Genetic and biochemical characterization of hemicellulose polysaccharides utilization in Bacillus subtilis
183
0
0
Texto
Imagem


+7
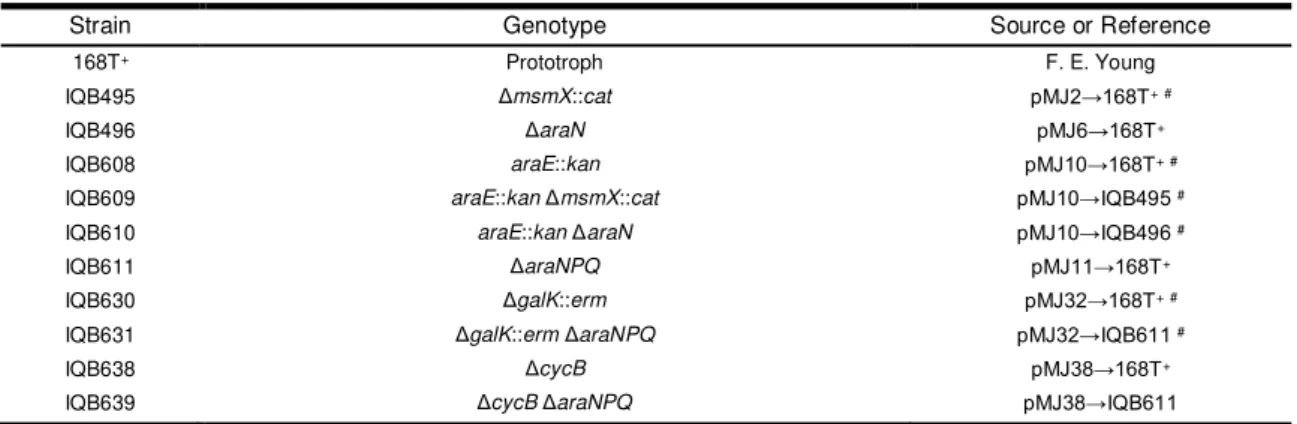
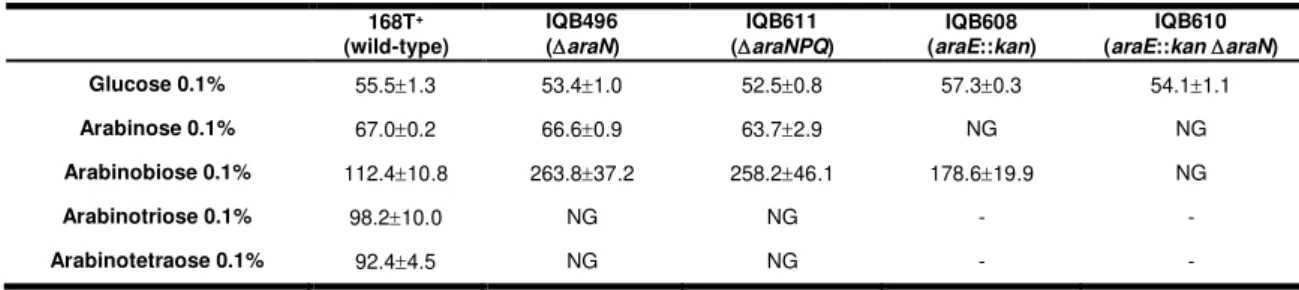


Documentos relacionados