UNIVERSIDADE FEDERAL DO CEARÁ FACULDADE DE MEDICINA
DEPARTAMENTO DE PATOLOGIA E MEDICINA LEGAL PROGRAMA DE PÓS- GRADUAÇÃO EM MICROBIOLOGIA MÉDICA
TEREZINHA DE JESUS SANTOS RODRIGUES
CEPAS DO COMPLEXO Candida parapsilosis DE ORIGEM ANIMAL: CLASSIFICAÇÃO TAXONÔMICA, SENSIBILIDADE ANTIFÚNGICA E
ATRIBUTOS DE VIRULÊNCIA IN VITRO
TEREZINHA DE JESUS SANTOS RODRIGUES
CEPAS DO COMPLEXO Candida parapsilosis DE ORIGEM ANIMAL: CLASSIFICAÇÃO TAXONÔMICA, SENSIBILIDADE ANTIFÚNGICA E
ATRIBUTOS DE VIRULÊNCIA IN VITRO.
Dissertação apresentada ao Programa de Pós-Graduação em Microbiologia Médica da Universidade Federal do Ceará, como requisito parcial à obtenção do título de Mestre em Microbiologia Médica.
Orientador: Prof. Dr. Marcos Fábio Gadelha Rocha.
.
Dados Internacionais de Catalogação na Publicação Universidade Federal do Ceará
Biblioteca de Ciências da Saúde
R617c Rodrigues, Terezinha de Jesus Santos.
Cepas do complexo Candida parapsilosis de origem animal: classificação taxonômica, sensibilidade antifúngica e atributos de virulência in vitro / Terezinha de Jesus Santos Rodrigues. – 2013.
39 f. : il.
Dissertação (Mestrado) - Universidade Federal do Ceará. Faculdade de Medicina. Programa de Pós-Graduação em Microbiologia Médica, Fortaleza, 2013.
Orientação: Prof. Dr. Marcos Fábio Gadelha Rocha.
1. Animais. 2. Candida. 3. Virulência. I. Título.
TEREZINHA DE JESUS SANTOS RODRIGUES
CEPAS DO COMPLEXO Candida parapsilosis DE ORIGEM ANIMAL: CLASSIFICAÇÃO TAXONÔMICA, SENSIBILIDADE ANTIFÚNGICA E
ATRIBUTOS DE VIRULÊNCIA IN VITRO
Dissertação apresentada ao Programa de Pós-Graduação em Microbiologia Médica da Universidade Federal do Ceará, como requisito parcial à obtenção do título de Mestre em Microbiologia Médica.
Data da defesa: 29/ 10/ 2013
Banca Examinadora
___________________________________ Prof. Dr. Marcos Fábio Gadelha Rocha
Universidade Estadual do Ceará Orientador
____________________________________ Profa. Dra. Erika Helena Salles de Brito
Universidade da Integração Internacional da Lusofonia Afro-Brasileira Examinadora
____________________________________ Profa. Dra. Ana Karoline da Costa Ribeiro
Universidade de Fortaleza Examinadora
____________________________________ Prof. Dr. José Júlio Costa Sidrim
Aos meus Pais (in memoriam), por terem
me gerado e Zenaide Ramos Lopes da
Silva por me cria r e orienta r com
ensinamentos que me tornaram a pessoa
que hoje sou. À minha filha Bruna
Rafaella Santos Rodrigues, por sua
existência, que me deu força e coragem
para continuar na luta.
AGRADECIMENTOS
À Universidade Federal do Ceará
Ao Programa de Pós-Graduação em Microbiologia Médica, da Universidade Federal do Ceará.
Ao Centro Especializado em Micologia Médica (CEMM), da Universidade Federal do Ceará.
À Deus por ter me concedido o Dom da vida, por me iluminar em toda caminhada.
Á minha filha Bruna Rafaella Santos Rodrigues meu maior incentivo para continuar na
batalha pela vida.
Aos Professores Doutores do Centro Especializado em Micologia Médica (CEMM) por todo
o apoio que sempre me prestaram: especialmente ao Prof. Dr. José Júlio Costa Sidrim, com
quem aprendi tudo que sei em micologia.
À Professora Dra. Raimunda Sâmia Nogueira Brilhante pelo apoio e dedicação que sempre
me disponibiliza, pelas vezes que me ajudou nas horas difíceis.
Ao Professor Dr. Marcos Fábio Gadelha Rocha, por sua sabedoria, além da paciência e
dedicação para me orientar neste trabalho.
À Professora Dra. Rossana de Aguiar Cordeiro pelos ensinamentos e por me Co- orientar no
início deste trabalho.
À Dra. Débora Castelo Branco de Souza Collares Maia, pelos ensinamentos, apoio e ajuda
que me deu.
À Professora Vaulice Sales Café e ao Professor Dr. José Luciano Bezerra Moreira, por terem
me apoiado no início de tudo, que sempre me transmitiram muito de seus conhecimentos e
também, me ajudaram a superar os momentos mais difíceis que enfrentei. Muito do que sei
em Microbiologia, aprendi com eles.
À Professora Dra. Tereza de Jesus Pinheiro Gomes Bandeira pelo apoio.
À médica infectologista Silviane Praciano Bandeira pela amizade e pelos ensinamentos de
À Dra. Camila Alencar Moreira pelo tempo que me disponibilizou, no início deste trabalho.
Ao amigo e companheiro de trabalho Daniel Teixeira Lima pelo apoio.
A amiga e comadre Francisca Mônica Sousa café, pelo incentivo e apoio de sempre.
A secretária do PPGMM. Carolinda Vilma Soares de Oliveira pela ajuda prestada.
Aos amigos, especialmente a Glaucia Morgana de Melo Guedes, Giovanna Riello Barbosa, Ramila
de Brito Macedo, Carlos Eduardo Cordeiro Teixeira, Paula Bittencourt Vago, Francisca Jakelyne de
Farias Marques, Pedro Henrique de Aragão Rodrigues, Joyce Fonteles Ribeiro, Camila Fernandes;
Jonathas Sales de Oliveira, Érica Pacheco Caetano, Lucas Pereira de Alencar, Manoel Paiva de Araújo
Neto, Vitor Luz, Jamille Alencar Sales, Natalya Fechine Silva.
Todo grupo do CEMM e do PPGMM, que de alguma forma contribuíram com meu trabalho,
pelo companheirismo e incentivo que me proporcionaram.
Às minhas primas Maria Celeste Santos Vale, responsável por minha vinda a Fortaleza e por
ter aberto as portas da UFC, para mim e Antônia Maria Silva Figueiredo pelo apoio.
A todos os amigos funcionários da Faculdade de Medicina pelo companheirismo e apoio de
Agradeço todas as dificuldades que enfrentei;
Não fosse por elas, eu não teria saído do lugar.
Mesmo as críticas nos auxiliam muito.
RESUMO
Mudanças importantes na epidemiologia das infecções fúngicas nas últimas décadas têm
resultado no isolamento frequente de leveduras do gênero Candida. As leveduras do
complexo Candida pa rapsilosis, por exemplo, têm sido apontadas tanto como agentes de
infecções como componentes da microbiota de animais. Diante disso, os objetivos deste
trabalho foram realizar a identificação molecular de cepas do complexo C. parapsilosis,
isoladas de fontes veterinárias e mantidas no Centro Especializado em Micologia Médica;
assim como avaliar seus atributos de virulência e perfil de sensibilidade a antifúngicos, in
vitro. Para tanto, foram utilizados 28 cepas do complexo C. parapsilosis obtidos de cães,
psitacídeos, rapinantes e camarão. Inicialmente foi realizada a fenotipagem das cepas, com
base na análise de suas características morfológicas e bioquímicas, que as ratificou como C.
parapsilosis (lato sensu). A identificação molecular das espécies foi realizada por PCR-REA. A fim de analisar o perfil de sensibilidade das cepas, empregou-se o teste de microdiluição em
caldo com anfotericina B, itraconazol, fluconazol, voriconazol e caspofungina, segundo
metodologia padronizada pelo M27-A3. CLSI (2008). No tocante aos atributos de virulência,
a capacidade da produção de fosfolipases foi avaliada pelo método de cultivo em ágar gema
de ovo, a produção de proteases foi analisada através de cultivo em ágar albumina bovina e a
formação de biofilme foi em microplaca de poliestireno com 96 poços. A análise genotípica
evidenciou 13 C. parapsilosis (stricto sensu), 10 C. orthopsilosis e 05 C. metapsilosis. As
concentrações inibitórias mínimas (MICs) variaram de 0,125 a 1 μg/mL para anfotericina B, de 0,5 a 16 μg/mL para fluconazol, de 0,03125 a 0,5 μg/mL para itraconazol, de 0,03125 a 0,25 μg/mL para voriconazol e de 0,0625 a 2 μg/mL para caspofungina. Foi observada
resistência em 03 cepas de C. parapsilosis (stricto sensu) ao fluconazol e 01 cepa apresentou
MIC elevado (2 μg/mL) para caspofungina. No que tange aos isolados de C. orthopsilosis,
notou-se que 05 isolados apresentaram MICs elevados (2 μg/mL) para a caspofungina.
Enquanto que, 02 isolados de C. metapsilosis revelaram-se resistentes ao fluconazol. Quanto à
virulência, todas as cepas foram capazes de formar biofilmes, sendo, 20, 7 e 01, classificadas
como produtoras moderadas, fortes e fracas respectivamente. Observou-se, ainda que 23/28
isolados apresentaram atividade proteolítica. Por outro lado, nenhuma foi capaz de produzir
fosfolipases. Estes dados sinalizam que padrões de virulência, patogenicidade sensibilidade
antifúngica, in vitro, podem variar entre as espécies do complexo C. parapsilosis.
ABSTRACT
In the past decades, important changes in the epidemiology of fungal infections have resulted
in the frequent isolation of yeasts of the genus Candida. Yeasts of the Candida parapsilosis
species complex, for example, have been pointed out as infectious agents and components of
the microbiota of animals. Thus, the work aimed at identifying molecularly the strains of the
C. parapsilosis species complex recovered from veterinary sources and maintained at the
Specialized Medical Mycology Center, as well as evaluating their in vitro antifungal
susceptibility profile and attributes of virulence. For such, 28 strains of the C. parapsilosis
species complex, recovered from dogs, psittacines, raptors and prawn were assessed. Initially,
the strains were phenotypically identified, based on their morphological and biochemical
characteristics, which confirmed their identification as C. parapsilosis lato sensu. The
molecular identification of the strains was then carried out through PCR-REA. In order to
analyze the in vitro antifungal susceptibility, the broth microdilution assay was performed,
according to the document M27-A3 of the Clinical Laboratory Standards Institute (CLSI)
2008, using amphotericin B, itraconazole, voriconazole, fluconazole and caspofungin.
Concerning the virulence attributes, the ability of producing phospholipase was evaluated on
egg yolk agar, while protease production was assessed on bovine serum albumin agar, and
biofilm formation was tested in 96-well polystyrene microplates. The genotypical analysis
identified 13 C. parapsilosis stricto sensu, ten C. orthopsilosis and five C. metapsilosis. The
minimum inhibitory concentrations varied from 0.125 to 1 μg/mL foramphotericin B, from
0.03125 to 0.5 μg/mL, for itraconazole, from 0.03125 to 0,25 μg/mL for voriconazole, from
0.5 to 16 μg/mL for fluconazol and from 0.0625 to 2 μg/mL for caspofungin. Fluconazole
resistance was observed in three strains of C. parapsilosis stricto sensu and two of C.
metapsilosis, while one strain of C. parapsilosis stricto sensu and five C. orthopsilosis
presented high MIC values for caspofungin (2 μg/mL). As for the virulence attributes, none of
the tested strains produced phospholipases, while 23/28 presented proteolytic activity, and all
of the strains produced biofilm, with one weak producer, 20 moderate producers and seven
strong producers. These data show that the in vitro antifungal susceptibility and production of
virulence attributes vary among species of the C. parapsilosis species complex, which can
lead to differences in pathogenicity and therapeutic response.
LISTA DE FIGURAS
Figura 1 - Características macro e micromorfológicas de Candida spp. A. Colônias em meio Sabouraud dextrose. B. Blastoconídios, preparação com lacto fenol azul algodão ... 17
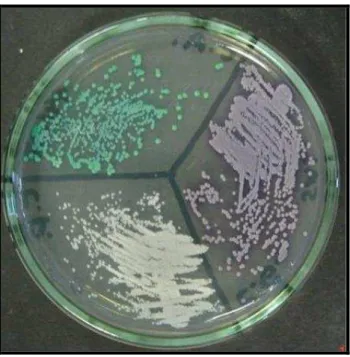
Figura 2 – CHROM ágar: Candida parapsilosis colônias brancas e colônias lilás ... 18
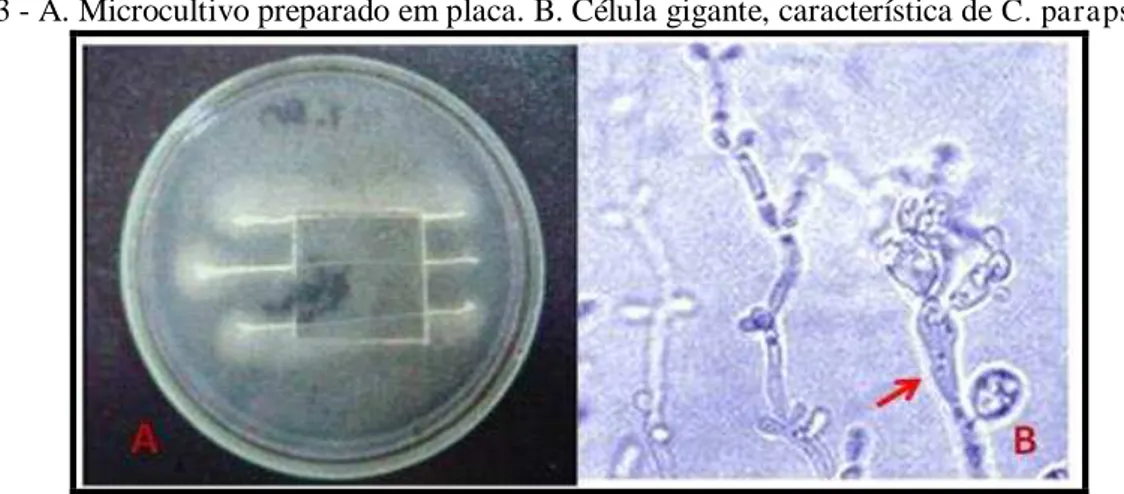
Figura 3 - A. Microcultivo preparado em placa. B. Célula gigante, característica de C. parapsilosis... 19
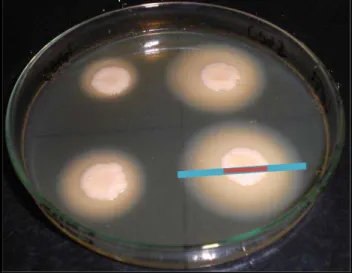
Figura 4 - Prova de produção da enzima uréase. Tubo da esquerda representa um teste positivo, apresentando alteração de coloração (rosa), devido à presença da enzima no meio. Tubo da direita representa um teste negativo, característico de C. parapsilosis lato sensu, permanecendo com a coloração original do meio (amarelo) ... 20
Figura 5 - Prova de assimilação de carboidrato. As setas indicam assimilação de diferentes açúcares evidenciada pela a formação de halo onde ocorreu o crescimento fúngico ... 21
Figura 6 - Prova de assimilação de nitrogênio. A seta indica assimilação de peptona evidenciada pela formação de halo onde ocorreu o crescimento fúngico ... 22
Figura 7 - Teste de fermentação de carboidratos, a seta indica a formação de gás no interior do tubo de Durhan ... 23
SUMÁRIO
1. INTRODUÇÃO ...11
2. REVISÃO DE LITERATURA ...12
2.1. Aspectos gerais do gênero Candida ...12
2.2. O gênero Candida e sua correlação com o homem e animais...13
2.3. Complexo Candida parapsilosis ...15
2.4. Características fenotípicas de Candida parapsilosis lato sensu ...17
2.5. Identificação molecular das espécies do complexo Candida parapsilosis ...23
2.6. Atributos de virulência associados à Candida spp. ...24
3. PERGUNTAS DE PARTIDA ...28
4. HIPÓTESES ...28
5. OBJETIVOS ...29
5.1 Objetivo Geral ...29
5.2 Objetivos específicos ...29
6. CAPÍTULO 1 ...30
8. PERSPECTIVAS ...34
11
1. INTRODUÇÃO
As leveduras do gênero Candida são consideradas espécies saprófitas ou comensais
da microbiota associada à pele e às mucosas de homens e de outros animais. Esses
microrganismos, na maioria das vezes, não conferem danos aos seus hospedeiros, mas diante
de distúrbios nas barreiras físicas ou imunológicas, podem se tornar patogênicos e causar
infecções superficiais ou profundas. Na América Latina, as três principais espécies envolvidas
em quadros clínicos humanos são C. albicans, C. tropicalis e C. parapsilosis. No entanto
Candida parapsilosis, anteriormente, foi considerada essencialmente saprófita e não
patogênica, até o ano de 1940, quando foi implicado como agente etiológico de um caso fatal
de endocardite. A partir de então, C. parapsilosis tem sido considerada agente etiológico de
candidíases em homens e animais, causando infecções de origem exógena.
Em meados da década de 1990, observou-se a existência de três grupos genéticos
distintos na espécie C. parapsilosis, os quais foram posteriormente reclassificados em três
espécies distintas, designadas como C. parapsilosis (stricto sensu), C. orthopsilosis e C.
metapsilosis. Tais espécies estão atualmente agrupadas no Complexo Candida parapsilosis. No entanto, em humanos, ainda são poucos os relatos que tratam das características desse
complexo. Sendo praticamente inexistentes estudos sobre as espécies do Complexo C.
parapsilosis em animais.
Semelhantemente a outras espécies do gênero Candida, os membros do Complexo C.
parapsilosis apresentam como principais fatores de virulência a secreção de enzimas
hidrolíticas, como proteases e fosfolipases. Além disso, há a capacidade de formação de
biofilme. Quanto à sensibilidade antifúngica, diversos antifúngicos apresentam atividades
contra as espécies do complexo C. parapsilosis, com destaque para os derivados azólicos, os
poliênicos e as equinocadinas. Cepas desta espécie são menos sensíveis a caspofungina,
devido a uma mutação genética que pode conferir tal tolerância. Deste modo, tornam-se
necessários estudos que avaliem o perfil de sensibilidade, para o monitoramento de possíveis
resistências a estes antifúngicos.
Diante do exposto, os objetivos deste trabalho foram realizar a identificação
molecular de cepas do complexo C. parapsilosis isoladas de fontes veterinárias e mantidas na
micoteca do Centro Especializado em Micologia Médica (CEMM), da Universidade Federal
do Ceará (UFC), assim como avaliar seus atributos de virulência e perfil de sensibilidade a
12
2. REVISÃO DE LITERATURA 2.1. Aspectos gerais do gênero Candida
Desde meados da década de 1980, os fungos têm reemergido como um dos principais
microrganismos responsáveis por infecções em humanos, principalmente, em indivíduos
imunocomprometidos e pacientes hospitalizados com doenças subjacentes graves (SIDRIM:
ROCHA, 2004; TROFA; GACSER; NOSANCHUKI, 2008). Espécies de Candida são a
segunda principal causa de infecção da corrente sanguínea nosocomial. Sendo citadas como
os microrganismos responsáveis por 8 a 15% de todos os casos de infecções hospitalares nos
Estados Unidos. O gênero Candida, atualmente, é responsável por cerca de 80% das infecções
fúngicas em hospitais (MURRAY et al., 2006), resultando em quadros de morbidade e
mortalidade substanciais, ocasionando impacto significativo na saúde pública (TROFA; GACSER; NOSANCHUKI, 2008).
O gênero Candida pertence ao Reino Fungi, Filo Ascomycota, Classe
Saccharomycetes, Ordem Sa ccharomycetales. As leveduras desse gênero são consideradas
espécies saprófitas ou comensais da microbiota associada à pele e mucosas do homem e
outros animais, tais como cães, gatos, ruminantes e aves (SIDRIM; ROCHA, 2004;
BENTUBO et al., 2007). Esse gênero compreende mais de 200 espécies de organismos
leveduriformes, unicelulares, anamórficos com mecanismo de reprodução assexuada que
inclui brotamento simples, brotamento fissão e divisão binária (BRITO 2005). A maioria dos
organismos desse gênero é capaz de exibir pleomorfismo – formando simultaneamente
blastoconídios e estruturas filamentosas, em condições ambientais adequadas, independente
da temperatura (SIDRIM; ROCHA, 2004).
Geralmente, esses organismos não causam nenhum dano aos seus hospedeiros, mas
frente à presença de distúrbios nas barreiras física ou imunológica, podem se tornar
patogênicos e causar infecções superficiais ou profundas. Porém, infecções de pele e mucosas podem acontecer em indivíduos saudáveis (SIDRIM; ROCHA, 2004; BRITO 2008).
As espécies do gênero Candida são os patógenos fúngicos oportunistas mais
comumente encontrados. Algumas características desses organismos contribuem para sua
patogenicidade como, por exemplo, o dimorfismo entre as formas de levedura e hifa, a
hidrofobicidade de sua superfície celular, produção de tubo germinativo, mudanças
fenotípicas, capacidade de adesão a diversos tecidos e superfícies inanimadas, assim como a
secreção de enzimas proteolíticas, sendo as proteases e as fosfolipases as de maior relevância
13
Infecções em animais causados por Candida spp. são pouco frequentes, no entanto,
nos últimos anos houve aumento considerável de infecções como: dermatomicoses, otites,
infecção urinária, entre outros, causadas principalmente pela espécie C. albicans (BRITO et
al.,2009; MAIA,2009).
Mudanças importantes na epidemiologia das infecções fúngicas nas últimas décadas
têm resultado no isolamento de outras espécies do gênero, tais como: C. parapsilosis, C.
tropicalis, C. krusei, C. glabratae C. famata, comumente citadas como “espécies de Candida
não-albicans”. Dados da literatura sugerem que as infecções por esses organismos são mais
comuns em pacientes com imunossupressão grave (COSTA et al., 2009). Atualmente sabe-se
que esses organismos podem causar diversas enfermidades em hospedeiros suscetíveis, tais
como otite, endoftalmite, infecção intestinal e urinária (CAFARCHIA et. al., 2006).
Candida parapsilosis (lato sensu) se destaca entre as espécies de Candida
não-albicans, devido aos relatos de altas taxas de morbidade e de mortalidade relacionadas ao complexo C. parapsilosis. Torna-se relevante a busca de uma terapia mais eficaz, além de
estudos mais intensos para identificar características genéticas, como a expressão de fatores
de virulência (TROFA; GACSER; NOSANCHUKI, 2008).
Originalmente descrita por Ashford em 1928 como Monilia parapsilosis - levedura
não fermentadora de maltose isolada de amostras de fezes de um paciente com diarréia em
Porto Rico - somente em 1932 a espécie foi reclassificada como Candida parapsilosis
(SIDRIM; ROCHA, 2004; TROFA; GACSER; NOSANCHUKI, 2008; NOSEK et al., 2009).
O microrganismo foi considerado essencialmente saprófita e não patogênico até o
ano de 1940, quando foi implicado como agente etiológico de caso fatal de endocardite. A
partir de então, C. parapsilosis tem sido considerado agente etiológico de candidíases em
humanos e outros animais, sendo essas infecções comumente de origem exógena O
microrganismo pode ser isolado de fontes ambientais, como ar, solo, água salobra ou salina
(TROFA; GACSER; NOSANCHUKI, 2008; NOSEK et. al., 2009).
2.2. O gênero Candida e sua correlação com o homem e animais
A facilidade de adaptação e disseminação dessa levedura relaciona-se com a
diversidade de infecções em que configura como o único agente etiológico de fungemias, em
seres humanos, tais como: endocardite, endolftalmite, artrite séptica, peritonite, vaginite e
onicomicose. Durante a última década episódios envolvendo C. parapsilosis têm aumentado
14
mais isolada em culturas de sangue de pacientes hospitalizados. Entretanto, em alguns países
da Europa, Ásia e da América do Sul, o número de isolados supera o de C. albicans (TROFA;
GACSER; NOSANCHUKI, 2008).
Diversos estudos mostram o aumento da incidência de infecções por C. parapsilosis
(FALAGAS et al., 2010). No Estado do Ceará, estudos retrospectivos mostraram que esta
espécie é agente etiológico frequente em quadros de candidemia e onicomicose
(BRILHANTE et al., 2005; MEDRANO et. al., 2006). Ao investigar a presença de fungos no
ar, em áreas críticas e não críticas, de dois hospitais terciários em Fortaleza – Ceará, Pantoja
et al. (2009), relataram que C. parapsilosis foi a espécie mais frequentemente isolada entre as
espécies de Candida, salientando-se a emergência do microrganismo como agente de
infecções hospitalares em nossa região. Recentemente, foi descrito caso de meningite por C.
parapsilosis, como manifestação oportunista em um paciente com AIDS (SIDRIM et. al., 2011).
Dentre as espécies do gênero Candida isoladas de animais, destaca-se Candida
parapsilosis, como importante agente de infecções, bem como, componente da microbiota ((SANTOS; MARIN, 2005; BRITO et al., 2009; BRILHANTE et al., 2010). A importância
dessa levedura como comensal da microbiota de animais domésticos e selvagens vem sendo
investigada pelo Centro Especializado em Micologia Médica da Universidade Federal do
Ceará. Em estudo sobre a distribuição de leveduras em cães saudáveis, Brito et al., (2008),
realizaram a coleta de swabs das mucosas perianais, prepúcio, vagina e cavidade oral em 203
cães. Segundo os autores, C. parapsilosis foi o segundo microrganismo mais comumente
isolado desses animai. Outro estudo sobre a microbiota de Psitacídeos (Nymphicus
hollandicus), os autores relataram que C. para psilosis faz parte da microbiota do trato
gastrintestinal dessas aves (BRITO et al., 2008; ALENCAR, 2012; BRILHANTE et al.,
2010).
As candidíases em aves são frequentemente responsáveis pela mortalidade de
animais mantidos em cativeiro. Uma vez que a microbiota bacteriana do trato gastrintestinal
das aves exerce importante efeito regulador da população de Candida spp., qualquer
desequilíbrio da microbiota digestiva, pelo uso incorreto de antibióticos ou por mudanças no
pH gastrintestinal, pode resultar na proliferação de fungos (MAIA, 2009; BRILHANTE et al.,
2013). A micose no inglúvio desenvolve-se como uma infecção primária ou secundária, uma
vez que este sítio anatômico é suscetível a infecções por espécies de Candida, por ser um
órgão saculiforme, com nutrientes necessários para o crescimento dessas espécies (KANO et
15
Tsai et al. (1992) relataram incidência de 15,4% de candidíase em 241 psitaciformes
e passeriformes que foram a óbito. No entanto, foi constatado que a região vestibular da
cavidade nasal foi, frequentemente, acometida por levedura, constatando-se a importância
dessa região anatômica para entrada e multiplicação de Candida spp. (MAIA, 2009). Kano et
al. (2001) diagnosticaram, através de exames histopatológicos e imuno-histoquímico, a
ocorrência de ingluvite, em um psitacídeo de dois meses, associada a Candida spp. Onde a
espécie envolvida foi identificada como Candida parapsilosis por técnicas moleculares.
Vieira e Acqua-Coutinho (2009) observaram a ocorrência também de C. parapsilosis, em
associação com outras espécies de Candida, causando infecção no inglúvio de filhotes de
papagaios (KANO et al. 2001; MAIA, 2009). No início da década de 70, Cragg e Clayton
evidenciaram leveduras em cultura de fezes frescas de gaivotas (Larusoccidentalis), onde a
espécie de C. parapsilosis foi a mais prevalente. Pesquisando microbiota de emas (Rhea
americana), Brilhante et al. (2013) relataram que C. parapsilosis foi a espécie com maior frequência de isolamento em relação a outras Candida spp. (ALENCAR, 2012; BRILHANTE
et al., 2013). Em outro trabalho, realizado com equinos Cordeiro et al. (2013), relataram o
isolamento de C. parapsilosis como membro da microbiota nasal desses animais Em bovinos,
as candidíases constituem a terceira maior causa infecciosa de mastites no Brasil. Em outros
países, embora a incidência dessa infecção ainda seja pequena, o aumento de sua frequência
tem sido relatado (KIVARIA; NOORDHUIZEN, 2007; SANTOS; MARIN, 2005).
2.3. Complexo Candida parapsilosis
A existência de grupos com diversidade genética distinta entre os isolados de Candida
parapsilosis lato sensu foi sugerido em 1990. Em estudo pioneiro no ano de 1992, por meio de técnica de Reação em Cadeia de Polimerase e Amplificação Aleatória de DNA Polimórfico
- PCR-RAPD, foi constatado que os oligonucleotídeos RP2 e RP4-2, eram capazes de
diferenciar cepas de Candida parapsilosis, fisiologicamente homogêneas, em três grupos
geneticamente distintos (LEHMAN; LASKER, 1992).
A partir de então diversos estudos, baseados em técnicas moleculares, comprovaram a
existência dessa diversidade genética. Eletroforese de enzimas codificadas por multilocus,
bem como sequenciamento de genes ribossomais e mitocondriais (NOSEK et. al., 2009; LIN,
et al. 1995), Em 2005, Tavanti et al., em estudos visando a caracterização molecular de
espécies patogênicas do gênero Candida por meio da técnica de "Multilocus Sequence
16
parapsilosis, que foram classificados como: 21 isolados pertencente ao grupo I, nove no
grupo II e dois isolados no III. Com base na reação de RAPD, com os oligonucleotídeos
RPO2 e sequenciamento das regiões ITS1, ITS2, -5.8S, do rRNA, os autores amplificaram o
gene da Desidrogenase Alcóolica Secundária SADH (Seconda ry Alcohol Dehydrogena se).
Baseados nesses dados e na diferença existente entre os três grupos, também relatadas por
outros autores. Tavanti e colaboradores (2005) propuseram a seguinte denominação
taxonômica para os três grupos, Candida parapsilosis (stricto sensu), Candida orthopsilosis e
Candida metapsilosis, para os isolados pertencentes aos grupos, I, II, e III, respectivamente
(TAVANTI et al 2005; TAVANTI et al., 2007; LOCKHART et al., 2008; TOSUN et al.,
2013).
Alguns estudos têm mostrado dados epidemiológicos sobre as espécies do Complexo
C. parapsilosis, sendo todos relativos ao isolamento em humanos. No entanto, em 2007, foi publicada a primeira descrição detalhada sobre a participação desses organismos em infecções
humanas, na qual foi confirmada a participação de C. orthopsilosis em 4,5% dos casos
estudados. Em outro estudo realizado, foi observado que 6,1% dos isolados foram
identificados como C. orthopsilosis e 1,8% como C. metapsilosis. De acordo com os autores,
embora exista grande variação na frequência de isolamento desses organismos em cada
região, C. orthopsilosis, parece ter maior prevalência na América do Sul (TAVANTI et al.,
2007; LOCKHART et al., 2008).
No Brasil, estudos relatam a interferência das espécies do Complexo C. parapsilosis,
também em humanos. Em 2010, Gonçalves e colaboradores investigaram a frequência do
isolamento desses microrganismos em 141 amostras de hemocultura humana e relataram a
presença de C. parapsilosis (stricto sensu) (88%), C. orthopsilosis (9%) e C. metapsilosis
(3%). Em estudo realizado com isolados de C. parapsilosis (lato sensu) em crianças
hospitalizadas, em um hospital do estado de São Paulo, a análise molecular confirmou das 49
cepas analisadas (83,7%), como Candida parapsilosis stricto sensu, seguida por C.
orthopsilosis (10,2%),e C. metapsilosis (6,1%) (GONÇALVES et al., 2010; RUIZ et al., 2013).
Vale ressaltar que, até o momento, não há nenhum estudo sobre o isolamento de
espécies do Complexo C. parapsilosis em animais, apesar do aumento da importância das
candidíase em pequenos animais. Com isso, não há informações sobre as características
fenotípicas e moleculares, bem como, o perfil de sensibilidade a drogas antifúngicas contra
17
relacionadas com a patogenicidade, resistência antifúngica e caráter epidemiológico das
espécies de Candida inseridas nesse complexo.
2.4. Características fenotípicas de Candida parapsilosis lato sensu
A identificação das espécies de Candida tem significado importante na terapêutica
das doenças causadas por essas espécies, principalmente em casos de doenças sistêmicas, visto que essa identificação prévia pode antecipar o início do tratamento adequado. As
espécies pertencentes ao gênero Candida entre elas C. parapsilosis, possuem características
morfológicas peculiares que podem ser identificadas por métodos fenotípicos convencionais.
No entanto, as espécies do Complexo C. parapsilosis, até o momento somente são
identificadas por técnicas moleculares como PCR (SIDRIM; ROCHA, 2004; TAVANTI et
al., 2007).
A microscopia direta do material clínico preparado com KOH (10 a 40%) consiste no
diagnóstico inicial, onde observam-se estruturas como blastoconídios, associadas ou não a
pseudo-hifas e pseudomicélios, de forma que as diversas espécies não podem ser
diferenciadas somente por esta técnica. No entanto, apenas a análise microscópica não
permite a identificação. Para a análise macroscópica, as amostras clínicas precisam ser
cultivadas em meio de culturas, específicos ao isolamento primário, Agar Sabouraud Dextrose
a 2%, onde as espécies de Candida apresentam colônias de coloração branca ou creme com
textura glabrosa úmida, superfície lisa convexa. Com isso faz-se necessário, a união entre análises macro e micromorfológica das colônias (Figura 1) para identificação das
características morfológicas dessas espécies (SIDRIM; ROCHA, 2004: BRITO, 2005;
KONEMAN et al., 2008).
Figura 1 - Características macro e micromorfológicas de Candida spp. A. Colônias em meio Sabouraud dextrose. B. Blastoconídios, preparação com lacto fenol azul algodão
(Fonte: CEMM, 2013)
18
CHROMagar Candida, meio de cultura cromogênico diferencial, é incubado à
temperatura ambiente (25 – 30ºC) por até 72hs, permitindo a avaliação da morfologia das
colônias através dos padrões distintos de coloração, sendo utilizado para separar as espécies
de Candida em culturas de leveduras, sobretudo, quando observadas em culturas mistas. A
espécie de Candida pa rapsilosis apresenta coloração lilás e, algumas vezes, branca (Figura 2).
No entanto, este meio não é suficiente para um diagnóstico seguro, devido às interpretações
subjetivas, sendo útil somente para auxiliar na identificação da morfologia (DE HOOG et al.,
2000; SIDRIM; ROCHA, 2004; KONEMAN et al., 2008).
Figura 2 – CHROM ágar: Candida parapsilosis colônias brancas e colônias lilás
(Fonte: CEMM, 2013)
Posteriormente, são realizados testes micromorfológicos, com microcultivos em
placa utilizando ágar arroz ou ágar fubá com Tween 80 (Figura 3A). A avaliação permite a
identificação da micromorfologia, uma vez que formam estruturas específicas, como hifas e pseudo-hifas, dispostas em padrões característicos para cada espécie. Dessa forma, C.
19
Figura 3 - A. Microcultivo preparado em placa. B. Célula gigante, característica de C. parapsilosis
(Fonte: CEMM, 2008).
A formação de tubos germinativos ou hifas verdadeiras por Candida albicans tem
sido sugerido como importante fator de virulência, pois as hifas têm maior capacidade de
aderir e penetrar nas células epiteliais humanas de que os blastoconídios. Até então somente
C. albicans e C. dubliniensis formam tubo germinativo, uma projeção alongada que emerge do blastoconídio. Para isso, precisam ser postas em contato com soro humano ou de animais,
em temperatura de 37°C, por um período de duas a três horas (DE HOOG et al., 2000;
MORETTI et al., 2000; SIDRIM; ROCHA, 2004; BRITO, 2005; ÁLVARES et al., 2007).
Quanto às provas bioquímicas, Candida spp., são submetidas à prova da uréase, realizada em meio de cultura Ágar Ureia de Christensen, que permite a detecção colorimétrica
da enzima e determina a qual divisão pertence o microrganismo. Este teste avalia se a
levedura produz ou não a enzima uréase. Um fragmento da colônia fúngica é repicado para o
meio com indicador de pH, incubado em estufa a 37°C durante duas a quatro horas. Quando,
positivo a enzima uréase entra em contato com o meio e hidrolisa a ureia com liberação de
amônia, gerando uma mudança no pH observada pela coloração róseo intensa. As espécies de
Candida em geral não produzem uréase, (Figura 4) (DE HOOG et al., 2000; SIDRIM;
20
Figura 4 - Prova de produção da enzima uréase. Tubo da esquerda representa um teste positivo, apresentando alteração de coloração (rosa), devido à presença da enzima no meio. Tubo da direita representa um teste negativo, característico de C. parapsilosis lato sensu, permanecendo com a coloração original do meio (amarelo)
(Fonte: CEMM, 2013).
A assimilação de carboidratos também denominada de auxograma consiste na
utilização de um carboidrato como única fonte de carbono utilizada pela levedura. Cada
espécie do gênero Candida apresenta um perfil de assimilação de carboidratos diferente e a
leitura é feita, durante o tempo de 24 a 96 horas, mediante visualização de halos de
crescimento que se formam em volta do carboidrato assimilado (Figura 5). Os carboidratos testados são: inulina, ramnose, L-arabinose, celobiose, dextrose, sacarose, rafinose, dulcitol,
melibiose, trealose, galactose, maltose, xilose, inositol e lactose. C. parapsilosis assimila os
carboidratos: arabinose, dextrose, galactose, sacarose, trealose e xilose. (DE HOOG et al.,
21
Figura 5 - Prova de assimilação de carboidrato. As setas indicam assimilação de diferentes açúcares evidenciada pela a formação de halo onde ocorreu o crescimento fúngico
(Fonte: CEMM, 2009).
Na assimilação de nitrogênio as espécies de Candida podem assimilar ou não
nitrogênio, de forma que este teste avalia a capacidade que a levedura tem de crescer
aerobicamente na presença de um composto nitrogenado, fornecido como única fonte
energética. Para isso, em meio sólido, acrescido da suspensão fúngica e destituído de qualquer
fonte de nitrogênio, são colocadas alíquotas de peptona, utilizada como controle positivo, e
nitrato de potássio. Quando há formação de um halo, após 96 horas de incubação a 30ºC, em
volta do nitrato de potássio o teste é considerado positivo (Figura 6). Geralmente as espécies
de Candida, isoladas de amostras clinicas, assimilam peptona por esta ser uma fonte orgânica
de nitrogênio, a capacidade de assimilar ou não nitrato de potássio como fonte de nitrogênio
inorgânica, varia para cada espécie (SIDRIM; ROCHA, 2004; BRITO et al., 2009). No
22
Figura 6 - Prova de assimilação de nitrogênio. A seta indica assimilação de peptona evidenciada pela formação de halo onde ocorreu o crescimento fúngico
(Fonte: CEMM, 2013).
Na fermentação, o carboidrato é utilizado como única fonte de energia onde avalia-se
a capacidade da levedura em questão utilizar esse carboidrato e crescer anaerobicamente na
presença do mesmo. A levedura é suspensa em solução salina a 0,9% (Escala de McFarland5),
depois colocada em um tubo de ensaio contendo meio de cultura liquido e um tubo de
Durhanem posição invertida. A positividade se dá pela produção de gás carbônico, no interior
do tubo de Durhan, formando bolhas visíveis. Na presença de um indicador de pH, haverá
mudança de coloração do meio, em virtude da acidificação. Geralmente, os carboidratos
utilizados neste teste são: dextrose, maltose, sacarose, galactose, lactose e trealose, além de
outros. O carboidrato fermentado, também é obrigatoriamente assimilado pela mesma cepa,
no entanto, o inverso não é verdadeiro (Figura 7). A espécie C. parapsilosis fermenta obrigatoriamente dextrose e varia para os demais carboidratos testados (DE HOOG et al.,
23
Figura 7 - Teste de fermentação de carboidratos, a seta indica a formação de gás no interior do tubo de Durhan
(Fonte: CEMM, 2010).
Quanto à sensibilidade antifúngica, as três espécies do complexo C. parapsilosis são
geralmente sensíveis aos azólicos e poliênios, mas a sensibilidade às equinocandinas ainda é
incerta. Em particular, dados sobre sensibilidade in vitro às equinocandinas, como
caspofungina, indicam que C. parapsilosislato sensu está entre as espécies menos sensíveis do
gênero Candida. Existem relatos de que as concentrações inibitórias mínimas de caspofungina
ante a C. orthopsilosis e C. metapsilosis são menores que aquelas ante C. parapsilosis stricto
sensu. Entretanto, há uma escassez de dados in vivo sobre os efeitos desse composto contra as espécies do complexo C. parapsilosis. (SPREGHINI et al., 2012).
2.5. Identificação molecular das espécies do complexo Candida parapsilosis
A existência de grande diversidade genética entre os isolados clínicos de C.
parapsilosis tem sido mostrada em diversos estudos. Análises baseadas em técnicas moleculares, tais como Amplificação Aleatória de DNA Polimórfico – RAPD (LEHMAN;
LASKER, 1992), eletroforese de enzimas codificadas por multilocus, bem como
sequenciamento de genes ribossomais e mitocondriais (NOSEK et al., 2002), comprovaram a
existência de três grupos genéticos distintos nesta espécie, denominados grupos I, II e III.
Posteriormente, Tavanti et al. (2005) propuseram a reclassificação dos grupos em três
espécies distintas, designadas como C. parapsilosis (stricto sensu), C. orthopsilosis e C.
metapsilosis para os isolados pertencentes aos grupos I, II e III, respectivamente. Tais
espécies estão agrupadas no Complexo Candida parapsilosis (TAVANTI et al., 2005).
A grande maioria dos isolados clínicos pertence à espécie C. pa rapsilosis (stricto
24
desenvolvimento de técnicas de genotipagem. Assim como outras espécies do gênero
Candida, os organismos do Complexo C. parapsilosis apresentam como principais fatores de
virulência a secreção de enzimas hidrolíticas, tais como, proteases, fosfolipases, bem como a
capacidade de aderência a superfícies inanimadas e formação de biofilme (TORO et al.,
2011).
As cepas específicas de microrganismos podem ser distinguidas com base no seu
DNA ou RNA, ou fragmentos de DNA produzidos quando este DNA é clivado por
endonucleases de restrição específicas (enzimas de restrição), as mesmas reconhecem
sequencias específicas de DNA que possuem uma estrutura polidrômica, onde os sítios de
DNA reconhecidos por diferentes endonucleases de restrição diferem em sua sequência,
comprimento e frequência de ocorrência, como resultado, diferentes endonucleases clivam o
DNA de uma mesma amostra em diferentes regiões, resultando em fragmentos de
comprimentos distintos, onde essa peculiaridade em diferentes cepas de um mesmo
organismo é classificada de polimorfismo do comprimento do fragmento de restrição (RFLP-
do inglês Restriction Fragmente Length - Polymorphism). Esses fragmentos movem-se
através de uma estrutura labiríntica em gel de agarose ou acrilamida em velocidades
diferentes que permitem sua separação. O DNA pode ser corado com brometo de etídio e
visualizado em um transiluminador (KONEMAN et al., 2008).
2.6. Atributos de virulência associados à Candida spp.
A capacidade da levedura em aderir, infectar e causar doença em conjunto é definida
como potencial de virulência ou patogenicidade. Os fatores de virulência são determinados
geneticamente e são expressos pelos microrganismos quando estes são submetidos à condições
adversas.
Os biofilmes representam o tipo de crescimento microbiano predominante na
natureza e são cruciais ao desenvolvimento de infecções, uma vez que servem de nicho aos
agentes patogênicos e estão associados a altos níveis de resistência a agentes antimicrobianos
(TAMURA et al., 2007). As infecções associadas à utilização de implantes médicos invasivos
estão relacionadas com a formação de biofilmes nesses dispositivos. O desenvolvimento do
biofilme depende do tipo e do número de células que aderem ao dispositivo, do tipo de
superfície que constitui o dispositivo. A formação do biofilme inicia-se com a adesão
microbiana seguida pela fase de maturação (TAMURA et al., 2007). Constitui um forte fator
25
antifúngica controlando a penetração das substâncias através da matriz das células de defesa
do hospedeiro. Segundo relatos a formação de biofilme por espécies de Candida, incluindo
Candida parapsilosis, têm sido associada às taxas de mortalidades significativas, onde
isolados de C. parapsilosis com capacidade para formar biofilme, in vitro, elevaram as taxas
de mortalidade em 71,4% em relação às cepas deficientes (não formadoras de biofilme) que
foi de 28%. (TROFA; GACSER; NOSANCHUKI, 2008). Os dispositivos contaminados por
C. parapsilosis precisam ser removidos para desinfecção. No entanto há relatos de
tratamentos bem sucedidos com terapia antifúngica na formação de biofilme em infecções
isoladas. (TROFA; GACSER; NOSANCHUKI, 2008)
As fosfolipases são enzimas extracelulares relacionadas a processos patogênicos
oportunistas causados por leveduras. São implicadas como fatores de virulência de vários
organismos, como por exemplo: Candida albicans, que produz fosfolipase B, codificada pelos
genes PLB1 e PLB2. Nota-se assim que o tipo de fosfolipase varia de acordo com o
organismo. (GHANNOUM, 2000; CASTELO BRANCO, 2009). Por vários anos,
acreditou-se que somente Candida albicans, produzia fosfolipases. No entanto, em (1998) alguns
autores relataram a produção da enzima por outras espécies de Candida spp., com uma
capacidade muito menor que C. albicans (GHANNOUM 2000; CASTELO BRANCO, 2009).
A importância dessa enzima na virulência de C. parapsilosis, não está bem esclarecida, são
contraditórios os relatos, enquanto alguns autores relataram resultados positivos em 51% dos
casos, outros chegaram a relatar nenhum caso de positividade (TROFA; GACSER;
NOSANCHUKI, 2008). No entanto a produção de fosfolipase por leveduras de origem aviária
são poucos, porém, Cafarchia et al. (2008) avaliaram a capacidade de produção de fosfolipase
em leveduras isoladas de aves migratórias, de passeriforme, fezes e cloaca de rapinantes,
colhidas dos recintos. Das leveduras isoladas da cloaca e das fezes, 48,1% e 73,3%
apresentavam produção de fosfolipase, respectivamente. No geral, os isolados obtidos da
cloaca apresentaram produção de fosfolipase estatisticamente inferior àquela das leveduras
obtidas por meio da cultura de fezes (CAFARCHIA et al., 2008; BRILHANTE et al., 2013)
Price et al. (1982) descreveram uma metodologia para determinação da fosfolipase
em C. albicans, onde utilizou-se cultivos de cepas em placas contendo meio ágar Sabouraud
dextrose, acrescido de gema de ovo, uma rica fonte e fosfolipídios. Este é um método bastante
factível, que tornou-se clássico para triagem da produção de fosfolipases por Candida spp.,
sendo bastante citado na literatura. Quando positivos, há formação de uma zona de
precipitação densa, ao redor da colônia, devido à formação de complexos de cálcio com os
26
(CAFARCHIA et al., 2008; ZENG et al., 2008; COSTA et al., 2009; VIEIRA; COUTINHO,
2009). A atividade de fosfolipase (Pz) é interpretada pela razão entre o diâmetro da colônia e
o diâmetro do halo formado pela colônia e zona de precipitação (figura 8). Quando Pz for
igual a um (Pz = 1), o resultado é negativo. Quando Pz é menor que um, (Pz<1) atividade enzimática positiva, e Pz≥0,64 baixa atividade e Pz< 0,64 alta atividade enzimática (GHANNOUM, 2000; VIEIRA; COUTINHO, 2009). No entanto, não é uma técnica
adequada para testar cepas com baixa produção enzimática, sendo necessária a confirmação
por métodos mais sensíveis e específicos (COSTA et al., 2009 ALENCAR, 2012).
As proteinases são enzimas que degradam vários substratos, tais como queratina,
colágeno, albumina, fibronectina e hemoglobina. Estas enzimas possuem habilidades para
invadir as células destruindo tecidos e são classificadas de acordo com seu mecanismo
catalítico em quatro classes sendo estas: serinas, cisteínas, aspárticas e metaloproteinases.
Algumas espécies de leveduras expressão proteinases aspárticas. A aspartil proteinase (SAP),
é secretada por C. albicans, entretanto, genes homólogos já foram identificados em outras
espécies de Candida, como C. parapsilosis por exemplo. A aspartil proteinase é secretada por
uma família de dez genes, com um sistema proteolítico importante para o comensalismo e o
desenvolvimento de infecções por Candida spp. Alguns autores relatam que os genes (SAP1 a
SAP3) são expressos na fase leveduriforme e (SAP4 a SAP6) na filamentosa e que os
mesmos, parecem ter importância no desenvolvimento de infecções de mucosas e sistêmicas
por espécies de Candida. No entanto, Candida albicans tem maior expressão do gene SAP2
in vitro. Os genes SAP, são regulados por mudanças no ambiente na transição entre a forma,
leveduras e hifas, e diante de fenótipos alternativos (NAGLIK et al., 2004; SCHALLER et
al.,2005; COSTA, 2006; TROFA; GACSER; NOSANCHUKI, 2008).
As proteases secretadas por Candida spp. hidrolisam o colágeno, a queratina e a
mucina, assim como degradam anticorpos e citocinas. Alguns autores relatam que a secreção
de proteinase por Candida spp têm papel importante na patogenicidade dessas espécies. Essas
proteinases têm por função hidrolisar as proteínas do hospedeiro envolvidas nos mecanismos
de defesa, permitindo assim a penetração da levedura no tecido conjuntivo. A secreção de
proteases contribui para a aderência dos blastoconídios às células do hospedeiro, facilitando
assim a invasão dos tecidos pelas hifas através da degradação proteolítica da queratina e do
colágeno. Vários estudos apontam uma forte relação entre a secreção de protease e o grau de
27
Figura 8 - Produção de fosfolipase (Pz) em ágar gema de ovo. Pz representa a razão entre o diâmetro da colônia (linha vermelha) e o diâmetro total (colônia+zona de precipitação, linha azul)
28
3. PERGUNTAS DE PARTIDA
1. Qual a proporção das diferentes espécies do complexo C. parapsilosis em animais
saudáveis?
2. Qual o perfil de sensibilidade antifúngica desses isolados?
3. Estas leveduras apresentam diferenças na produção de atributos de virulência in vitro?
4. HIPÓTESES
1. C. parapsilosis stricto sensu corresponde a espécie mais frequente do Complexo Candida
parapsilosis em animais saudáveis;
2. Os organismos do Complexo Candida parapsilosis isolados da microbiota de animais
saudáveis apresentam diferentes perfis de sensibilidade a drogas antifúngicas;
3. Os organismos do Complexo Candida parapsilosis isolados da microbiota de animais
29
5. OBJETIVOS
5.1 Objetivo Geral
O objetivo deste trabalho foi realizar a identificação molecular de cepas do complexo C.
parapsilosis isoladas de fontes veterinárias e mantidas em nossa micoteca, com posterior
análise do perfil de sensibilidade a antifúngicos e de seus atributos de virulência in vitro.
5.2 Objetivos específicos
1- Confirmar a identificação fenotípica de isolados do complexo C. parapsilosis de origem
animal mantidos em banco de culturas fúngicas;
2- Realizar a identificação molecular dos isolados do complexo C. parapsilosis por meio da
análise de restrição do gene SADH;
3- Determinar o perfil de sensibilidade dos isolados in vitro;
30
6. CAPÍTULO 1
Complexo Candida parapsilosis e sua sensibilidade antifúngica e atributos de virulência
Candida parapsilosis complex from animals and its antifungal susceptibility and virulence
attributes
31
Candida parapsilosis complex from animals and its antifungal susceptibility and virulence attributes
Raimunda Sâmia Nogueira Brilhanteª, Terezinha de Jesus Santos Rodriguesª, André Jalles
Monteiroc, Débora de Souza Collares Maia Castelo-Brancoª, Carlos Eduardo Cordeiro
Teixeiraª, Ramila de Brito Macedoª, Silviane Praciano Bandeiraª, Rossana de Aguiar
Cordeiroª, Tereza de Jesus Pinheiro Gomes Bandeiraa,d, José Júlio Costa Sidrimª, Marcos
Fábio Gadelha Rochaa,b
a Department of Pathology and Legal Medicine, College of Medicine, Postgraduate Program
in Medical Microbiology, Specialized Medical Mycology Center, Federal University of
Ceará, Fortaleza, Ceará, Brazil.
b College of Veterinary Medicine, Postgraduate Program in Veterinary Sciences, State
University of Ceará, Fortaleza, Ceará, Brazil.
c Department of Statistics and Applied Mathematics, Federal University of Ceará, Fortaleza,
Ceará, Brazil.
d School of Medicine, Christus College - UNICHRISTUS, Fortaleza, Ceará, Brazil
CorrespondingAuthor: R.S.N. Brilhante. Rua Barão de Canindé, 210; Montese. CEP:
32
Abstract
Yeasts of the Candida parapsilosis complex have been identified as agents of
infections and as components of the microbiota of animals. The objectives of this study were
to perform the molecular identification of strains of the C. parapsilosis complex isolated from
veterinary sources and to assess their in vitro antifungal susceptibility and virulence attributes.
We used 28 isolates of C. parapsilosis (lato sensu) recovered from dogs, psittacines, raptors
and prawns and maintained in our fungal collection. Initially, the strains were phenotypically
characterized, based on the analysis of their morphological and biochemical features, which
confirmed their identity as C. parapsilosis (lato sensu). The molecular identification of the
species was performed by PCR-REA. Then, the susceptibility of the strains to amphotericin
B, itraconazole, fluconazole, voriconazole and caspofungin was assessed through broth
microdilution, according to CLSI (M27-A3). Additionally, the ability of the strains to produce
phospholipases, proteases and biofilm was analyzed. Genotypical analysis showed thirteen C.
parapsilosis (stricto sensu), ten C. orthopsilosis and five C. metapsilosis strains. In vitro
resistance to fluconazole was observed in three strains of C. parapsilosis (stricto sensu) and
two of C. metapsilosis, while high caspofungin MICs (2 μg/mL) were observed in one strain
of C. pa rapsilosis (stricto sensu) and five C. orthopsilosis. Additionally, 23/28 isolates
presented protease production, while none were able to produce phospholipases. All tested
strains were able to form biofilms, out of which twenty were moderate producers, seven were
strong producers and one was a weak producer. Our data indicate that the antifungal
susceptibility and the virulence traits vary among the species of the C. parapsilosis complex.
Keywords: Candida parapsilosis. Animals. Molecular identification. Antifungals. Virulence.
33
7. CONCLUSÕES
1. As cepas previamente identificadas como pertencentes ao complexo Candida parapsilosis,
isoladas de fontes veterinárias e mantidas na micoteca do Centro Especializado em Micologia
Médica tiveram sua identificação original confirmada, por análises morfológica e bioquímica
2. A análise de restrição do gene SADH se mostrou eficaz para distinguir as três espécies
pertencentes ao complexo C. parapsilosis e revelou que das 28 cepas analisadas, 13 eram C.
parapsilosis stricto sensu, 10 C. orthopsilosis e cinco C. metapsilosis. C. pa rapsilosis stricto
sensu foi a espécie predominante entre as cepas oriundas de cães, enquanto que C.
orthopsilosis foi predominante entre aquelas isoladas de psitacídeos.
3. A avaliação da sensibilidade antifúngica in vitro detectou resistência a fluconazol em cepas de C. parapsilosis stricto sensu e C. metapsilosis. Ademais, foram observadas concentrações
inibitórias mínimas elevadas para caspofungina, especialmente em C. orthopsilosis.
4. Nenhuma cepa do complexo C. parapsilosis secretou fosfolipases, mas 82,1%
34
8. PERSPECTIVAS
Com o avanço das técnicas moleculares e a consequente detecção de diferenças
genéticas entre microrganismos fenotipicamente indistinguíveis, novas espécies fúngicas tem
sido propostas. No entanto, ainda não se sabe ao certo a relevância clínico-epidemiológica
dessas distinções de espécies. Nesse contexto, a presente pesquisa visou buscar diferenças
fenotípicas entre as espécies do complexo Ca ndida parapsilosis, com destaque para a
sensibilidade antifúngica e para a produção de atributos de virulência in vitro. Dentre as cepas
avaliadas, observaram-se algumas diferenças entre as espécies do complexo quanto à
colonização de diferentes classes animais, ao perfil de sensibilidade antifúngica e à produção
de proteases. Portanto, torna-se necessário realizar mais estudos nessa área, visando à inclusão de um maior número de cepas de origem veterinária e à realização de estudos in vivo para
averiguar se essas diferenças in vitro estão associadas às diferenças na patogenicidade e na
35
REFERÊNCIAS
ALENCAR, L. P. Leveduras e Geotrichumcapitatum do Trato Digestivo e Fezes de Emas (Rhea americana) mantidas em cativeiro: Identificação e Sensibilidade Antifúngica in vitro. 2012. Dissertação (Mestrado em Ciências Veterinárias) - Faculdade de Veterinária, Universidade Estadual do Ceará, Fortaleza, 2012.
ÁLVARES, C. A. et al. Candidíase vulvovaginal: fatores predisponentes do hospedeiro e virulência das leveduras. Journal Brazilian Pathology Medical Laboratory, v. 43, n. 5, p.319-327, 2007.
ASADZADEH, M. et al. Rapid molecular differentiation strains isolated from clinical specimens in Kuwait. Journal Medical Microbiology, v. 58, p. 745–752, 2009.
ASHFORD, B. K. Moniliaparapsilosis. American Journal of Tropical Medicine, v.8, p. 507-518, 1928
BENTUBO, H. D. L. et al. Expectativa de vida e causas de morte em cães na área metropolitana de São Paulo (Brasil). Ciência Rural, v.37, p.1021-1026, 2007.
BRILHANTE, R. S. N. et al. Antifungal susceptibility and genotypical pattern of Microsporumcanisstrains. Canadian Journal of Microbiology, v. 51, p. 1480-3275, 2005.
BRILHANTE, R.S.N. et al. Yeasts from Macrobrachiumamazonicum: a focus on antifungal susceptibility and virulence factors of Candida spp. FEMS Microbiology Ecology, v.76, p.268-277, 2011.
BRILHANTE, R.S.N. et al. Characterization of the gastrointestinal yeast microbiota of cockatiels (Nymphicushollandicus): a potential hazard to human health. Journal Medical Microbiology, v.59, p.718–723, 2010.
BRILHANTE, R.S.N. et al. Detection of Candida species resistant to azoles in the microbiota of Rheas (Rhea americana): possible implications for human and animal health. Journal of
Medical Society General Microbiology, v. 62, p.000-000, 2013.
BRITO E. H. S. Caracterização fenotípica e perfil de sensibilidade antifúngica de cepas de candidaspp e malasseziapachydermatis, oriundas de cães. 2005. 85 f. Dissertação (Programa de Pós graduação em Ciências Veterinárias) da Faculdade de Veterinária da Universidade Estadual do Ceará, 2005.
BRITO, E. H. S. Uma abordagem fenotípica e molecular em leveduras do gêneroCandidaisoladas de cães. 2008. 135f. Tese (Programa de pós-graduação em Ciências Veterinárias da Faculdade de Veterinária) - Universidade Estadual do Ceará, Fortaleza, 2008.
BRITO, E.H.S. Uma abordagem fenotípica e molecular em leveduras do
36
BRITO, E.H.S. et al. The anatomical distribution and antimicrobial susceptibility of yeast species isolated from healthy dogs. The Veterinary Journal,v.182, p. 320-326, 2009.
CAFARCHIA, C. et al. Occurrence of yeasts in cloacae of migratory birds. Mycopathologia, v.161, p. 229-234, 2006.
CAFARCHIA, C. et al. Phospholipase, activity of yeasts from wild birds and possible implications for human diseases. Medical Mycology, v.46, p.1-6, 2008.
CLINICAL AND LABORATORY STANDARDS INSTITUTE. Reference method for broth dilution antifungal susceptibily testing of yeasts. 3rd ed. [S.l.], 2008. (CLSI document M27-A3, v. 28, n. 14)
COLOMBO, A.L.; GUIMARÃES, T. Epidemiology of hematogenous infections due to Candida spp. Rev. Sociedade Brasileira Medicina Tropical, v.36, n.5, p. 599-607, 2003.
CORDEIRO, R. A. et al. Species of Candida as a component of nasal microbiota of healthy horses. Medical Mycology, 2013.
COSTA, K. R. C. et al. Candida albicans and Candida tropicalis in oral candidosis: quantitativeanalysis, exoenzyme activity and antifungal drug sensitivity. Mycopathologia, v. 167, p. 73-79, 2009.
COSTA, K. R. C. Isolamento, quantificação, atividade enzimática e sensibilidade a antifúngicos de leveduras da saliva de pacientes imunocompetentes portadores de lesão bucal. 2006. 113f. Dissertação (Ciências farmacêuticas) – Universidade de São Paulo, Ribeirão Preto, 2006.
COUTO, E.M.P.; CARLOS, D.; MACHADO, E. R. Candidíase em Neonatos: uma revisão epidemiológica. Ensaios e Ciência Ciências Biológicas, Agrárias e da Saúde, v. 15, p. 197-213, 2011.
CRAGG, J.; CLAYTON, Y.M. Bacterial and fungal flora of seagull droppings in Jersey.
Journal of Clinical Pathology, v.24, p.317-319, 1971.
DE HOOG, G. S. et al. Atlas of Clinical Fungi. 2nd ed. Baarn: Centraal bureau voor Schimmsl cultures, 2000.
FALAGAS, M. E.; ROUSSOS, N.; VARDAKAS, K.Z. Relative frequency of albicans and the various non-albicans Candida spp among candidemia isolates from inpatients in various parts of the world: a systematic review. International Journal of Infectious Diseases, v.14, p.954-966, 2010.
GHANNOUM, M. A. Potential role of phospholipases in virulence and fungal pathogenesis. Clinical Microbiology Reviews, v. 13, n. 1, p.122-143, 2000.
37
KANO, R. et al. Molecular identification of Ca ndida parapsilosisfrom crop mucosa in a cockatiel. Journal of Veterinary Diagnostic Investigation, v. 13, p. 437-439, 2001.
KIVARIA, F.M.; NOORDHUIZEN, J.P.T.M.A retrospective study of the aetiology and temporal distribution of bovine clinical mastitis in smallholder dairy herds in the Dar es Salaam region of Tanzania. Veterinary Journal, v.173, p.617-622, 2007.
KONEMAN, E. et al. Diagnóstico microbiológico. 6. ed. Rio de Janeiro: Guanabara Koogan, 2008.
LEHMANN, P.F.; LIN, D.M.; LASKER, B.A. Genotypic identification and characterization of species and strains within the genus Candida by using random amplified polymorphic DNA. Journal Clinical Microbiology, v. 30, p. 3249-3254, 1992.
LIN, D.M. et al. Three distinct genotypes within Candida parapsilosis from clinical sources. Journal Clinical Microbiology, v. 33, p.1815-1821, 1995.
LOCKHART, S. R. et al. Geographic distribution and antifungal susceptibility of the newly described species Candida orthopsilosis and Candida metapsilosis in comparison to the closely related species Candida parapsilosis. Journal Clinical Microbiology, v. 46, p.2659-2664, 2008.
LOPES, A. et al. Successful caspofungin treatment of invasive refractory candidiasis in the extremely low birthweight neonate. Acta Médica Portuguesa, v. 23, p. 719-722, 2010.
MAIA, D.C.B.S.C. Leveduras isoladas do trato gastrintestinal determinação da microbiota e análise fenotípica de calopsitas (Nymphicushollandicus). 2009, 86 p. Dissertação (Mestrado em CiênciasVeterinárias) - Faculdade de Veterinária, Universidade Estadual do Ceará, Fortaleza, 2009.
MEDRANO, D.J.A. et al. Candidemia in a brazilian hospital:
theimportanceofCandidaparapsilosis. Revista do Instituto de Medicina Tropical de São Paulo, v.48, p.17-20, 2006.
MIRHENDI, H. et al. Molecular screening for Candida orthopsilosis and Candida metapsilosis among Danish Candida pa rapsilosis group blood culture isolates: proposal of a new RFLP profile for differentiation. Journal Medical Microbiology, v.59, p.414-420, 2010.
MONTAGNA, M.T. et al. Project Group Invasive fungal infections in neonatal intensive care units of southern Italy: a multicentre regional active surveillance (AURORA project).
Journalofpreventive medicine and hygiene, v.51, p.125-130, 2010.
MORETTI, A. et al. E. Isolation of Candida rugosafrom turkeys. Journal of Veterinary Medicine, v. 47, p. 433-439, 2000.
38
NAGLIK, J. et al. Candida albicans proteinases and host/pathogen interactions. Cellular Microbiology,v. 6, p. 915-926, 2004.
NOSEK, J. et al. Biology and genetics of the pathogenic yeast Candida parapsilosis. Current
Genetics, v. 55, p.490-509, 2009.
PANTOJA, L.D.M. et al. Ants (Hymenoptera: Formicidae) as carriers of fungi in Hospital Environments: An Emphasis on the Genera Tapinoma and Pheidole. Journal of Medical Entomology, 2009.
PRICE, M.F.; WILKINSON, I.D.; GENTRY, L.O. Plate method for detection of phospholipase activity of Candida albicans. Sabouraudia, v.22, p.201-207, 1982.
RUIZ, L. et al. Candidemia by species of the Candida parapsilosis complex in children’s hospital: prevalence, biofilm production and antifungal susceptibility. Mycopathologia, v.175, p. 231-239, 2013.
RUIZ, L.S. et al. Outbreak of fungaemia due to Candida parapsilosis in a neonatal intensive care unit: molecular genotyping by analysis microsatellite markers. Revista Iberoamericana Micologia,v.30, p.112-115, 2012.
SANTOS, R.C.; MARIN, J.M. Isolation of Ca ndida spp. from mastitic bovine milk in Brazil. Mycopathologia, v.159, p.251-253, 2005.
SIDRIM, J. J. C.; ROCHA, M. F. G. Antifungal susceptibility and genotypical pattern ofMicrosporumcanisstrains. Canadian Journal of Microbiology, v. 51, p. 1480-3275, 2005.
SIDRIM, J.J.C. et al. Candida parapsilosis meningitis as the first manifestation of AIDS: case report. Journal of Medical Microbiology, v. 60, p.1530-1533, 2011.
SIDRIM, J.J.C. et al. Candida species isolated from the gastrointestinal tractof cockatiels (Nymphicushollandicus): In vitro antifungal susceptibility profile and phospholipase activity. Veterinary Microbiology, v.145, p.324-328, 2010.
SILVA, S. et al. Candida glabrata, Candida parapsilosis and Candida tropicalis: biology, epidemiology, pathogenicityandantifungalresistance. FEMS Microbiology Reviews, v.36, p.288-305, 2012.
SPREGHINI, E. et al. In vitro and in vivo effects of echinocandins against. Candida, v. 67, p. 2195-2202, 2012.
TAMURA, N. et al. Fatores de virulência de Candidaspp isoladas de cateteres venosos e mãos de servidores hospitalares. Revista da Sociedade Brasileira de Medicina Tropical, v.40, p.91-93, 2007.
39
TAVANTI, A. et al. Genotyping of Candida orthopsilosis clinical isolates by amplification fragment length polymorphism reveals genetic diversity among independent isolates and strain maintenance within patients. Journal Clinica Microbiology, v.45, p.1455-1462, 2007.
TORO, M. et al. Characterization of Candida parapsilosis complex isolates. Clinical Microbiology Infection, v. 17, p. 418–424, 2011.
TOSUN, I. et al. Distibuition, virulence attibutes and antifungal susceptibility patterns of Candida parapsilosis complex stains isolated from clinical samples. Medical Mycology, v. 51, p. 483-492, 2013.
TROFA, D.; GA´CSER, A.; NOSANCHUKI, J. D. Candida parapsilosis, an emerging fungal pathogen. Clinical Microbiology Reviews, v.21, p. 606–625, 2008.
TSAI, S.S. et al. Aspergillosis and candidiasis in psittacines and passeriformes birds with particular reference to nasal lesions. Avian Pathology, v. 21, p. 699-709, 1992.
VIEIRA, R.G.; ACQUA-COUTINHO, S.D. Phenotypical characterization of Candida spp. isolated from crop of parrots (Amazonaspp.). Pesquisa Veterinária Brasileira, v. 29, p.452-456, 2009.