The different response of sugarcane genotypes in multiple stress
Texto
Imagem

Documentos relacionados
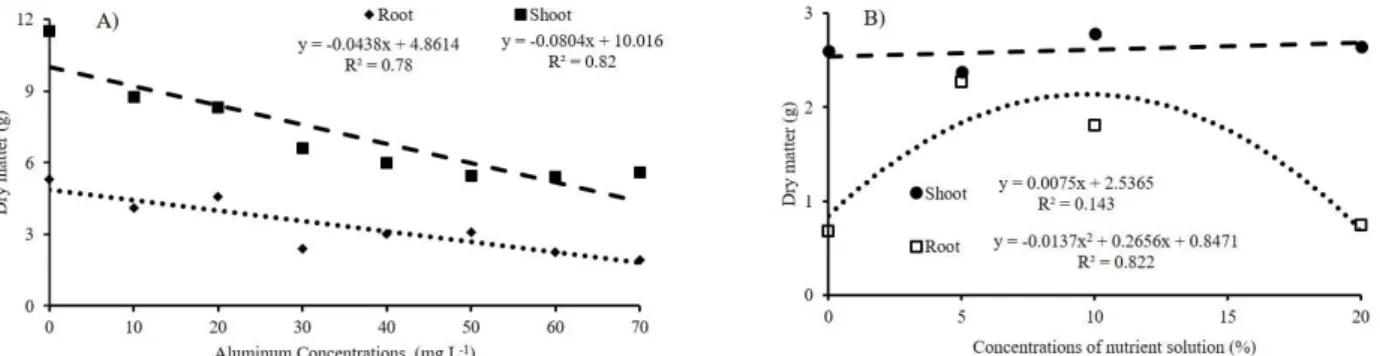
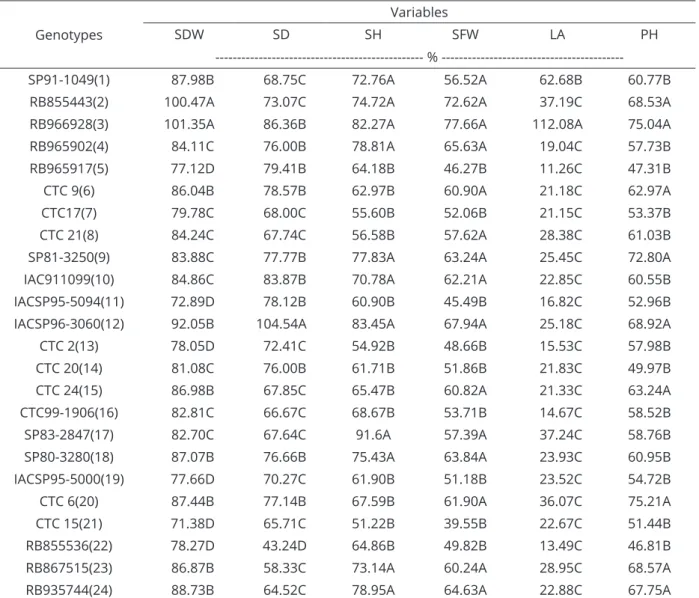
Regression adjustments for the variables: shoot length (with and without growth regulator); root length (cm); shoot and root fresh mass (g); shoot and root dry mass (g); leaf area
desc_accao toca_sequencia_3 toca_sequencia_4 toca_sequencia_3 escolhe_resposta_3 nivel_seguinte toca_sequencia_4 toca_sequencia_3 toca_sequencia_2 toca_sequencia_1
© 2003 John Wiley & Sons, Ltd ISBN 0 470 84909 6 (cased) ISBN 0 470 84910 X (pbk) 4 Cosmological Models In Section 4.1 we turn to the ‘concordance’
Similarmente ao que foi feito nos testes realizados anteriormente com a bobina de Rogowski apenas, as formas de onda observadas e registradas no osciloscópio utilizado são referentes
A partir dos estudos ressaltados neste tópico, é possível ver a importância de uma vida saudável, não só fisicamente como culturalmente, na qual sejam
[...] c. Permitir que os indivíduos e a coletividade compreendam a natureza complexa do meio ambiente natural e do meio ambiente criado pelos homens como resultante da
Baseada no modelo de estágios de desenvolvimento de leitura (estágio logográfico, alfabético e ortográfico), Frith (1985) define a dislexia do desenvolvimento como
Floral biology of Pilosocereus tuberculatus (Werderm.) Byles & Rowley: a bat pollinated cactus endemic from the “Caatinga” in northeastern Brazil. British bat calls:
