIUA tati^fwnó ^emaleA, Qiving fieAolÁM compa/iablz to otheA òtudieA and indlo.atA.nQ tke, vahxe, o{ tkeAe... One egg is produced and
tive outros problemas a não ser com a droga). A nossa perspectiva teórica valoriza os factores sociais, os quais desempenham um papel extremamente importante no processo de
