Protocolos de sincronização de cio em
bovinos
Avaliação da resposta a um esquema Ovsynch modificado
em vacas de leite
Lúcia Susana da Costa Marciel
Orientador: Professora Doutora Rita Payan Carreira Co-orientador: Dr. José Pedro Azevedo
UNIVERSIDADE DE TRÁS-OS-MONTES E ALTO DOURO VILA REAL, 2010
Ciências Veterinárias
Protocolos de sincronização de cio em
bovinos
Avaliação da resposta a um esquema Ovsynch modificado
em vacas de leite
Lúcia Susana da Costa Marciel
Orientador: Professora Doutora Rita Payan Carreira Co-orientador: Dr. José Pedro Azevedo
UNIVERSIDADE DE TRÁS-OS-MONTES E ALTO DOURO VILA REAL, 2010
ii
“Não é o desafio que define quem somos nem o que
somos capazes de fazer. O que nos define é o modo
como enfrentamos esse desafio.”
iii Agradecimentos
Ao terminar este trabalho desejo agradecer a todas as pessoas que directa ou indirectamente contribuíram para a sua realização, em especial:
À Professora Doutora Rita Payan Carreira por ter aceite ser minha orientadora e pelos ensinamentos, conselhos e criticas ao longo do trabalho.
Ao Dr. José Pedro Azevedo por ter orientado o meu estágio e por tudo o que me ensinou e pelo apoio que demonstrou ao longo de todo este percurso.
Ao professor Doutor Jorge Colaço pela disponibilidade e ajuda que me prestou no decorrer deste trabalho.
À Intervet Shering na pessoa do Director da área dos bovinos, Dr. Tiago Teixeira pela cedência da hormona hCG que permitiu a realização deste trabalho e pela sua disponibilidade e conselhos prestados.
Aos produtores, Sr. Manuel Balazeiro e D. Almerinda por terem permitido a realização deste estudo nas suas explorações e por toda a disponibilidade e amabilidade com que me trataram.
A todos os meus amigos que me apoiaram e estiveram presentes durante estes anos, em especial à Amélia, à Ana, ao Diogo e ao Frostes.
Aos meus sobrinhos simplesmente por existirem e me fazerem sorrir e viver a vida com mais alegria.
À minha irmã por ser o meu ombro amigo.
À minha mãe por seu o meu pilar, por me incentivar e fazer-me acreditar.
Ao meu pai, por tudo aquilo que foi e continua a ser para mim, apesar de já não se encontrar entre nós; continua a dar-me força e motivação para não desistir dos meus sonhos.
iv Resumo
O maneio reprodutivo é essencial para optimizar a performance reprodutiva nos actuais sistemas de produção, e a fraca detecção de cio é um dos principais factores limitantes do sucesso das metas reprodutivas propostas para a maioria das explorações leiteiras. Com vista a melhorar a taxa de concepção das explorações, têm-se procurado optimizar vários protocolos de sincronização de estro e/ou de ovulação, que geralmente apresentam pequenas diferenças consoante são aplicados a bovinos de leite ou de carne. Os mais frequentemente utilizados são os protocolos que utilizam as prostaglandinas, associadas ou não a agonistas de GnRH. Existem protocolos de sincronização da ovulação desenhados para permitir a inseminação sem detecção do cio, já que esta última é um dos principais factores de insucesso da IA nas explorações de leite. Estes protocolos controlam o desenvolvimento da onda folicular e a ovulação, permitindo a inseminação das fêmeas num momento fixo e pré-determinado, procurando facilitar o maneio, conseguir um aumento da taxa de submissão até determinado período pós-parto sem baixar a taxa média de gestação da exploração e diminuir o número de vacas a serem refugadas por ineficiência reprodutiva. Neste trabalho procurou-se avaliar o resultado da introdução de um protocolo modificado de sincronização da ovulação em duas explorações através do estudo das taxas de gestação da exploração, estabelecendo-se a comparação entre este tratamento (OvSynch modificado), o protocolo OvSynch tradicional e um grupo controlo de fêmeas. Em concreto, o protocolo modificado consistiu na administração de PGF2α ao dia 8 após a primeira administração de GnRH, e diferiu do OvSynch tradicional no
facto de a segunda dose do análogo da GnRH (5 ml de Receptal ®) ter sido substituída por hCG (1500 UI de Chorulon®).
Em ambas as explorações estudadas, os animais tratados de acordo com o protocolo modificado apresentaram taxas de gestação inferiores tanto às dos grupos controlo como à do grupo tratado com OvSynch tradicional, sendo estas diferenças mais pronunciadas na exploração B. Nesta exploração em particular, o grupo tratado com OvSynch tradicional apresentou uma taxa de gestação superior quer ao grupo controlo quer ao tratado com o protocolo OvSynch modificado (77,78%, 69,77% e 20%, respectivamente). Na exploração A, as principais diferenças foram encontradas entre o grupo tratado com o protocolo modificado e os restantes grupos (25%, 50% e 62,96%, respectivamente para os protocolos OvSynch modificado e tradicional e para o grupo controlo), que não diferiam estatisticamente entre si.
No trabalho agora apresentado não foram encontrados dados favoráveis à utilização do hCG no esquema utilizado do protocolo OvSynch modificado.
v Abstract
In order to improve farm conception rate, many efforts have been made to improve different oestrus and/or ovulation synchronisation protocols that have some small differences when applied to dairy or beef cattle. The most frequently used in practice are those including prostaglandins associated or not to GnRH agonists. There are protocols for ovulation synchronisation that allow farmers to inseminate without heat detection, since this issue is one of the major factors for unsuccessful AI in dairy farms. By controlling the follicular wave and ovulation these protocols allow cow insemination in a fixed and pre-determined moment, turning easier the farm management, raising the submission rate up to a certain period after birth, without decreasing the farm pregnancy rate and reducing the number of cows culled due to reproductive inefficiency.
This study aimed to evaluate the response to a modified ovulation synchronization protocol in two dairy farms, through the study of the pregnancy rates of the farms, establishing a comparison between this treatment (modified OvSynch), the traditional OvSynch protocol and a control group within each farm. Specifically, the new protocol consisted on the administration of PGF2α at day 8, after the first GnRH dosis and on the replacement of the second dose of GnRH
analogue (5 ml Receptal ®) by hCG (1500 IU Chorulon®). In both farms, animals treated according to the modified protocol showed lower pregnancy rates than the two other groups. These differences were more pronounced at farm B, where higher pregnancy rates were observed for the traditional OvSynch group in comparison to controls and the modified Ovsynch group (77.78%, 69.77% and 20% respectively). In farm A, the main differences were found between the group treated with the modified protocol and the other groups (25%, 50% and 62.96% respectively for the modified and the traditional Ovsynch protocol and for the control group).
Our study fail to demonstrate any advantage in replacing the second GnRH agonist administration by hCG.
vi Índice Geral
Resumo ... iv
Abstract ... v
Índice de Figuras ... viii
Índice de Tabelas ... viii
Lista de Siglas/Acrónimos, Abreviaturas e Sinais/Símbolos ... ix
I. Introdução ... 11
1. Fisiologia reprodutiva das vacas ... 11
2. Dinâmica folicular ... 13
3. Luteogénese e luteólise ... 16
4. Maneio reprodutivo na exploração: a detecção de cio ... 18
5. Momento da inseminação ... 22
6. Factores que contribuem para a diminuição da taxa de concepção ... 23
7. Métodos de controlo do estro e/ou ovulação ... 26
7.1 Protocolos de sincronização do estro ... 27
7.1.1. Indução do estro com prostaglandinas ... 27
7.1.2. Controlo da onda folicular e indução do estro com PGF2α (Selectsynch) ... 29
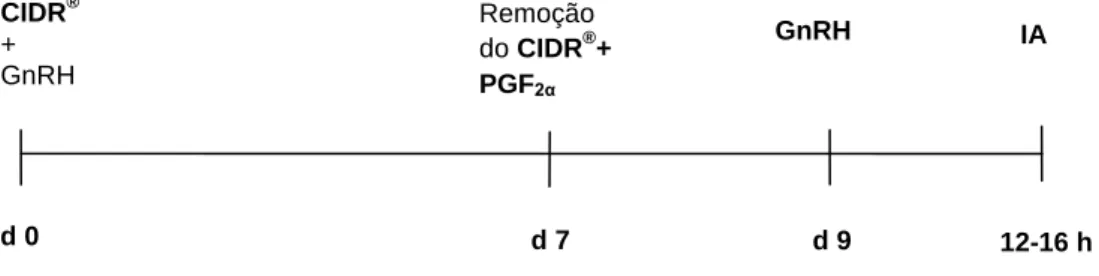
7.1.3. Sincronização da ovulação com progesterona e prostaglandinas ... 31
7.2. Protocolos de sincronização da ovulação ... 32
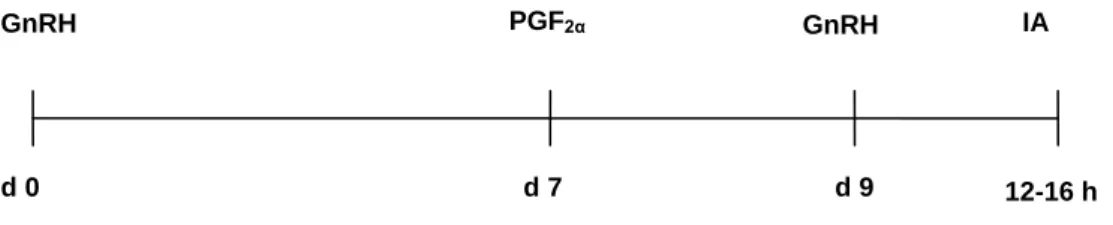
7.2.1. Protocolo Ovsynch ... 32
7.2.2. Modificações ao protocolo Ovsynch ... 34
7.2.2.1. Pré sincronização antes da IATF (Presynch) ... 34
7.2.2.2. Co-sincronização (Cosynch) ... 36
7.2.2.3. Protocolo Ovsynch de 56 horas ... 37
7.2.2.4. Protocolo Ovsynch associado à progesterona ... 37
7.2.2.5. Protocolo Heatsynch ... 38
vii
II. Avaliação da resposta a um esquema Ovsynch modificado em vacas de leite na região
de entre o Douro e Minho de Portugal ... 40
1. Introdução ... 41
2. Material e métodos ... 42
2.1. Caracterização das explorações ... 42
2.2. Tratamentos ... 44
2.3. Animais ... 45
2.4. Registo de dados e análise estatística... 46
3. Resultados ... 47
4. Discussão e conclusão ... 53
viii Índice de Figuras
Figura 1. Representação esquemática do ciclo éstrico da vaca, para uma duração média de ciclo de 21
dias ... 12
Figura 2. Aumento e declínio das hormonas reprodutivas durante o ciclo éstrico ... 18
Figura 3. Protocolos baseados no uso de PGF2α para sincronização do estro em bovinos ... 29
Figura 4. Representação esquemática do protocolo Selectsynch ... 30
Figura 5. Representação esquemática de um protocolo de sincronização à base de progesterona ... 32
Figura 6. Representação esquemática do protocolo Ovsynch ... 33
Figura 7. Representação esquemática do protocolo Ovsynch modificado com inclusão de um esquema de pré-sincronização ... 35
Figura 8. Representação esquemática do protocolo Cosynch ... 36
Figura 9. Representação esquemática do protocolo Ovsynch de 56 horas ... 37
Figura 10. Representação esquemática do protocolo Ovsynch associado à progesterona ... 38
Figura 11. Representação esquemática do protocolo hCG-synch ... 39
Figura 12. Esquema do protocolo Ovsynch implementado nas duas explorações ... 44
Figura 13. Esquema do protocolo Ovsynch modificado utilizado ... 45
Figura 14. Representação gráfica da taxa de gestação do grupo controlo, grupo 1 e grupo 2 da exploração A e B ... 52
Índice de Quadros Quadro1. Sinais primários e secundários de cio ... 21
Quadro 2. Caracterização geral das explorações A e B no que respeita à produção média de leite aos 305 dias no ano 2009, e alguns dos parâmetros reprodutivos utilizados na sua selecção ... 42
Quadro 3. Caracterização geral dos grupos controlo, 1 e 2 da exploração A e B quanto à idade, produção de leite e número de partos ... 47
Quadro 4. Parâmetros reprodutivos relativos ao parto anterior à introdução no estudo para o grupo controlo, e para os grupos 1 e 2 para ambas as explorações ... 48
Quadro 5. Número de IA antes de implementados os protocolos Ovsynch e Ovsynch modificado ... 49
Quadro 6. Parâmetros reprodutivos relativos aos grupos controlo, e grupos 1 e 2 para cada uma das explorações ... 50
Quadro 7. Taxas de gestação relativas à exploração e aos grupos controlo, 1 e 2 para as explorações A e B ... 50
Quadro 8. Percentagens para o global dos diagnósticos de gestação, incluindo diagnósticos de gestação por exame ginecológico ou através do retorno ao cio, das explorações A e B ... 51
ix
Listas de Siglas/Acrónimos, Abreviaturas e Sinais/ Símbolos
a – Ano
AM/PM - Manhã/tarde
ANOVA - Análise de variância BEN - Balanço energético negativo BRSV - Vírus sincicial respiratório bovino BVD - Vírus da diarreia bovina
CIDR - Controlled intravaginal drug release CL - Corpo lúteo
d-Dia
dpp - Dias pós parto EB - Benzoato de estradiol ECP - Cipionato de estradiol
FSH - Hormona folículo-estimulante g - Grama
GnRH - Hormona libertadora de gonadotropina h - Hora
hCG - Gonadotropina coriónica humana IA - Inseminação artificial
IA/C - Número de inseminações por concepção IATF - Inseminação artificial a tempo fixo IBR - Rinotraqueíte infecciosa bovina IGF-I - Insulin-like growth factor I IM - Intramuscular
IEP - Intervalo entre partos kg - Quilograma
kg/305d - Produção de leite em quilos aos 305 dias LH - Hormona luteinizante m - Mês m2 - Metro quadrado Máx - Máximo Mín - Mínimo ml - Mililitro n - Amostra (nº animais) Nº - Número
x
NºIA/fecundação - Número de inseminações artificiais por fecundação OT - Oxitocina
p - Valor de p
PGE2 - Prostaglandina E2
PGF2α - Prostaglandina F2α
P-IAF - Intervalo entre o parto e a inseminação artificial fecundante PRID - Progesterone release intravaginal device
PVE - Período voluntário de espera
P-1ªIA - Intervalo entre o parto e a primeira inseminação artificial UI - Unidade internacional vs - Versus χ2 -Qui-quadrado μg - Micrograma ® - Marca registada % - Percentagem 1ª - Primeira 2ª - Segunda < - Menor > - Maior = - Igual
11 I. Introdução
Nos últimos anos tem-se vindo a observar o decréscimo progressivo dos índices reprodutivos em explorações intensivas de bovinos de leite, em particular da taxa de concepção, que se reflecte num acréscimo do intervalo entre partos (Lucy et al., 2004). Esta redução nos índices reprodutivos da exploração decorre de uma espiral de efeitos que engloba não apenas factores biológicos próprios ao animal, de que são exemplos o balanço energético negativo, o nível nutricional, o arrastar do processo de involução uterina e a quantidade de leite produzido, mas ainda factores ambientais (conforto e bem-estar na exploração) e humanos, como sejam o maneio nutricional e a detecção de cios (Bello e Pursley, 2006).
Na tentativa de dar resposta às preocupações conjuntas dos proprietários e dos veterinários, e em simultâneo maximizar as probabilidades de uma fêmea ficar gestante com um número reduzido de IA, foram-se desenvolvendo vários protocolos de sincronização de cios, alguns dos quais independentes da detecção de cios. Na maior parte destes protocolos, a manipulação da(s) onda(s) de desenvolvimento folicular, associada ou não ao encurtamento da fase lútea, procuram ainda encurtar o número de dias em aberto, indirectamente visando o encurtamento do intervalo entre partos (Lucy et al, 2004; Cavalieri et al., 2006; Lane et al., 2008).
Neste trabalho procuraremos fazer uma revisão geral do ciclo reprodutivo da vaca, com realce dos fenómenos de desenvolvimento folicular cíclico, para depois abordar os protocolos desenvolvidos para manipular o ciclo éstrico em bovinos. Estes dados ser-nos-ão úteis para, na segunda parte, discutir os resultados obtidos na implementação de um protocolo Ovsynch modificado em duas explorações na região entre Douro e Minho de Portugal.
1. Fisiologia reprodutiva das vacas
As vacas são uma espécie poliéstrica contínua. Uma vez atingida a puberdade, os ciclos éstricos sucedem-se ao longo do ano até ser interrompido pela gestação (Ball e Peters, 2004)
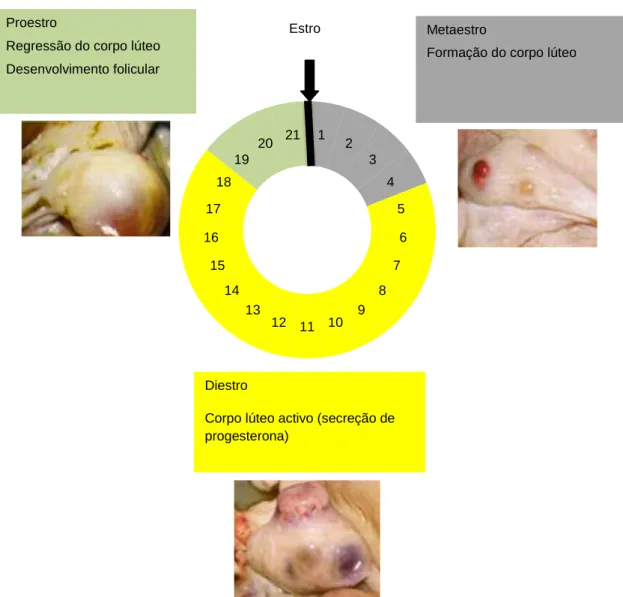
O ciclo éstrico dos bovinos tem uma duração média de 3 semanas, mas pode variar entre os 17 e os 25 dias (Stevenson, 2007). De uma maneira geral, a duração do ciclo éstrico é 1 a 2 dias mais curto nas novilhas do que nas vacas (Stevenson, 2007). Classicamente, o ciclo éstrico divide-se em quatro fases (Ball e Peters, 2004) (Figura 1):
- estro (dia 0); - metaestro (dia 1-4);
12 - diestro (dia 5-18);
-proestro (dia 19 até o início do estro);
Figura 1 – Representação esquemática do ciclo éstrico da vaca, para uma duração média de ciclo de 21 dias. (Adaptado de Sánchez, 2008)
O estro caracteriza-se pela receptividade sexual da fêmea e é nesta fase que ocorre o crescimento folicular rápido, de forma a permitir a posterior ovulação do folículo. O metaestro, do dia 1 ao dia 3-4, caracteriza-se pela maturação final do folículo, pela ovulação e formação do corpo hemorrágico, ainda com fraca capacidade de produzir progesterona, e que evoluirá para corpo lúteo. O diestro, do dia 4-5 ao dia 18, inicia-se quando a progesterona atinge concentrações significativas e continua até à regressão do corpo lúteo (CL). Nesta altura as concentrações sanguíneas de progesterona diminuem rapidamente e inicia-se o proestro, do dia 18-19 até ao início do estro no dia 20-21 (Ball e Peters, 2004; Stevenson, 2007), fase em
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 Estro Metaestro
Formação do corpo lúteo
Diestro
Corpo lúteo activo (secreção de progesterona)
Proestro
Regressão do corpo lúteo Desenvolvimento folicular
13
que ocorre a selecção e crescimento folicular acelerado associado ao aumento da síntese de estradiol (Fortune et al., 2004; Lucy et al., 2004).
O período de estro pode ter uma duração de 2 a 50 horas sendo referido como média as 12 a 18 horas (Stevenson, 2007). A ovulação ocorre geralmente de 24 a 30 horas depois do início do estro (Walker et al., 1996; Jainudeen e Hafez, 2000). Os primeiros sinais de cio coincidem geralmente com o início do pico pré-ovulatório de hormona luteinizante (LH) e da hormona foliculo-estimulante (FSH) (Stevenson, 2007).
A gestação é definida como o período que decorre desde a fecundação até ao parto e na vaca tem uma duração em média de 280-285 dias (Ball e Peters, 2004), embora possa variar entre os 270 e os 292 dias (Stevenson, 2007).
Após o parto, as vacas apresentam um período de aciclia, geralmente designado por anestro pós-parto. O anestro pós-parto corresponde ao intervalo entre o parto e a primeira ovulação (Jainudeen e Hafez, 2000; Stevenson, 2007) ou até à primeira manifestação comportamental de cio (Peter et al., 2009). O intervalo do parto até à primeira ovulação apresenta variações, podendo ir dos 10 aos 110 dias (Jainudeen e Hafez, 2000) conforme a raça da vaca, nutrição, produção de leite, estação do ano e presença do vitelo (Ball e Peters, 2004). Nas vacas de leite a ovulação do primeiro folículo dominante após o parto ocorre geralmente entre os dias 14-21 dias (Lucy, 2001). No entanto cerca de 20 a 25% das vacas ainda se mantêm em anestro 60 dias pós-parto (Stevenson, 2007).
O anestro pós-parto engloba dois eventos indispensáveis à fertilidade da vaca: a involução do útero e o reinício da actividade ovárica. A involução uterina geralmente está completa por volta do dia 30 pós-parto (Ball e Peters, 2004). No entanto, para outros autores a involução uterina completa ocorre em média um pouco mais tarde, por volta dos 45 dias pós-parto podendo variar dos 32 aos 50 dias (Jainudeen e Hafez, 2000).
Aos 10 dias depois do parto já se pode observar ondas consecutivas de desenvolvimento folicular (Espinosa, 2009) e num estudo realizado por Savio et al. (1990) com vacas de leite, os autores observaram que o primeiro folículo dominante que se desenvolveu pós-parto ovulou em 74% das vacas.
2. Dinâmica folicular
Na vaca reconhece-se a existência de três ondas de desenvolvimento folicular em cada ciclo. Embora esta seja a regra, existem algumas fêmeas que apresentam apenas duas ondas de desenvolvimento folicular (Savio et al., 1988; Sirois e Fortune, 1988; Noseir, 2003) enquanto outras apresentam quatro ondas (Sirois e Fortune, 1988; Bleach et al., 2004). Esta é uma característica que se mantém de forma consistente numa vaca, e explica as variações que se observam na duração do ciclo entre animais.
14
A primeira onda de desenvolvimento folicular inicia-se logo após a ovulação, no dia 0 do ciclo éstrico (considerando-se o dia 0 como o dia da ovulação), a segunda no dia 8 ou 9 e a terceira no dia 15 ou16 (Ginther et al., 1989a; Adams et al., 2008). Podem existir diferenças nos dias em que se iniciam as ondas de desenvolvimento folicular caso a vaca apresente duas ou quatro ondas foliculares. (Townson et al., 2002; Bleach et al., 2004).
A primeira onda é iniciada pelo pico de FSH que se segue 20 a 30 horas após o pico pré-ovulatório de LH (Hafez et al., 2000). À emergência de todas as ondas foliculares está associado uns aumentos transitórios detectáveis na FSH, que precedem o aparecimento ecográfico de cada onda em 1 a 2 dias (Stevenson, 2007).
Cada onda de crescimento folicular apresenta quatro fases: recrutamento folicular;
selecção do folículo dominante; dominância e;
atrésia folicular ou ovulação (Sunderland et al., 1994; Diskin et al., 2002).
O recrutamento consiste na mobilização de folículos antrais de 3 mm (15-25 folículos; Roche, 2004), para uma onda de crescimento folicular (Kanitz, 2003) e ocorre todos os 7 a 10 dias do ciclo éstrico (Roche, 2004). Ao recrutamento está associado um aumento da FSH, sendo este essencial para que os folículos sejam recrutados (Adams et al., 1992; Ginther et al., 1998; Mihm e Bleach, 2003).
A seguir ao recrutamento sucede-se a selecção que é o processo pelo qual apenas um dos folículos da onda de crescimento folicular é seleccionado para se tornar dominante (Beg e Ginther, 2006). Quando o folículo de maior tamanho da onda de crescimento, alcança um diâmetro em média de 8.5 mm (aproximadamente 3 dias de crescimento) ocorre uma diferenciação entre o futuro folículo dominante e os restantes folículos subordinados, designando-se este processo por divergência (Ginther et al., 1996; Beg e Ginther, 2006). Antes da divergência todos os folículos em crescimento apresentam a capacidade de se tornarem dominantes, no entanto, após a divergência, o folículo de maior tamanho tende a tornar-se o folículo dominante, enquanto todos os outros regridem (Austin, 2001). A divergência é acompanhada por uma diminuição da concentração sanguínea de FSH. Esta diminuição é devida a um aumento da libertação de estradiol na corrente sanguínea, produzido pelo folículo de maior tamanho, que suprime as concentrações de FSH abaixo das requeridas pelos folículos de menor tamanho. (Ginther et al., 2000). Com a divergência, o folículo dominante adquire capacidade de responder à LH, e torna-se independente da FSH sérica para o seu crescimento. Para além do estradiol a inibina também contribui para a diminuição da concentração de FSH (Ginther et al., 2001).
15
O folículo dominante pode ser distinguido pelo seu maior tamanho, maior concentração em estradiol (Austin et al., 2001), e aumento dos receptores de LH (Beg e Ginther, 2006). O folículo dominante seleccionado apresenta aproximadamente 10 mm de diâmetro na altura em que adquire a capacidade ovulatória (Kanitz, 2003) que ocorre um dia depois do início da divergência folicular. Pensa-se que a aquisição da capacidade ovulatória envolva um aumento da expressão dos receptores de LH nas células da granulosa do folículo dominante e que esta alteração possa ser importante para o posterior crescimento e maturação do folículo dominante (Sartori et al., 2001).
O folículo dominante apresenta cerca de 16 mm um dia antes da ovulação (Ginther et al., 1989b; Beg e Ginther, 2006). Lucy et al. (2004) referem que o folículo dominante aumenta de tamanho até uma dimensão entre os 14 a 20 mm, e que mantém a dominância por 5 a 7 dias.
Qualquer folículo dominante pode maturar e ovular se o corpo lúteo regredir no tempo apropriado (Fortune et al. 2004). A administração de prostaglandina F2α (PGF2α) durante o
crescimento e fase estática do folículo dominante, conduzindo à regressão do corpo lúteo, desencadeia a ovulação (Stevenson, 2007).
A maturação folicular segue acções coordenadas da LH e da FSH através dos receptores nas células da teca e da granulosa, respectivamente. A LH liga-se às células da teca e estimula a produção de androgénios, os quais subsequentemente se difundem através da membrana basal nas células da granulosa (Fortune e Quirk, 1988). A ligação da FSH às células da granulosa aumenta a actividade da aromatase, que converte os androgénios em estradiol. Por sua vez, os níveis elevados de estradiol e a FSH regulam também os receptores para a LH nas células da granulosa nos folículos pré-ovulatórios em maturação. A ligação da LH e FSH pelas células da granulosa é necessária para a regulação final da maturação folicular e eventual ovulação em resposta a estas hormonas (Stevenson, 2007).
Nas situações em que um folículo dominante não ovula, o que poderá ser devido à presença de concentrações elevadas de progesterona, perde as características de dominância e torna-se atrésico, desenvolvendo-se um novo folículo dominante a partir de uma nova vaga de folículos. As concentrações de estradiol folicular e a secreção de inibina-A diminuem no folículo dominante que vai perdendo a dominância, o que favorece o aumento da concentração sanguínea de FSH dando-se início à fase de recrutamento da onda seguinte (Lucy et al., 2004; Fortune et al., 2004).
Na dinâmica do ciclo éstrico, o aumento do estradiol tem um efeito positivo no hipotálamo induzindo a libertação de hormona libertadora de gonadotropina (GnRH). O estradiol parece desencadear pulsos mais frequentes de GnRH assim como um aumento da resposta hipofisária a cada pulso de GnRH, o que por sua vez leva à indução do pico
pré-16
ovulatório de LH que promove a ovulação do folículo dominante (Hafez et al., 2000). Os títulos de estradiol são máximos antes do pico pré-ovulatório de LH, enquanto que as concentrações de FSH são minimas (Stevenson, 2007). O estradiol é também a hormona responsável por desencadear o comportamento de cio (Nebel, 2003) e pela preparação do tracto reprodutivo para o processo associado com a fecundação (Ball e Peters, 2004). Os picos de síntese de LH e FSH têm geralmente uma duração de 6 a 12 horas (Hafez et al., 2000; Stevenson, 2007), pois a hipófise torna-se refractária à acção da GnRH em resposta à (Stevenson, 2007) diminuição da concentração dos receptores de GnRH ou à depleção do “pool” de gonadotropinas. Por isso, na altura da ovulação os níveis de estradiol, progesterona e LH são baixos (Hafez et al, 2000).
3. Luteogénese e luteólise
Após a ovulação as células da teca e da granulosa diferenciam-se em células lúteas pequenas e células lúteas grandes respectivamente, formando o corpo lúteo responsável pela secreção de progesterona. Os dois tipos de células do corpo lúteo secretam progesterona. As células lúteas pequenas apresentam receptores de LH e respondem melhor à LH in vitro do que as células lúteas grandes em termos de secreção de progesterona (Stevenson, 2007). À medida que o corpo lúteo cresce as células lúteas pequenas transformam-se em células lúteas grandes (Alila e Hansel, 1984). As células lúteas pequenas representam cerca de 20% do total da população de células lúteas e contribuem com cerca de 15% da secreção de progesterona pelo corpo lúteo. O restante é produzido pelas células lúteas grandes (Stevenson, 2007). Para além de receptores para a LH, as células lúteas grandes possuem também receptores para a prostaglandina E2 (PGE2) e PGF2α, além de produzirem neurofisina e oxitocina (OT) (Guldenaar
et al., 1984; Harrison et al., 1987).
A luteólise é o processo pelo qual ocorre a regressão do corpo lúteo e dá-se por volta do dia 15 a 17 do ciclo éstrico (Ball e Peters, 2004; Choudhary, 2005). A PGF2α é o factor
luteolítico, e tem uma origem uterina. Para que a PGF2α actue é necessário existir uma
determinada quantidade de 17β-estradiol (17βE2) em circulação, uma vez que esta hormona
determina os teores de oxitocina e seus receptores, tanto ao nível do útero como do corpo lúteo (Stevenson, 2007).
O CL no início de função é refractário à acção da PGF2α, até aos cerca dos 5 a 6 dias. O
mecanismo responsável pela ausência de resposta luteolítica do corpo lúteo neste período ainda não foi totalmente esclarecido (Choudhary, 2005; Shirasuma et al., 2008), embora esteja descrito que a concentração e afinidade de receptores específicos para a PGF2α nas células
lúteas de bovinos são similares nos dias 2, 4, 6 e 10 do ciclo éstrico (Wiltbank et al., 1995; Stevenson, 2007). Este tema mantém-se controvérso, existindo alguns autores que associam a
17
incapacidade de a PGF2α ter capacidade luteolítica antes do dia cinco do ciclo éstrico ao facto
de existir um escasso número de receptores até essa altura (Diskin et al., 2002; Santos et al., 2003; Cavalieri et al., 2006; Lane et al., 2008) ou por se fazer sentir uma acção antiluteolítica (Espinosa, 2009). Na fase inicial da gestação, cerca de 20 dias após a concepção, as células lúteas grandes desaparecem, permanecendo as células lúteas pequenas, algumas das quais se expandem e tornam-se células lúteas grandes no corpo lúteo de gestação (Stevenson, 2007).
Os pulsos marcados de PGF2α produzidos pelo útero na ausência de um embrião viável
induzem regressão ou lise do corpo lúteo, conduzindo ao final da fase lútea (McCracken et al., 1999; Skanzynski et al., 2001). Após 14 dias de influência da secreção de progesterona, o endométrio secreta pulsos de PGF2α, cada um com uma duração aproximada de 6 horas, por
um total de cerca de 36 horas. A PGF2α causa um declínio imediato de progesterona e
oxitocina para níveis basais dentro de 6 a 10 horas na circulação periférica e no conteúdo do corpo lúteo (Stevenson, 2007). Vários trabalhos sugerem que a PGF2α produzida pelo corno
uterino adjacente ao ovário que tem o corpo lúteo é transferida da veia uterina efluente na artéria ovárica através de um mecanismo de transferência contra corrente (McCracken et al., 1999).
Sabe-se que os efeitos luteolíticos da PGF2α ocorrem em parte através da intervenção
directa sobre as células lúteas, mas a sua acção é também obtida de forma indirecta através da redução do fluxo sanguíneo ao corpo lúteo (Niswender et al., 2000; Stevenson, 2007). A PGF2α apresenta propriedades vasoconstritoras, mas ainda não foi determinado se a
diminuição do fluxo sanguíneo do corpo lúteo, que tanto ocorre na luteólise espontânea como na induzida pela PGF2α exógena, é uma causa ou consequência da luteólise (Stevenson,
2007).
A produção de estradiol pelo folículo dominante após um período prolongado de influência progestagénica, como se observa na fase final do desenvolvimento folicular, desencadeia o processo de luteólise através da indução da produção de PGF2α (Araújo et al.,
2009). Esta é produzida em consequência da estimulação dos receptores endometrais da oxitocina pelos níveis crescentes de estradiol, que quando ocupados activam a fosfolipase A2 e
a cascata do ácido araquidónico. O ácido araquidónico é o precursor da síntese uterina de PGF2α (Sílvia et al., 1991; Stevenson, 2007).
18
Figura 2 - Aumento e declínio das hormonas reprodutivas durante o ciclo éstrico. Fonte:
http://www.cnpgc.embrapa.br/publicacoes/doc/doc48/03cicloestral.html
4. Maneio reprodutivo na exploração: a detecção do cio
O cio é a exibição da receptividade sexual por parte da fêmea, que se observa no estro, que na vaca se manifesta durante algumas horas, através de alterações físicas e comportamentais (Nebel, 2003).
A duração do cio tem uma média de 12 a 18 horas (Allrich, 1994; Jainudeen e Hafez, 2000), mas em alguns casos pode encontrar-se reduzida a pouco mais de 30 minutos, enquanto que noutros se pode prolongar até 35.8 horas (O’Connor, 2007). A duração e a intensidade com que a fêmea manifesta o cio pode ser influenciado por factores exógenos e endógenos. A duração média do cio pode ser encurtada por factores ambientais, como por exemplo quando os animais estão estabulados em permanência em pisos de cimento ou quando as temperaturas ambientes são demasiado elevadas (Britt et al., 1986). De acordo com Britt e colaboradores (1986), as vacas que se encontram em piso de terra evidenciam um maior número de montas e reflexos de imobilidade e uma duração do cio superior aos de vacas que se encontram em piso de cimento. De entre os factores endógenos que mais interferem com a duração e intensidade do cio em fêmeas de aptidão leiteira inclui-se o nível de produção de leite (Lopez, 2004). Segundo o estudo realizado por Dransfield et al. (1998) o cio tem uma duração em média de 7 horas, com 25% das vacas a apresentarem cios de curta duração.
A correcta detecção do cio é indispensável à fertilidade potencial da vaca, e particularmente relevante se a fêmea for submetida a beneficiação por inseminação artificial (IA), e é também essencial ao sucesso económico da exploração. A detecção insuficiente e/ou imprecisa dos cios leva a IA fora do período fértil, e por consequência menores taxas de concepção e intervalos mais longos entre partos (Rocha et al., 2001; Graves, 2009). De acordo com os resultados obtidos por Sturman et al. (2000) em vacarias de leite, 19% das inseminações são realizadas em vacas na fase lútea ou em fases iniciais da gestação. Neste
19
estudo, as inseminações de vacas gestantes estiveram na origem de 17% dos casos de morte embrionária. Este estudo retrata bem o impacto que tem actualmente a deficiente detecção do estro nos parâmetros reprodutivos em vacarias de leite, com o consequente aumento do intervalo parto/fecundação, do número de IA/fecundação e do intervalo entre partos, já para não falar das consequentes perdas económicas.
O recente declínio na eficiência reprodutiva da fileira leiteira pode estar associado a uma fraca manisfestação do estro (Maillard, 2006; Opsomer et al., 2007a). Esta tem sido explicada pelo facto de a expressão do estro ser um comportamento que resulta das acções do estradiol no hipotálamo, e os teores de estradiol tendem a ser menores em animais de elevada produção, em consequência do aumento do metabolismo hepático (Sangsritavong et al., 2002; Wiltbank et al., 2005). A intensidade (número de montas) e a duração do cio dependem do comportamento individual das vacas assim como das interacções das vacas entre si e com o ambiente em que se inserem (Dransfield et al., 1998).
Vários factores têm sido apontados como podendo justificar a deficiente manifestação comportamental do estro em vacas de leite, entre eles (Gwazdauskas, 1985; Dransfield et al., 1998; Rensis e Scaramuzzi, 2003; Opsomer et al., 2007a):
Confinamento em estábulos com um piso demasiado abrasivo ou liso;
Ocorrência de alterações podais, que tornam a deslocação e as montas dolorosas;
Uma fraca condição corporal ou nutrição deficiente; Stresse térmico;
Desequilíbrios no metabolismo dos esteróides, resultantes de uma síntese deficiente e de um metabolismo acelerado das hormonas esteróides em vacas de leite de alta produção, com consequente decréscimo dos valores circulantes de esteróides endógenos e consequente encurtamento do comportamento de cio nestas vacas (Wiltbank et al., 2005). O aumento da ocorrência de ciclos anormais (curtos ou longos) que se tem vindo a observar nos últimos anos também torna mais difícil predizer quando uma vaca cíclica retorna ao cio (Graves, 2009; Opsomer et al., 2007a, Espinosa, 2009).
Os trabalhos de Keown e Kononoff (2007) referem que cerca de 68% dos cios ocorrem entre as 6 da tarde e as seis da manhã. No entanto, em contraste, estudos publicados anteriormente evidenciaram que a actividade de monta e o início do cio ocorrem de modo uniforme ao longo do dia (Dransfield et al., 1998; Xu et al., 1998; Nebel et al., 2000). É possível que a evolução no tempo da componente genética do efectivo, do nível produtivo dos animais
20
ou da relação entre a produção de leite e o nível nutricional possam justificar as diferenças encontradas entre estes trabalhos.
A observação das vacas para detecção do cio deveria ser feita pelo menos duas vezes por dia, por um período mínimo de 30 minutos (Maillard, 2006; O’Conner, 2007). No entanto, existem autores que defendem que esta duração e frequência são actualmente insuficientes para uma detecção bem sucedida do estro (Dransfield et al., 1998; Diskin e Sreenan, 2000; Graves, 2009), defendendo que o momento ideal para a detecção de cio é de manhã muito cedo (antes de serem ordenadas e alimentadas), ao final da tarde (depois de ordenhadas e alimentadas) e à noite/madrugada (Graves, 2009). O sinal primário, mais evidente, de que a vaca se encontra em cio é o reflexo de imobilidade, que se caracteriza pelo facto de a fêmea permitir a monta (Nebel, 2003; Diskin e Sreenan, 2000; Graves, 2009). Num estudo realizado por Dransfield et al. (1998), a média de saltos por cio numa vaca de leite foi de 8,5 saltos. Existem ainda sinais secundários que poderão auxiliar na detecção de cio em vacas, embora sejam menos fiáveis por serem muito variáveis na indicação do período fértil (Allrich, 1994; Diskin e Sreenan, 2000; Graves, 2009).
Em muitas explorações, a taxa de detecção de cio por meio da observação de sinais indiciadores (Quadro 1) é inferior a 50% (Senger, 1994; Nebel, 2003; Portaluppi e Stevenson, 2005; Maillard, 2006). Nos trabalhos de Rocha et al. (2001), desenvolvidos no Norte de Portugal, tendo por base os dados da taxa de não retorno aos 90 dias e no registo de nascimento de vitelos, foi reportada uma taxa média de detecção de cio de 38.1%. Estes autores encontraram ainda diferenças significativas na taxa de detecção de cio entre explorações (variando entre 14.8 a 60.8%).
Dada a dificuldade que se observa na correcta detecção do cio, foram desenvolvidos diversos dispositivos ou metodologias para facilitar a sua detecção. Entre estes, destacam-se os detectores de monta sensíveis à pressão (Walker et al., 1996; Dransfield et al., 1998; Nebel, 2003; Diskan e Sreenan, 2000), os marcadores de tinta (O’Connor, 2007; Graves, 2009), os podómetros/coleiras (Senger, 1994; O’Connor, 2007; Diskan e Sreenan, 2000), os arreios submandibulares, a colocação de câmeras de vídeo no local, o recurso a rufiões, a medição da impedância do muco vaginal (também conhecido por método de Draminsky; Jainudeen e Hafez, 2000; Senger, 1994; O’Connor, 2007) e a medição da progesterona no sangue ou leite de vacas suspeitas em cio (Jainudeen e Hafez, 2000; Pembertonet et al., 2001; Graves, 2009).
21 Quadro 1 – Sinais primários e secundários de cio.
Tipo de sinal Manifestação comportamental/física Referências
Sinais primários •Reflexo de imobilização Diskin e Sreenan, 2000; Yoshida e Nakao, 2005; Cutullic et al., 2009
Sinais secundários
•Mucosa vaginal edematosa, lisa, húmida e avermelhada
Diskin e Sreenan, 2000; Nebel, 2003
•Muco viscoso e transparente pendente na comissura ventral da vagina
Diskin e Sreenan, 2000; Nebel, 2003 •Diminuição da ingestão e produção de leite Diskin e Sreenan, 2000; Balls e Peters,
2004; Cutullic et al., 2009 •Aumento ligeiro da temperatura corporal Jainudeen e Hafez, 2000
•Tentativa de montar outras vacas Diskin e Sreenan, 2000; Cutullic et al., 2009
•Cheirar ou lamber com convicção a genitália das outras vacas
Diskin e Sreenan, 2000; O’Connor, 2007
•Aumento da vocalização, com mugidos altos e frequentes
Nebel, 2003, O’Connor, 2007
•Pôr a cabeça por cima das outras Nebel, 2003, O’Connor, 2007 •Aumento do nervosismo, com agitação,
impaciência, aumento da actividade; as vacas ficam irrequietas, movimentam-se com rapidez, agitam a cauda enérgica e frequentemente, e as suas reacções ao contacto físico são mais rápidas
Diskin e Sreenan, 2000; O’Connor, 2007; Ranasinghe et al., 2009
•Reflexo de Flehmen - elevar o lábio superior na inspiração, como que num sorriso, com a cabeça levantada e o pescoço esticado (semelhante ao reflexo de Flehman)
Nebel, 2003; O’Connor, 2007
•Formação de um grupo sexualmente activo na periferia do efectivo
Nebel, 2003
•Alteração de rotinas Balls e Peters, 2004 •Três a quatro dias depois do estro as vacas
podem apresentar corrimentos sero-sanguinolentos, geralmente designados de corrimento metaéstrico. Este sinal indica que dentro de 15 a 20 dias provavelmente se repetirá o episódio de cio
Diskin e Sreenan, 2000; Keown e Kononoff, 2007
Vários estudos indicam que a eficiência da detecção de cio aumenta quando se alia a utilização destes dispositivos à observação visual (Senger, 1994; O’Connor, 2007).
22 5. Momento da inseminação
Para alguns autores o tempo de vida fértil do oócito bovino após a ovulação é de 8 a 12 horas (Ziegler, 2002); no entanto, há quem defenda que este período é mais alargado e poderá chegar às 20 a 24 horas (Jainudeen e Hafez, 2000), diminuindo a viabilidade do oócito ao longo do tempo (Nebel, 2003).
Também os espermatozóides têm um período de vida limitado, habitualmente de 30 a 48 horas (Ziegler, 2002; Nebel, 2003).
Para que ocorra a fecundação é necessário que os espermatozóides atravessem o útero, entrem nos oviductos e completem o processo de maturação, conhecido como capacitação. O tempo necessário para que um número significativo de espermatozóides viáveis e passíveis de sofrer capacitação atinja o local de fecundação requer um mínimo de seis horas, aumentando de forma progressiva o número de espermatozóides que chegam à ampola do oviducto entre as 8 e as 18 horas pós-IA; a partir deste momento, o número de espermatozóides viáveis diminui (Hank, 1983; Wilmut e Hunter, 1984).
Com base neste conhecimento, se a inseminação ocorrer demasiado cedo os espermatozóides perderão capacidade fecundante antes de terem oportunidade de fecundar o oócito. Por outro lado se se atrasa demais a inseminação, o oócito perderá a sua viabilidade, tornando-se incapaz de ser fecundado ou tendendo a originar um embrião com fraca capacidade de sobrevivência.
Uma vez que a ovulação ocorre 10 a 12 horas após o final do estro (Jainudeen e Hafez, 2000), o momento óptimo para a inseminação encontra-se perto do fim do estro. Como em condições práticas as vacas não são observadas continuamente, torna-se difícil a determinação exacta do fim do estro. Considerando o período de vida limitado tanto do oócito como do espermatozóide, existe uma “janela” de cerca de 12 horas durante a qual se obtêm as melhores taxas de concepção. Consequentemente, considerou-se que de um ponto de vista prático está aconselhado utilizar-se a regra manhã/tarde (AM/PM): todas as vacas observadas em cio durante a manhã serão inseminadas durante a tarde. As vacas ainda em cio na manhã seguinte serão inseminadas novamente. As vacas observadas em cio durante a tarde ou à noite serão inseminadas na manhã seguinte (Graves et al., 1997; Graves, 2009).
Num trabalho em que avaliaram a eficiência do momento de beneficiação através das taxas de não retorno, Nebel et al. (1994) compararam a IA realizada de acordo com a regra AM/PM e a inseminação num único momento predefinido do dia (IA realizadas apenas no período da manhã, independentemente do momento da detecção do cio), estes autores não encontraram diferenças significativas nas taxas de não retorno aos 60, 75 e 90 dias: 65.6%, 60.6% e 57.8% para a regra manhã/tarde e 64.6%, 60.1% e 58.4% para a IA uma única vez ao dia, respectivamente aos 60, 75 e 90 dias. Contudo, apesar dos resultados serem semelhantes
23
entre os dois protocolos, os produtores continuam a realizar a IA com base na regra AM/PM (Graves, 2009).
6. Factores que contribuem para a diminuição da taxa de concepção
Nos últimos anos, a diminuição progressiva da taxa de concepção global em explorações intensivas de bovinos de leite, e que rodando os 30 a 40%, tem sido fonte de preocupação para os produtores e veterinários do sector (Lucy et al., 2004). A fraca capacidade de detecção de cios é uma das causas que contribui para o decréscimo da taxa de concepção, apesar de existirem outras (Lopez et al., 2004; Wiltbank et. al, 2005; Thatcher, 2006; Opsomer et al., 2007b), como sejam:
O atraso no retorno à actividade cíclica no pós-parto; Um prolongamento da fase lútea no pós-parto;
O aumento do intervalo entre a primeira IA e a fecundação;
Um aumento do metabolismo das hormonas esteróides em vacas de alta produção;
A consequente diminuição da concentração da progesterona em vacas de leite de alta produção;
O stresse térmico.
A duração do intervalo entre o parto e a primeira ovulação é decorrente do reinício da actividade ovárica e da involução uterina. Se a primeira está dependente da replecção das reservas hipofisárias de LH, que por sua vez podem ser moduladas por diferentes factores ambientais e endógenos (como o balanço energético do animal), o segundo é muito influenciado pelo estatuto hígido do útero, pelo que a presença de inflamação é um importante modulador. O atraso no retorno à actividade cíclica no pós-parto pode estar associado à ocorrência de doenças puerperais, principalmente àquelas que surgem no primeiro mês após o parto, e ainda ser consequência do balanço energético negativo (BEN) no início da curva de lactação (Beam e Butler, 1998; Opsomer et al., 2007c). As vacas em lactação estão geralmente em BEN durante o período pós-parto inicial, uma vez que não obtêm a energia suficiente para as demandas do organismo através da alimentação que recebem. O balanço energético negativo e a perda de peso associado exercem um efeito inibitório tanto directo como indirecto no crescimento e desenvolvimento folicular. A hormona luteinizante e a IGF-I (Insulin-like growth factor I) actuam sinergeticamente para promover o desenvolvimento folicular. Nas vacas em balanço energético negativo, os pulsos de LH e as concentrações plasmáticas de IGF-I
24
encontram-se diminuídos, o que parece comprometer o crescimento folicular durante o período de pós-parto inicial e atrasar o retorno à ciclicidade (Lucy, 2000). Sabe-se que o folículo dominante nas vacas em balanço energético negativo requer mais tempo e um tamanho maior para estabelecer as concentrações de estradiol capazes de desencadear a ovulação (Lucy, 2001). Foi demonstrado que o metabolismo fisiológico das hormonas esteróides se encontra aumentado devido às exigências da alta produção (Lopez et al., 2004; Wiltbank et. al, 2005) Este aumento do metabolismo leva a uma diminuição dos níveis de estradiol endógeno. Em consequência, o folículo pré-ovulatório e o oócito ficam expostos a um prolongamento nos pulsos de LH que pode levar à ovulação de um folículo super-estimulado ou a um oócito prematuramente activado e assim promover uma diminuição da fertilidade (Wiltbank et. al, 2005).
Outra das causas que contribui para a diminuição da taxa de concepção é o prolongamento da fase lútea. Opsomer et al. (1998) descreveram um aumento de fases lúteas anormais no pós-parto em vacas de leite de alta produção. Cerca de 20% das vacas em estudo apresentavam um aumento da concentração da progesterona por mais de 20 dias sem que tivessem sido submetidas anteriormente a uma IA (não podendo portanto estar gestantes). A alteração na fase lútea das vacas de leite dificulta o maneio reprodutivo. As fases lúteas longas atrasam a inseminação de vacas que não estão em protocolos de inseminação e torna mais difícil prever quando é que uma vaca cíclica retorna ao cio (Opsomer et al., 2007b). No entanto, a informação disponível na literatura é controversa, e Espinosa (2009) defende a ocorrência de ciclos curtos durante o pós-parto de vacas devido a um subdesenvolvimento nos corpos lúteos com anomalias na função endócrina, não se normalizando os ciclos até uma segunda ou terceira ovulação. Comparando aptidões produtivas distintas (carne e leite) o anestro pós-parto é mais curto nas raças de produção de leite do que nas de carne que amamentam os seus vitelos, sendo nestas últimas a amamentação e a presença do vitelo o principal factor condicionante do arranque da actividade cíclica (Espinosa, 2009).
Estudos recentes demonstraram que o BEN, a nutrição, os distúrbios e as doenças pós-parto são factores de risco para o atraso na ciclicidade e prolongamento da fase lútea (Opsomer et al., 2007a).
Outros factores que podem contribuir para a diminuição da taxa de concepção e para o aumento do intervalo entre a primeira IA e a concepção são a falha na fecundação, que pode estar associada a má qualidade do sémen ou a má técnica de inseminação, ou ainda a um oócito envelhecido; um aumento da mortalidade embrionária ou fetal, que comprometa o sucesso da gestação; e também problemas na exibição do cio (Opsomer et al., 2007a).
Em paralelo com o decréscimo da taxa de fertilidade à primeira IA, tem-se observado ainda um aumento das gestações gemelares, presumivelmente causado por taxas de ovulação
25
elevadas (Kinsel et al., 1998), que pode ser consequência de uma deficiente função folicular e de uma sinalização inadequada do estradiol em vacas de leite de alta produção (Fricke e Wiltbank, 1999). O estradiol e a inibina são essenciais para controlar o crescimento folicular através do seu efeito inibitório na FSH durante a onda folicular. A diminuição da concentração sanguínea de estradiol nas vacas em lactação reduz o feedback negativo sobre a FSH durante a onda folicular, conduzindo a um aumento de FSH que favorece a divergência e dominância de mais do que um folículo nas vacas em lactação. Existe também a possibilidade de outros mecanismos que controlam o crescimento folicular, seja a inibina ou outros factores ainda não descobertos, estarem comprometidos pela lactação (Lucy, 2001). A gestação gemelar é indesejável porque a produção de leite e a fertilidade após o nascimento de gémeos diminuem ainda mais (Beerepoot et al., 1992).
A diminuição na taxa de concepção também pode ser devida a uma diminuição da concentração da progesterona nas vacas de leite de alta produção (Lucy, 2001) que apresentam uma metabolização mais rápida da progesterona conduzindo a uma diminuição da sua concentração (Wiltbank et al., 2006). A progesterona é requerida para a gestação e existe uma associação entre baixos níveis de progesterona e a morte embrionária precoce, apesar de ainda não ter sido estabelecida uma relação directa entre progesterona baixa e a perda embrionária. Desconhece-se a concentração mínima absoluta de progesterona necessária para manter a gestação numa vaca (Lucy, 2001).
A taxa de concepção também pode ser diminuida pelo stresse térmico, tendo-se verificado que este reduz a secreção de LH, conduzindo a uma secreção inadequada de estradiol pelos folículos dominantes, diminuindo as concentrações plasmáticas de estradiol (Rensis e Scaramuzzi., 2003), que é um componente crítico da cascata luteolítica (Stevenson, 2007), o que atrasa a luteólise em vacas em lactação conduzindo a um alongamento na fase lútea das vacas (Lucy, 2001). Estes acontecimentos resultam numa diminuição da qualidade do oócito, falha na implantação e infertilidade (Hansen, 1997; Rensis e Scaramuzzi, 2003).
Com a meta de um vitelo por ano (Jainudeen e Hafez, 2000; Stevenson, 2007; Espinosa, 2009), o intervalo parto-fecundação limita-se a cerca de 85 dias (Opsomer et al., 1998; Lyimo et al., 2000), durante os quais deve ocorrer a involução do útero, o reinício da actividade ovárica e a adequada detecção de cios. Por consequência, torna-se fácil de entender a importância crescente da implementação de protocolos de sincronização do estro, para submeter vacas à IA no tempo apropriado, como forma de aumentar as taxas de concepção.
26 7. Métodos de controlo do estro e/ou ovulação
Os protocolos de manipulação do estro e/ou ovulação podem ser agrupados em dois tipos principais: os que sincronizam o estro e os que sincronizam a ovulação. Os protocolos de sincronização do estro requerem a detecção do cio. Embora tendam a apresentar uma maior concentração dos animais que evidenciam comportamento de cio, apresentam também uma janela temporal mais ou menos variável em relação à altura da inseminação, pouco controlo sobre a altura da inseminação, estando o momento da inseminação sob dependência da capacidade da detecção de cios na exploração (Santos et al., 2003). Por outro lado, os protocolos de sincronização da ovulação permitem a inseminação num momento pré-determinado, ajustado aos produtos administrados, e prescindem da detecção do cio (Pursley et al., 1995). Este tipo de protocolos controlam a onda de desenvolvimento folicular e, por conseguinte, a ovulação (Murugavel et al., 2003a).
Um protocolo de sincronização ideal deve proporcionar um início do estro preciso e uma fertilidade muito próxima da obtida em ciclos não manipulados, com taxas de gestação superiores a 55% após uma única inseminação artificial (Austin et al., 1999).
Todos os métodos farmacológicos de sincronização do estro devem ser considerados como ferramentas de maneio reprodutivo úteis, cujo objectivo principal é aumentar a eficiência reprodutiva. Em paralelo, permitem melhorar a organização da reprodução ou corrigir alguma deficiência organizacional. Em alguns casos os sistemas de manipulação do estro podem ser usados como tratamento para certos transtornos reprodutivos, tais como o sub-estro ou “cio silencioso” ou a doença ovárica quística (Bartolome et al., 2000; Rensis et al., 2008). Contudo, os métodos farmacológicos de manipulação do ciclo éstrico não devem ser nunca considerados um substituto do maneio adequado. Caso surjam respostas negativas ao programa de sincronização, estas poderão ser devidas a diversos factores, nomeadamente, a nutrição, a fase de pós-parto, o estado de saúde, factores genéticos individuais, incumprimento dos protocolos (Lucy et al. 2004; Lopez, 2006; Maillard, 2006; Cavalieri et al., 2006), variação do metabolismo interno em particular das taxas de excreção da progesterona (Wiltbank et al., 2005).
Em vacas cíclicas o ciclo éstrico pode ser manipulado de três formas (Lucy et al., 2004; Thatcher et al., 2006):
Através do uso de prostaglandinas, para induzir uma luteólise precoce;
Através do uso sequencial de prostaglandinas e análogos da GnRH, para obter o desenvolvimento folicular sincronizado após indução de luteólise;
27 7.1.Protocolos de sincronização do estro
7.1.1. Indução do estro com prostaglandinas
O efeito luteolítico da PGF2α nas vacas foi inicialmente descrito na década de 70
(Inskeep, 1973; Hafs et al., 1974; Lauderdale et al., 1974; Wenkoff, 1975). Desde então a PGF2α e os seus análogos sintéticos têm sido usados no controlo do ciclo em bovinos devido à
sua capacidade para induzir a regressão do corpo lúteo e por consequência estimular o desenvolvimento de uma nova onda folicular ovulatória e a sincronização do estro (Odde, 1990; Murugavel et al., 2003a, Cavalieri et al., 2006). A administração de uma ou várias injecções de prostaglandina para regredir um corpo lúteo responsivo faz com que a vaca retorne ao estro em 2 a 7 dias (Figura 3) (Lauderdale et al., 1974; Santos et al., 2003).
Para que o tratamento com PGF2α seja bem sucedido, ou seja obtenha efeitos
luteolíticos, as vacas devem estar na fase de diestro do ciclo éstrico. O corpo lúteo da vaca é geralmente responsivo à PGF2α apenas após o quinto ou sexto dia do ciclo éstrico (Wenkoff,
1975; Diskin et al., 2002; Murugavel et al., 2003a; Cavalieri et al., 2006), e até ao dia 15-17, após o qual a luteólise endógena foi já desencadeada (Ball e Peters, 2004; Choudhary, 2005). A aplicação única de uma dose luteolítica de prostaglandinas obriga à identificação das vacas que apresentam um CL responsivo no ovário, para rentabilizar a aplicação do fármaco, mas o que consequentemente acarreta custos acrescidos (Ball e Peters, 2004; Cavalieri et al., 2006).
Devido ao facto da PGF2α não ter efeito sobre os folículos em desenvolvimento, e não
ter também controlo sobre a onda de folículos emergente, as vacas num protocolo deste tipo entram em cio em dias diferentes depois da sua administração, havendo pouca precisão sobre o momento da inseminação ou da ovulação (Diskin et al., 2002; Santos et al., 2003; Cavalerie et al., 2006). O tempo desde a administração de PGF2α à ovulação depende da maturidade e
tamanho dos folículos da onda emergente, e ainda do facto de a onda existente já ter entrado no processo de atrésia. Se estiver presente um folículo dominante funcional por altura do tratamento, os animais entrarão em cio no espaço de 2 a 3 dias; no entanto, se os animais se encontrarem numa fase de pré-dominância da onda folicular será necessário um período adicional de 2 a 4 dias para que um folículo se torne dominante, o que alonga e torna mais variável o intervalo desde o tratamento ao início do cio (Diskin et al., 2002).
Na tentativa de ultrapassar estes inconvenientes, foi desenvolvido um protocolo que consiste em duas administrações de PGF2α dadas a intervalos de 7, 11 ou 14 dias. Neste
protocolo as vacas que na primeira administração se encontrem numa fase do ciclo éstrico que não o diestro, apresentarão um corpo lúteo funcional no momento da segunda administração de PGF2α (Figura 3) (Folman et al., 1990; Rosenberg et al., 1990; Kristula et al., 1992;
28
reprodutiva de vacas tratadas (Folman et al., 1990; Rosenberg et al., 1990) e uma maior taxa de concepção aos 30 dias (Folman et al., 1990)
Alguns autores aconselham a administração de PGF2α antes do final do período
voluntário de espera (PVE) de modo a aumentar o desempenho reprodutivo das vacas e reduzir o número de dias em aberto (Pankowski et al., 1995; Stevenson, 2001; Santos et al., 2003). Stevenson (2001) recomenda a administração de PGF2α 14 dias antes do fim do PVE;
este autor não aconselha a inseminação das vacas após a primeira administração, ainda que mais de 50% das vacas exibam cio em resposta à primeira administração. A segunda administração, a ser realizada no final do PVE, possibilita a ocorrência do primeiro serviço numa altura em que as vacas já são elegíveis para a IA. Com um protocolo similar, Santos et al. (2003) obtiveram uma taxa de detecção de cios entre 50% a 60% tendo utilizado duas administrações de PGF2α nos primeiros 50 dias do pós-parto, aconselhando a adopção deste
regime protocolar nas explorações.
A administração de PGF2α durante o período inicial do pós-parto leva a um aumento das
taxas de concepção ao primeiro serviço, o que poderá estar relacionada com os benefícios associados a uma melhoria da actividade uterina diminuindo desse modo o intervalo entre o parto e a concepção (Murugavel et al., 2003b; Kasimanickam et al., 2006). A administração de PGF2α durante o período de pós-parto não só induz a luteólise, como estimula os mecanismos
de defesa uterina, acelerando a expulsão das loccia e dos microrganismos que colonizam o útero no pós-parto (Kasimanickam et al., 2006). No entanto, Burton e Lean (1995), realizando uma metanálise sobre o efeito da prostaglandina F2α, administrada no pós-parto sobre o
desempenho reprodutivo de vacas de leite, concluíram que a administração sistémica de PGF2α
não diminui de forma significativa o número de dias em abertos, mesmo quando a sua aplicação se restringe a vacas com puerpério anormal (redução de 2,6 dias para todas as vacas e 3,3 dias para as vacas com puerpério anormal).
Posteriormente, foi desenvolvido um protocolo de sincronização para vacas de leite em lactação no qual são administradas três injecções de PGF2α (Figura 3). Neste protocolo,
conhecido como Targeted Breeding, todos os animais que não foram detectados em cio após a primeira ou segunda administração de PGF2α intervaladas de 14 dias, são tratados com uma
terceira administração de PGF2α aplicada com idêntico intervalo, sendo a IA realizada após
detecção do cio ou realizada às 72 a 80 horas (Nebel e Jobst, 1998; Stevenson, 2001; Murugavel et al., 2003a). No entanto, a realização de inseminação artificial a tempo fixo (IATF) neste protocolo não produz taxas de gestação aceitáveis devido ao inconsistente desenvolvimento folicular e ovulação. Nos trabalhos de Nebel et al. (1994), a taxa de não retorno aos 75 dias foi de cerca de 36% quando as vacas foram inseminadas após o
29
tratamento com PGF2α sem detecção de cio, em contraste com uma taxa de não retorno de
63.4% quando foram inseminadas com base na detecção de cio.
Figura 3 - Protocolos baseados no uso de PGF2α para sincronização do estro em bovinos (adaptado de Cavalieri et
al., 2006). (a) PGF2α administrada apenas às vacas com um CL e detecção do cio nos próximos 2-7 dias; (b) 2
administrações de PGF2α espaçadas 11 a 14 dias, detecção de cio e eventual IA realizadas nos 2-7 dias seguintes à
administração de PGF2α; (c) programa “Targeted Breeding”, administração de 3 doses de PGF2α espaçadas em 14
dias, a IA é realizada após detecção de cio ou realizada 72 a 80 horas após a terceira administração de PGF2α.
7.1.2. Controlo da onda folicular e indução do estro com prostaglandinas (Sincronização selectiva - Selectsynch)
Uma maior precisão do início do estro será conseguida através do controlo da onda folicular emergente, o que permitirá que um folículo dominante seleccionado recentemente esteja presente no momento do tratamento com PGF2α, imediatamente antes da indução do
cio, conseguindo-se deste modo uma maior precisão do estro e uma taxa de concepção adequada (Santos et al., 2003).
Detecção do cio e IA PGF2α (c) Detecção do cio e IA PGF2α PGF2α 14 d IA d 0 Detecção do cio e IA PGF2α 11-14 d (b) Detecção do cio e IA PGF2α d 0 Detecção do cio e IA PGF2α 2-7 d (a) 72-80 h 14 d d 0
30
O turnover folicular pode ser modulado através da administração de GnRH ou de estradiol. O tratamento das vacas com GnRH ou um seu análogo resulta num pico pré-ovulatório de LH e na ovulação do folículo dominante, com subsequente recrutamento de uma nova onda de desenvolvimento folicular (Kesler e Constantaras, 2004). No entanto, o uso de GnRH para o recrutamento de uma nova onda folicular apenas será efectivo se o folículo dominante for responsivo à LH. Dez milímetros de diâmetro parece ser o tamanho mínimo para os folículos serem responsivos à LH (Ginther et al., 1996; Sartori et al, 2001). Quando a GnRH ou seus análogos são administrados com o objectivo de induzir a ovulação e recrutamento de uma nova onda folicular, observa-se uma melhor resposta se a administração ocorrer entre os dias 5 e 9 do ciclo (Vasconcelos et al, 1999), que coincide com o período de tempo em que o folículo dominante da primeira onda folicular é responsivo ao LH (Ginther et al., 1996). A utilização de GnRH seguida 7 dias mais tarde pela administração de uma dose luteolítica de PGF2α resulta numa sincronização mais apertada do estro com uma elevada taxa detecção de
cios (Santos et al, 2001).
Figura 4 - Representação esquemática do protocolo Selecsynch
Uma alternativa ao uso de GnRH é o recurso aos ésteres do estradiol, como o cipionato de estradiol (ECP) ou benzoato de estradiol (EB) (Santos et al., 2003). Estes têm uma actuação diferente consoante os níveis séricos de progesterona na altura da sua administração. Na presença de elevados níveis séricos de progesterona (provenientes de fonte endógena ou exógena) os ésteres de estradiol actuam de forma a reduzir a secreção de LH, induzem a atrésia do folículo dominante e a emergência de uma nova onda folicular. Na presença de níveis séricos baixos de progesterona, estes induzem a libertação de GnRH que por sua vez induz um pico de LH que promove a ovulação. O intervalo que decorre desde a administração dos ésteres de estradiol até à emergência de uma nova onda folicular depende da dose e da altura do ciclo éstrico em que estes são administrados (Lucy et al., 2004)
Actualmente, com a preocupação com o uso de estrogénios em animais destinados ao consumo humano a sua utilização no maneio reprodutivo é limitada na Europa, ficando deste
d 0 PGF2α d 7 Detecção do cio + IA GnRH