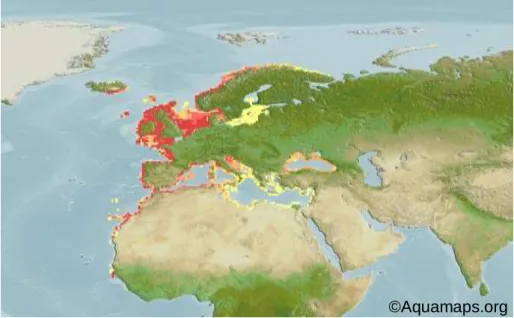
Effects of antidepressant fluoxetine on behavior and gene expression of European sea bass juveniles
60
0
0
Texto
Imagem


+7
Documentos relacionados