UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
Efeito da estrutura do habitat e da intensidade de pastoreio na
condição física de Apodemus sylvaticus em ambiente
agro-silvo-pastoril
Ricardo Jorge Ribeiro Fragoso
Mestrado em Biologia da Conservação
Dissertação orientada por:
Professora Doutora Margarida Santos-Reis
Doutor Luís Miguel Rosalino
O presente estudo foi realizado no âmbito de(a):
a) Um protocolo de investigação estabelecido desde 2007 entre a Companhia das Lezírias, S.A. (CL) e o Centro de Ecologia, Evolução e Alterações Ambientais (cE3c), unidade de I&D da Faculdade de Ciências da Universidade de Lisboa (FCUL).
b) Plataforma LTsER Montado (http://www.ltsermontado.pt/), dedicada à investigação sócio ecológica de longo prazo do sistema Montado.
i
Agradecimentos
Após um ano de trabalho excepcional, do qual nunca me vou esquecer, a tese finalmente acabou! Mas não posso deixar de agradecer a todas as pessoas que me ajudaram, pois sem elas nada teria sido feito.
À professora Margarida Santos-Reis, por me ter como aluno e dar a possibilidade de trabalhar com mamíferos, de me ter ensinado imensas coisas, por estar sempre disponível para responder às minhas dúvidas, quer quando estava em trabalho de campo quer quando já estava a escrever a tese, e por se esforçar por conseguir fazer as correcções mesmo que o tempo fosse escasso. Ao professor Luís Miguel Rosalino, por me ajudar na primeira semana de campo, dar ideias para variáveis a testar e mais importante, por me ter ajudado imenso em toda a estatística e escrita, estando sempre disponível para qualquer dúvida ou problema.
À Companhia das Lezírias, por me ter permitido o acesso aos seus terrenos e meios (enquanto os houve!) e estadia durante o trabalho de campo, e aos seus funcionários que sempre se demonstraram prestáveis para ajudar. Um agradecimento especial à Sandra Alcobia por me mostrado os cantos à Companhia, por me ter ajudado nos primeiros passos do trabalho de campo e me ter sempre ajudado cada vez que lhe ligava desesperado com algum problema.
Aos meus amigos que me mantiveram são, nas poucas vezes que saí do ritmo de trabalho de campo e “voltei à civilização”. Em especial à Sara e ao João por me ajudarem nalgumas etapas da tese ao mesmo tempo que me davam cama para dormir, mesmo sendo tão chato como sou. Um obrigado especial aos meus pais, que sempre me apoiaram, sempre estiveram no meu canto e me forneceram tudo o possível para conseguir fazer a tese e ser feliz.
ii
Resumo
Pressões antropogénicas, como a agricultura e o pastoreio, são bastante comuns em ecossistemas Mediterrânicos, como o montado. Estas pressões alteram significativamente os habitats, afectando as comunidades animais presentes, em especial os animais com uma mobilidade limitada, como é o caso dos pequenos mamíferos.
Este estudo pretendeu avaliar de que modo a presença de pastoreio e os diferentes níveis do habitat afectavam a condição corporal de Apodemus sylvaticus. Para tal, procedeu-se à captura de indivíduos para recolha das métricas corporais em locais pastoreado em simultâneo com locais excluídos ao pastoreio em diferentes anos (desde 1998, 2004 e 2008). Também se procedeu à caracterização do habitat onde essas capturas ocorreram.
O aumento da condição corporal não se revelou directamente proporcional com o período de exclusão, mas antes foi encontrado um certo nível de semelhança entre os locais de controlo com os locais isolados ao pastoreio desde 2004 e entre os locais isolados desde 1998 com os locais isolados desde 2008. Os machos possuíram uma maior média de condição corporal do que as fêmeas.
Em relação aos diferentes níveis do habitat, os modelos lineares generalizados revelaram que tanto variáveis de microhabitat como de macrohabitat influenciavam a condição corporal dos indivíduos, e que as variáveis importantes para os machos eram diferentes das importantes para as fêmeas. Assim, enquanto que para os machos, apenas a percentagem de cobertura de arbustos se tenha revelado como uma influência positiva sobre a condição corporal, para as fêmeas as variáveis correspondentes à percentagem de cobertura das gramíneas e a altura média destas possuíram influência positiva, ao passo que o número de sobreiros possuiu influência negativa sobre a condição corporal.
Estes resultados sugerem que a presença de abrigo e alimento são determinantes para a condição corporal de Apodemus sylvaticus, embora com diferente relevância para os machos e fêmeas, uma vez que estes possuem estratégias ecológicas diferentes. Apesar do pastoreio não se ter revelado como uma influência directa sobre a condição corporal, os habitats sobre esta pressão possuem abrigo e alimento para os pequenos mamíferos em menor disponibilidade, actuando assim como uma influência indirecta sobre estes animais. Deste modo, os locais isolados ao pastoreio intercalados com locais com pastoreio podem tornar-se locais de refúgio, com habitats mais complexos e com maior disponibilidade de recursos.
Por fim, é aconselhada uma gestão da paisagem que mantenha o mosaico de habitats e que permita conciliar as comunidades faunísticas nativas com os diferentes usos do solo.
iii
Summary
Anthropogenic pressures, such as agriculture and cattle grazing, are common in Mediterranean ecosystems, like the montado ecosystem. This pressures significantly alter the habitat, affecting the wild communities present, especially those with low vagility, such as the small mammals. This study aimed to assess how cattle grazing and the different habitat levels affected the physical condition of Apodemus sylvaticus. To collect physical metrics individuals were captured in grazed sites as well as sites excluded from grazing since different years (1998, 2004 and 2008). The characterization of the habitats were the captures took place was also made.
The physical condition increase wasn’t directly proportional with the exclusion period, but rather a certain degree of similarity was found between the grazed sites and sites excluded from grazing since 2004 and between the sites excluded since 1998 and the sites excluded since 2008. The males had a higher physical condition average than the females.
Regarding the different habitat levels, the generalized linear models showed that variable from both microhabitat and macrohabitat influenced the individual’s physical conditions, and that the variables important for the males were different from those important for the females. As so, while for the males only the shrub cover had a positive influence on the physical condition, for the females, grass cover and their average height had a positive influence, whereas the number of cork oak trees had a negative influence on the physical condition.
These results suggest that both the presence of food and shelter are determinants to the Apodemus
sylvaticus physical condition, although with different importance for males and females, since
they have different ecological strategies. While cattle grazing wasn´t a direct influence on the physical condition, habitats under this pressure have less availability of shelter and food for the small mammals. Thus, non-grazed sites interspersed with grazed sites can act like safe havens, with more complex habitats and more resources available
In the end, a landscape management is advised, to maintain the habitat heterogeneity and to allow a balance between the native animal communities and the different human land uses.
iv
Índice
Agradecimentos ... i Resumo ... ii Summary ... iii Lista de figuras ... v Lista de Tabelas ... v 1. Introdução ... 11.1. Os ecossistemas mediterrânicos e o papel do homem como modelador da paisagem 1 1.1.1 Caracterização dos ecossistemas mediterrânicos actuais. ... 1
1.1.2 Alterações antropogénicas da Paisagem Mediterrânica ... 1
1.1.3 A agricultura como agente de estruturação da paisagem ... 2
1.1.4 Pastoreio como agente de estruturação da paisagem Mediterrânica ... 2
1.1.5 O montado como sistema multifuncional ... 3
1.2. Biodiversidade: componente valorizadora das Paisagens Mediterrânicas ... 4
1.3. Pequenos Mamíferos: a base das comunidades de vertebrados ... 4
1.4. Condição corporal como indicador dos impactos dos ecossistemas agro-silvo-pastoris ... 6 1.5. Objetivos ... 6 2. Materiais e métodos ... 7 2.1. Área de estudo ... 7 2.2. Desenho amostral ... 8 2.3. Análise de dados ... 9 3. Resultados ... 12 3.1. Capturas ... 12
3.2. Índice de condição corporal da população de Apodemus sylvaticus ... 12
3.3. Hipóteses explicativas da variação da condição corporal ... 14
4. Discussão ... 21
4.1. Condição corporal vs exclusão pastoreio ... 21
4.2. Hipóteses explicativas da variação individual da condição corporal ... 21
4.3. Variáveis importantes para a condição corporal ... 21
4.4. A particularidade dos locais de com exclusão de pastoreio desde 2004 ... 23
4.5. Limitação dos dados ... 24
4.6. Considerações finais ... 24
5. Referências ... 26
v
Lista de figuras
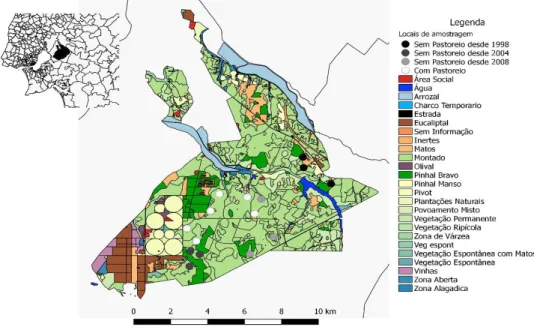
Figura 1 – Localização da área de estudo com os respectivos usos do solo e locais de amostragem. Figura 2 – Boxplot representando a mediana da condiçãos corporal de Apodemus sylvaticus por tipo de local de amostragem.
Figura 3 – Boxplot representando a mediana da condição corporal por sexo.
Figura 4 – Boxplot representando a variação da percentagem de cobertura por grupo de locais. Figura 5 – Boxplot representando a variação da percentagem de cobertura de gramíneas por grupo de locais.
Figura 6 – Boxplot representando a variação da altura média de gramíneas por grupo de locais. Figura 7 – Boxplot representando a variação do número de sobreiros por grupo de locais.
Lista de Tabelas
Tabela I – Descrição das variáveis e respectivas abreviaturas usadas nos modelos lineares generalizados.
Tabela II – Capturas das diferentes espécies de pequenos mamíferos por local de amostragem. Tabela III – Modelos lineares generalizados para as três primeiras hipóteses explicativas da variação da condição corporal dos machos.
Tabela IV – Modelo médio para a hipótese microhabitat com intervalos de confiança de 95% para cada variável.
Tabela V – Modelos lineares generalizados para a hipótese mista para os machos.
Tabela VI – Modelo médio para a hipótese mista para os machos, com intervalos de confiança de 95% para cada variável.
Tabela VII – Modelos lineares generalizados para as três primeiras hipóteses explicativas da variação da condição corporal das fêmeas.
Tabela VIII – Modelo médio para a hipótese microhabitat com intervalos de confiança de 95% para cada variável.
Tabela IX – Modelos lineares generalizados para a hipótese mista para as fêmeas.
Tabela X – Modelo médio para a hipótese mista para as fêmeas, com intervalos de confiança de 95% para cada variável.
1
1. Introdução
1.1. Os Ecossistemas Mediterrânicos e o papel do homem como modelador da paisagem 1.1.1 Caracterização dos Ecossistemas Mediterrânicos actuais.
A bacia do Mediterrâneo cobre uma área de 2,5 milhões de quilómetros quadrados, estendendo-se por 34 paíestendendo-ses e albergando cerca de 400 milhões de pessoas (Valavanidis 2013). Embora muita da área anteriormente fosse coberta por florestas de folha caduca e coníferas, a intervenção humana, que remonta desde há 8000 anos, mudou claramente toda a paisagem (Valavanidis 2013), tanto que actualmente apenas 4,7 % da vegetação original permanece (Geri 2010). O clima típico da região é caracterizado por Verões quentes e secos e Invernos relativamente frios e húmidos, resultando num tipo paisagem que se assemelha a uma savana, com reduzida densidade arbórea e maioritariamente composta por espécies arbustivas, paisagem esta bastante característica da bacia (Blondel 2006). Uma reduzida precipitação associada a práticas agrícolas insustentáveis causou uma degradação grave dos solos, levando mesmo a que muitas áreas corram o risco de desertificação, como o caso de Portugal, onde 60% do território se encontra em risco moderado (Valavanidis 2013). De facto, estudos focados em Ecossistemas Mediterrânicos mostram que estes são dos mais ameaçados do planeta (Valavanidis 2013). Mesmo assim, não se pode deixar de ter em conta que esta bacia possui uma grande dinâmica ao nível da estrutura dos habitats com alterações constantes, em grande parte devido a intervenção humana e, associada a esta dinâmica, uma grande diversidade quer de fauna como de flora (Blondel 2006).
1.1.2 Alterações antropogénicas da Paisagem Mediterrânica
A bacia do Mediterrâneo albergou as primeiras civilizações com produções agrícolas (Zeder 2008), civilizações estas que moldaram a paisagem durante milénios, levando os ecossistemas a estar frequentemente sob pressão devido ao aumento da densidade populacional humana, à gestão agrícola inadequada, à urbanização e às actividades industriais (Valavanidis 2013), causando grandes alterações nos ecossistemas e no biota de toda a bacia (Blondel 2006), alterações estas que causaram, entre outras, erosão dos solos, desflorestação e perda de biodiversidade (Blondel 2006; Bajocco 2012). De entre estes, o maior efeito desta antiga presença humana é a conversão dos habitats naturais em terrenos agrícolas ou urbanos, estimando-se que esta conversão atinja os 40 % (Valavanidis 2013).
A destruição das florestas nativas teve duas grandes consequências: a conversão das florestas decíduas em florestas esclerofilas e matos, o que afetou as espécies tanto ao nível genético como na sua distribuição, e a dessecação da bacia como um todo (Blondel 2006), devido principalmente a mudanças no uso do solo e da água para a agricultura (Blondel 2006).
O facto de serem pressões muito antigas permitiram às espécies residentes evoluírem e adaptarem-se aos impactos negativos sofridos, levando a que actualmente muitas espécies adaptarem-sejam resilientes a pressões como o pastoreio (Alrababah 2007). Em algumas situações esta pode ter sido benéfica pois criou ecossistemas dependentes da gestão humana tradicional (Gonçalves 2011). Estes ecossistemas são caracterizados por espécies de flora que prosperam com usos humanos e com a presença de gado em quantidades limitadas e que provavelmente não existiriam caso estes fossem abandonados ou a gestão se alterasse ao nível da intensidade do uso dos solos (Tasser & Tappeiner 2002).
2
1.1.3 A agricultura como agente de estruturação da paisagem
A agricultura é o uso de solo mais importante na Europa, com 34% de todo o território usado nesse sentido e 14% usado para pastagens (Reidsma 2006). No Mediterrâneo a agricultura sempre teve forte presença, sendo este um dos mais importantes factores responsáveis pela criação dos ecossistemas existentes na bacia actualmente e pelo mosaico de diferentes habitats presente, como já foi abordado anteriormente. Porém nos últimos três séculos a área destinada a esta prática aumentou consideravelmente, 466% para as áreas cultivadas e 2400% para culturas de regadio (Meyer & Turner II 1992), expansão esta que levou à destruição de grande parte dos elementos da paisagem, como florestas, matas e prados naturais (Jongman 2002).
Apesar da história agrícola da região, os terrenos existentes na bacia Mediterrânica não são propriamente propícios à agricultura (Perez 1990). Os solos não possuem a mesma abundância de nutrientes que locais mais férteis como é o caso de habitats tropicais (Perez 1990), o sistema hidrológico não é o ideal, já que a precipitação não é abundante (Perez 1990), e as colheitas são limitadas (Perez 1990). Apesar das más condições, as produções continuam e, associados ao crescimento das áreas agrícolas, surgiram sistemas mais intensivos, com uso de maquinaria pesada e componentes químicos prejudiciais para os ecossistemas (Pinto-Correia & Mascarenhas 1999).
A gestão cuidada dos sistemas agrícolas torna-se assim essencial para a conservação, já que sistemas intensivos e a grande escala levam a graves consequências ambientais nos ecossistemas (Reidsma 2006), pois estes não têm capacidade de sustentar este tipo de práticas (Pinto-Correia & Mascarenhas 1999). Mesmo assim, enquanto que noutras partes do planeta a conversão dos terrenos para a agricultura trouxe consequências negativas para os ecossistemas onde é praticada (Dunn 2004), nos climas Mediterrânicos, em especial na bacia do Mediterrâneo, esta ainda está associada a elevados valores de biodiversidade e a ecossistemas saudáveis (Bugalho 2011), onde a intensidade da actividade o permita, já que a biodiversidade depende maioritariamente da intensidade agrícola (Reidsma 2006).
1.1.4 Pastoreio como agente de estruturação da paisagem Mediterrânica
Uma das perturbações antrópicas típicas do Mediterrâneo é a pastorícia, actividade com grande história nesta região, já que o gado pasta nos bosques Mediterrânicos desde há milhares de anos (Schoenbaum 2017). Apesar desta tradição, o pastoreio não é ideal no Mediterrâneo, já que capacidade de carga de gado nos ecossistemas é reduzida quando comparada com o resto da Europa, conseguindo apenas sustentar metade dos indivíduos que os ecossistemas Atlânticos suportam (Perez 1990). Esta reduzida capacidade de carga dos terrenos da bacia foi tradicionalmente ultrapassada pelo regime de transumância, usado na maioria dos países Mediterrânicos, no qual o gado é deslocado para zonas de reduzida altitude no Inverno e Primavera e para zonas mais altas no Verão, quando estas são mais produtivas ao nível das pastagens (Perez 1990).
O pastoreio é considerado como o factor mais importante na degradação dos solos já que actua tanto directa como indirectamente, quer pelo sobrepastoreio como pela conversão de terrenos em pastagens (Bajocco 2012). Mesmo assim, nos sistemas tradicionais a biodiversidade aparenta não ser afectada, já que devido à antiga presença de gado e consequente pastoreio, as comunidades evoluíram no sentido de responder a esta pressão antropogénica (Osem et al. 2002), assim o pastoreio, não tende a afectar a biodiversidade dos ecossistemas Mediterrânicos, tanto quer ao nível da fauna como da flora (Milchunas et. al 1988). O pastoreio traz consequências para os solos, quer por pisoteio excessivo, compactando-os (Ascensão, 2012), quer pelo sobrepastoreio que remove o subcoberto arbustivo e convertendo-o em solo nú (Castro & Freitas 2009). Com
3 certos valores mais elevados de pastoreio, a regeneração do estracto arbóreo e dos arbustivos tende a diminuir ou mesmo desaparecer (Plieninger 2007).
Porém o pastoreio é possível ser sustentável, desde que haja uma gestão cuidada ao nível da sua intensidade (Kairis 2015). Um exemplo disso é o pastoreio tradicional, que aumenta a diversidade de habitats presente nos ecossistemas Mediterrânicos (Lumaret 1994), já que cria áreas abertas sem arbustos ou árvores (Verdu 2000). Práticas mais recentes promovem a homogeneidade através da distribuição uniforme do pastoreio por toda a área disponível e um maior número de cabeças de gado, o que possui um efeito extremamente negativo na biodiversidade e nos habitats (Fuhlendor & Eagle 2001). O outro extremo, que corresponde ao completo abandono do pastoreio e das pastagens, também prejudica o ecossistema (Pinto-Correia & Mascarenhas 1999) porque permite a expansão do subcoberto arbustivo e de outras árvores, reduzindo a produção de cortiça devido à maior competição entre as árvores, e aumentando o risco de incêndio (Pinto-Correia & Mascarenhas 1999).
Para a existência de ecossistema saudável é necessário haver um equilíbrio, uma heterogeneidade promovida por pastoreio em locais específicos que produz um mosaico de habitats, que é típico dos ecossistemas locais e aumenta a biodiversidade (Fuhlendor & Eagle 2001). Deste modo, o ideal seria um modelo composto por 4 tipos de gestão em mosaico: exclusão completa do pastoreio, rotação do gado, plantação de árvores esparsas em zonas erodidas e revegetação das zonas degradadas usando espécies nativas e tolerantes à pastorícia (Noymeir & Oron 2001; Alrababah 2007). A rotação do pastoreio permite a coexistência de espécies adaptadas a esta pressão e de espécies intolerantes (Moog et al. 2002). Assim, o sistema de pastoreio tradicional, que promove esta rotatividade do gado e das zonas das pastagens, é o sistema que possui uma maior diversidade de flora, aumentando a resiliência do ecossistema (Alrababah 2007).
1.1.5 O montado como sistema multifuncional
Em Portugal, o sistema existente mais característico da bacia do Mediterrâneo, com todas as suas características e influências, é o montado, sistema agro-silvo-pastoril que ocorre maioritariamente na Península Ibérica, sendo que em Portugal o montado de sobro ocupa 730 mil hectares (Pio 2008). Este habitat é caracterizado pela presença de espécies do género Quercus, principalmente
Quercus suber, coexistindo com uma grande variedade de subcobertos, desde arbustos
normalmente mantidos a pequenas densidades (Gonçalves 2011) a pastagens (Bugalho 2011), e em mosaico com vegetação ripícola e zonas agrícolas, conseguindo sustentar elevados valores de biodiversidade (Godinho 2010; Gonçalves 2011).
Neste habitat, a gestão humana é essencial para a sua persistência já que sendo um sistema agro-silvo-pastoril, possui um equilíbrio no qual um ecossistema silvestre é conciliado e coexiste com as pressões antropogénicas (Bugalho 2011), desde que estas pressões não sejam intensificadas nem abandonadas (Gonçalves 2011). O montado de sobro é um sistema multifuncional usado maioritariamente para a produção de cortiça e madeira e para a criação de gado (Godinho,2010; Bugalho 2011), actividades que criaram uma estrutura complexa de diferentes usos dos solos. Fora isto, os montados de sobro também são importantes para regularização do ciclo hidrológico e para o sequestro de carbono (Kerstin 2010). Estas características do montado reforçam a sua importância conservacionista, sendo considerado um habitat prioritário pela directiva Habitats e uma componente importante da Rede Natura 2000 (Olea 2006).
4
1.2. Biodiversidade: componente valorizadora das Paisagens Mediterrânicas
As alterações da paisagem que ocorreram na bacia do Mediterrâneo para uso humano promoveram uma elevada diversidade de habitats, criando um sistema em mosaico complexo e com bastante dinâmica (Geri 2010). Este mosaico de habitats que caracterizam os actuais ecossistemas Mediterrânicos é o responsável pela existência de um dos principais factores que conferem o elevado valor conservacionista da região Mediterrânica, a sua biodiversidade (Valavanidis 2013). De facto, mesmo com a elevada pressão antropogénica, os valores de biodiversidade são únicos. Apesar de apenas cobrirem cerca de 2% de toda a superfície terreste, estes ecossistemas possuem 20% de todas as espécies de plantas, apenas sendo ultrapassados pelas florestas tropicais (Valavanidis 2013). Este facto torna a bacia do Mediterrâneo um dos hotspots mundiais de biodiversidade (Geri 2010; Valavanidis 2013), não só simplesmente pela grande quantidade de espécies residentes, mas também pela quantidade de endemismos, que é bastante elevada (Myers 2000). O montado é um bom exemplo, uma vez que diversas espécies em risco de extinção e ameaçadas, como o lince Ibérico (Lynx pardinus), o gato bravo (Felis silvestris) e o rato de Cabrera (Microtus cabrerae), ou importantes do ponto de vista da conservação, como o coelho bravo (Oryctolagus cuniculus), sobrevivem aqui (Bugalho 2011).
1.3. Pequenos Mamíferos: a base das comunidades de vertebrados
Tal como foi dito anteriormente, o pastoreio. ao ser um agente transformador da paisagem, uma vez que reduz a regeneração do estrato arbóreo e a cobertura de arbustos e, com o correspondente aumento do pasto, afecta as populações animais que usam estes ambientes, em especial aquelas com reduzida capacidade dispersora, como é o caso dos pequenos mamíferos (Morris 1992; Tew & MacDonald 1994; Gonçalves 2011).
Os pequenos mamíferos Mediterrânicos (Mammalia: ordens Rodentia e Eulipotyphla) são muito importantes do ponto de vista ecológico para as cadeias tróficas por serem a principal presa de animais de níveis tróficos superiores, como por exemplo aves de rapina nocturnas e mamíferos carnívoros (Tew 2000; Rosalino 2011). Para além deste facto, são muito suscetíveis a alterações ambientais, tendo uma resposta bastante célere às perturbações ambientais (Cabral 2005), permitindo avaliar rapidamente os impactos destas alterações, e servindo para avaliar os efeitos
bottom-up nas comunidades (Wei-chun 1989). Estes animais foram considerados como
indicadores de sustentabilidade nos ecossistemas (Alain 2006), tornando os estudos focados nos pequenos mamíferos ainda mais importantes.
Os pequenos mamíferos tendem a ser mais afectados pela agricultura do que os mamíferos de maior porte devido a possuírem menores áreas vitais e menor capacidade de dispersão (Burgess & Sharpe 1981; Lomolino & Perault 2000).
Em locais de pastoreio, a diminuição do coberto arbustivo reduz a disponibilidade de refúgios, bem como a abundância de alimentos (Eccard et al. 2000), aumentado o risco de predação (Torre et al. 2007; Ascensão 2012). Outro impacto do pastoreio nos pequenos mamíferos é o efeito do pisoteio, que compacta os solos e dificulta a formação de tocas (Bilotta et al. 2007; Ascensão 2012). Em contraste, zonas com vegetação natural intacta tendem a possuir melhores condições de habitat (ex. refúgio) e maior abundância de alimento (Gonçalves 2011; Ascensão 2012; Gheler-Costa 2012).
De facto, certas práticas agrícolas, como o sobrepastoreio, são de tal maneira prejudiciais para as comunidades de roedores e soricomorfos, que estas são apontadas como uma das principais causas
5 de diminuição das suas populações. Mesmo assim, pastoreio a níveis intermédios de intensidade favorece os pequenos mamíferos (Schmidt 2005), que mantêm populações semelhantes a zonas sem pastoreio (Schmidt 2005). Isto deve-se ao facto de esta intensidade média permitir a manutenção de vegetação resistente ao pastoreio sem diminuir a sua cobertura drasticamente (Schmidt 2005).
Uma das espécies de roedores mais abundantes em terrenos agrícolas na Península Ibérica é o rato-do-campo (Apodemus sylvaticus, Linnaeus 1758), um roedor granívoro que também consome frutos e insectos (Khammes & Aulagnier 2007). Consequentemente, o seu carácter generalista permite-lhe adaptar-se a paisagens em constante mudança, como é o caso do montado, o que não sucede com outras espécies mais especialistas (Love et al. 2000).
Para ser possível manter populações estáveis de pequenos mamíferos em sistemas antrópicos e extremamente heterogéneos, são necessários mais estudos focados nos habitats e recursos destes sistemas, em especial nos diferentes níveis relacionados com estas espécies (Noguerales 2015). Por norma, para os pequenos mamíferos, o habitat é entendido a dois níveis; o macrohabitat e o microhabitat. O macrohabitat é definido como o tipo de vegetação dominante e o seu contexto paisagístico, ou seja, a sua complexidade e heterogeneidade (Noguerales 2015) enquanto o microhabitat é normalmente estudado ao nível da estrutura da vegetação (Kotler & Brown 1988). Alguns estudos confirmam que a distribuição e abundância dos pequenos mamíferos são determinadas pela estrutura do microhabitat (Bellows et al. 2001; Jorgensen 2004; Traba et al. 2009; Tarjuelo et al. 2011) enquanto outros referem que estas são mais facilmente explicadas pelas características do macrohabitat (Morris, 1984, 1987; Orrock et al., 2000; Corbalán 2006). No que se refere ao microhabitat, a disponibilidade de alimento e a percentagem de cobertura vegetal são os principais factores envolvidos na sua seleção pelos pequenos mamíferos (Traba et al. 2009; Tarjuelo et al. 2011). O rato-do-campo aparenta preferir microhabitats caracterizados por elevadas percentagens de cobertura arbustiva (Boitani 1985; Khidas et al. 2002; Torre et al. 2002; Michel 2007), tipicamente acima dos 30% de cobertura (Abramsky 1981), já que diminui o risco de predação e oferece zonas de alimento seguras debaixo da vegetação (Díaz et al. 1993; Morris & Davidson 2000), evitando locais sem subcoberto, devido ao elevado risco de predação (Rosalino, 2011). Alguns estudos até apontam uma dependência maior da estrutura da vegetação do que da disponibilidade de sementes, alimento bastante importante para a espécie (Tew 2000; Noguerales 2015).
Em locais em que o pastoreio é muito intenso, ou seja, na ausência de subcoberto denso, os pequenos aglomerados de arbustos na base das árvores normalmente representam o único abrigo existente, providenciando proteção contra predadores e alimento (Tew 2000; Gonçalves 2011). Tradicionalmente, o impacto de sistemas agrícolas e silvícolas nos pequenos mamíferos tem sido avaliado em termos de riqueza específica e diversidade (Burel 1998; Tew 2000; Alain 2006; Ascensão 2012;; Gheler-Costa 2012; Noguerales 2015,), mas poucos autores consideraram a condição corporal como indicador da saúde destas populações (Peig & Green 2009), o que pode ser importante pois estudos focados na condição corporal de pequenos mamíferos permitem comparar populações em sistemas agrícolas sobre as quais existem pressões negativas, como o pastoreio, com outras populações que não sofram estas pressões.
6
1.4. Condição corporal como indicador dos impactos dos ecossistemas agro-silvo-pastoris
Muitos estudos de ecologia animal baseiam-se em métodos não letais para estimar a condição corporal de diferentes indivíduos numa população, contando com medidas de massa corporal e medidas de tamanho corporal para calcular um índice de condição física (Green 2001; Stevenson & Wood, 2006). A condição corporal de um animal pode ser definida como o capital de energia acumulado no corpo como resultado da alimentação, que é assumida como um indicador da qualidade e saúde de um animal (Peig & Green 2009). Estes índices são importantes por permitirem comparações entre diferentes populações (Peig & Green 2010). Com recurso a estes índices também é possível analisar se as características e perturbações no habitat influenciam a fitness dos indivíduos (Maceda-Veiga et al. 2014), já que as condições corporais de uma população sob pressão de factores extrínsecos ou de habitats degradados serão menores do que as de uma população num habitat natural não perturbado.
Apesar de estudos anteriores sugerirem que a abundância de pequenos mamíferos é menor em locais com pastoreio, principalmente devido à diminuição da vegetação (Burel et al. 1998; Ascensão 2012; Noguerales 2015), poucos estudos analisaram a condição física (Carreiras 2014; Teixeira 2015), nos quais não obtiveram resultados que demonstrassem uma variação significativa da condição corporal pelos habitats analisados.
1.5. Objetivos
O objetivo deste estudo consiste em avaliar se, e de que forma, os diferentes níveis do habitat e a intensidade de pastoreio influenciam a condição física da comunidade de pequenos mamíferos que habita uma paisagem agro-silvo-pastoral, com ênfase no rato-do-campo Apodemus sylvaticus. Para tal são propostas 4 hipóteses passíveis de explicar variação nas condições corporais do referido modelo animal: 1) as características do microhabitat, 2) as características do macrohabitat, 3) as relações intra e interespecíficas dos indivíduos, e 4) hipótese mista entre as anteriores.
7
2. Materiais e métodos
2.1. Área de estudo
O estudo decorreu na Charneca do Infantado da Companhia das Lezírias S.A., empresa que possui a maior área de exploração agro-silvo-pastoril em Portugal. Localizada entre os rios Tejo e Sorraia compreende como principais áreas a Charneca do Infantado, a Lezíria de Vila Franca de Xira, o Catapereiro e os Pauis (Magos, Belmonte e Lavouras), abrangendo os concelhos de Benavente, Vila Franca de Xira e Salvaterra de Magos.
O clima é tipicamente Mediterrânico, com Verões quentes e seco e os Invernos frios e húmidos, onde a temperatura média anual é de 16.3º C e a pluviosidade média anual de 700 mm.
A Charneca do Infantado ocupa cerca de 10.000 ha (Companhia das Lezírias S.A. 2017a) e é caracterizada por solos arenosos, pobres e com problemas de drenagem. Esta área é maioritariamente florestada por montado de sobro (6.725 ha), embora também existam plantações de pinheiro bravo (971 ha), de pinheiro manso (508) e de eucalipto (476) (Companhia das Lezírias S.A. 2017b). Para além das zonas de floresta, existem também áreas agrícolas para a produção de milho em pivots, olivais e pauis reservados à produção de arroz. Tratando-se de paisagens bastante planas, possui várias linhas de água, embora apenas uma possua caudal permanente, a Ribeira de Vale Cobrão, existindo ainda vários charcos temporários (Figura 1). A Charneca também é caracterizada pela presença do gado bovino, com cerca de 3000 animais (Gonçalves 2011), na qual o pastoreio ocorre nas zonas de montado de sobro devidamente vedadas para esse propósito, de modo extensivo e biológico e em regime anual de transumância, uma vez que são deslocadas entre a Charneca no Inverno e a Lezíria no Verão (Companhia das Lezírias S.A. 2017c). Nos locais onde o pastoreio ocorre, a densidade de arbustos é bastante reduzida, quer pelo gado, para o cultivo de pasto ou para prevenção de incêndios florestais (Gonçalves 2011). Cerca de 700 hectares da área de estudo encontram-se vedados para exclusão do gado e para protecção dos habitats. Estas áreas são maioritariamente compostas por montados de sobro em várias etapas da sucessão ecológica (Gonçalves 2011).
8 Devido ao mosaico de diferentes espécies arbóreas presente na Charneca do Infantado, que confere um grande número de habitats disponíveis para a fauna, a biodiversidade encontrada é elevada, existindo um grande número de espécies importante do ponto de vista da conservação, como é o caso da águia de Bonelli (Aquila fasciata), do toirão (Mustela putorius), do gato bravo (Felis silvestris) e do rato de Cabrera (Microtus cabrerae), para além de muitas outras espécies mais comuns e algumas cinegéticas. Em termos da comunidade de pequenos mamíferos, esta encontra-se bem representada e distribuída pela área de estudo, inclusive nos locais de amostragem (Carreiras, 2014) com um total de cinco espécies da ordem Rodentia (Apodemus
sylvaticus, Mus spretus, Rattus norvegicus, Microtus lusitanicus e Microtus cabrerae), e quatro
da ordem Eulipotyphla (Erinaceus europaeus, Talpa ocidentalis, Suncus etruscus e Crocidura
russula) descritas (Gonçalves 2011; Gonçalves 2013). De facto, a biodiversidade presente na área
é de tal maneira relevante que a maior parte da Charneca está inserida na Zona de Proteção Especial do estuário do Tejo.
2.2. Desenho amostral
Para dar cumprimento ao objectivo do trabalho, procedeu-se à comparação entre locais com pastoreio e locais não pastoreados, tendo sido selecionados 15 locais de amostragem (Figura 1): 6 com a presença de gado, actuando como locais de controlo, e os restantes 9 locais excluídos ao pastoreio em diferentes anos: 3 desde 2008, 3 desde 2004 e 3 desde 1998. Esta diferença temporal no período de exclusão torna-se relevante pois tem como consequência diferentes graus de complexidade do habitat com potencial implicação na condição corporal dos pequenos mamíferos.
A amostragem decorreu entre os meses de Setembro de 2017 e Maio de 2018, abrangendo os 2 períodos de maior mobilidade para a espécie, o de pré-reprodução, em Outubro e Novembro, e o de pós-reprodução, em Março e Abril. De modo a que a condição corporal não sofresse a influência da época de amostragem foram amostrados em simultâneo locais com a presença de gado e locais excluídos ao pastoreio em diferentes anos.
Em cada local de amostragem foram colocadas 25 armadilhas Sherman (Folding Traps whith
Aluminium Treadle and Doors Galvanized), com uma dimensão de 8x9x23 centímetros, dispostas
em cruz e espaçadas cerca de 10 metros entre si. Dentro de cada armadilha foi colocado um isco composto por sardinha em lata e flocos de aveia e algodão cardado para servir de abrigo e manter as condições térmicas para os indivíduos dentro da armadilha, reduzindo assim o seu stress. O tempo de amostragem correspondeu a 4 noites seguidas de amostragem, onde as armadilhas eram verificadas todas as manhãs, repondo o isco ou algodão quando necessário e substituindo o isco após a segunda noite. Antes da amostragem as armadilhas foram colocadas 2 noites no campo, permanecendo fechadas, para permitir a habituação dos animais (Grunell & Flowerdew 2006). As armadilhas estiveram activas durante vários períodos de quatro noites, até serem capturados, no mínimo, 10 Apodemus sylvaticus
Os animais capturados foram sedados recorrendo ao uso de vapores de éter e tratados de forma asséptica. Para cada um dos indivíduos capturados foi registado o sexo e a classe etária, sendo de seguida retiradas as métricas necessárias para a identificação da espécie e para o cálculo da condição corporal: comprimento total (do qual para a condição corporal se retirou o comprimento da cauda), comprimento da cauda, comprimento da pata posterior e peso. Também era importante identificar se as fêmeas estavam prenhas ou em lactação, já que as que estavam não poderiam
9 entrar nos cálculos de condição corporal. Por fim, na primeira captura de cada indivíduo realizou-se a um corte do pêlo de forma a identificar eventuais recapturas e evitar duplicação de dados, sendo libertados no mesmo local da captura após a recuperação da anestesia
Em cada local de captura de um indivíduo procedeu-se à recolha dos parâmetros de habitat. O micro-habitat foi avaliado num raio de 3 metros em redor da armadilha, permitindo avaliar quais as condições onde o mesmo foi capturado (Tew 2000), tendo sido calculadas as percentagens de cobertura da vegetação (árvores, arbustos, gramíneas, árvores de fruto e silvados) e a altura média das gramíneas e arbustos. O macrohabitat foi analisado numa faixa de 50 metros em redor da linha de armadilhas nos quais se avaliou a topografia (planície, vale, colina e declive), a heterogeneidade do habitat (homogéneo, moderadamente heterogéneo e heterogéneo), a densidade do subcoberto (denso, disperso e matos agregados) e a densidade arbórea (número de árvores nos 50 metros).
Em cada local de amostragem procedeu-se ainda ao levantamento de indícios de carnívoros (ex. dejetos, pegadas e tocas) de forma a avaliar o risco de predação composto por 3 níveis consoante a frequência de ocorrência dos indícios e o número de espécies: nível 1 para indícios de 1 ou 2 espécies, nível 2 para indícios de 3 espécies, e nível 3 para indícios de mais de 3 espécies.
2.3. Análise de dados
Em primeiro lugar procedeu-se ao cálculo das condições corporais dos ratos de campo capturados utilizando o Scaled Mass Index (Peig & Green 2009). Para este cálculo foram excluídos os juvenis bem como as fêmeas prenhas ou em lactação.
Scaled Mass Index : 𝑀̂𝑖 = 𝑀𝑖[ 𝐿0 𝐿𝑖]
𝑏𝑆𝑀𝐴
𝑀𝑖 − Peso do corpo do indivíduo
𝐿𝑖− Comprimento da cabeça e tronco do indivíduo
𝑏𝑆𝑀𝐴− Estimado pelo “Standardized Major Axis” (SMA) regressão de M no L
𝐿0− Valor arbitrário do L (por exemplo, média aritmética da população em estudo)
𝑀̂𝑖− Condição corporal do indivíduo
De seguida, para analisar a variação das condições por ano de exclusão, e testando previamente os pressupostos de homogeneidade e homocedasticidade, foi realizada uma ANOVA, tendo como variável independente o número de anos de exclusão ao pastoreio, ou seja, os locais de 1998, 2004, 2008 e os locais de controlo. Para analisar possíveis diferenças entre os grupos de locais foi realizado o teste de Tukey.
As variáveis passíveis de influenciar as condições corporais (Tabela I) foram agrupadas em 3 grupos considerando as hipóteses em teste: microhabitat para as variáveis relativas à estrutura do habitat, macrohabitat para o tipo de habitat dominante e o número de anos de exclusão ao pastoreio e relações para o nível de risco de predação, o número de indivíduos de outras espécies e o número de outros Apodemus capturados naquela armadilha. Foi testada a multicolinearidade
10 para as variáveis correspondentes ao microhabitat usando o fator de inflação das variâncias (VIF)(Zar 2010). Assim, usando o nível limite sugerido por Zuur (Zuur et al. 2007), foram excluídas todas as variáveis com VIF>5, calculando o VIF novamente para as restantes variáveis até nenhuma possuir valores acima do nível de corte.
Para analisar as diferenças entre os dois sexos, foram desenvolvidos modelos lineares generalizados (GLM) com distribuição Gaussiana em separado para os machos e fêmeas para cada um dos 3 grupos de variáveis, nos quais a variável aleatória (variável dependente) correspondia às condições corporais e as explicativas (variáveis independentes) correspondiam às variáveis dos grupos microhabitat, macrohabitat e relações.
De seguida procedeu-se ao desenvolvimento de um conjunto de modelos com todas as combinações possíveis das variáveis independentes, tendo em conta o modelo global (função
dredge) através do package MuMin (Barton 2015). Estes foram ordenados com base no valor de
AIC (Akaike Information Criteria)(Burnham e Anderson 2002), ou mais concrectamente, o AICc (AIC corrigido para amostras pequenas) uma vez que se tratava de amostras pequenas (Bolker et al. 2009) tendo sido selecionados os modelos com ΔAICc <2 (i.e. diferença entre o menor AIC e o AIC do modelo em análise) que equivalem aos melhores modelos (Burnham e Anderson 2002). Quando mais de um modelo cumpria este critério, foi necessário efetuar uma média dos modelos, usando novamente o package MuMin. Por último, foram calculados os intervalos de confiança (IC a 95%) das variáveis que integram os melhores modelos para determinar para as quais é possível definir a direção (positiva ou negativa) da influência sobre a condição corporal, i.e. aquelas cujos IC não passam o 0.
Depois do cálculo de todos os modelos, criou-se um modelo médio juntando apenas as variáveis significativas para cada nível de habitat e relações, de modo a testar a 4ª hipótese, analisando este modelo médio da mesma maneira que os restantes modelos prévios.
Por fim, analisou-se a robustez do melhor modelo segundo o likelihood ratio test, que analisa a percentagem da variável dependente explicada pelo modelo (Mangiafico 2016).
Tabela I – Variáveis e respectivas abreviaturas usadas nos modelos lineares generalizados
Nome da variável Abreviatura Definição Tipo de
variável
Hipótese % de cobertura de
arbustos
%Arbusto Percentagem de cobertura de arbustos num buffer de 3 metros em redor da armadilha
Contínua Microhabitat
% de cobertura de árvores de fruto
%Fruto Percentagem de cobertura de árvores de fruto num buffer de 3 metros em redor da armadilha
Contínua Microhabitat
% de cobertura de sobreiros
%Sobreiro Percentagem de cobertura de sobreiros num buffer de 3 metros em redor da armadilha
Contínua Microhabitat
% de cobertura de gramíneas
%Gramíneas Percentagem de cobertura de gramíneas num buffer de 3 metros em redor da armadilha
Contínua Microhabitat
% de cobertura de manta morta
%Mtmorta Percentagem de cobertura de manta morta num buffer de 3 metros em redor da armadilha
Contínua Microhabitat
Altura média de arbustos
Altarbustos Altura média de arbustos num buffer de 3 metros em redor da armadilha
11
Altura média de gramíneas
Altgramíneas Altura média de gramíneas num buffer de 3 metros em redor da armadilha
Contínua Microhabitat
% de cobertura de solo
Solo Percentagem de cobertura de solo num buffer de 3 metros em redor da armadilha
Contínua Microhabitat
Densidade do subcoberto
Denso/Disperso/Matos agregados
Densidade do subcoberto com categorias denso, disperso ou agregado
Categórica ordinal
Macrohabitat
Topografia Plano/Declive/Colina/Vale Topografia numa faixa de 50 metros com categorias plano, declive, colina ou vale
Categórica ordinal
Macrohabitat
Heterogeneidade Homogéneo/moderadamente heterogéneo/heterogéneo
Heterogeneidade numa faixa de 50 metros com categorias homogéneo,
moderadamente heterogéneo ou heterogéneo Categórica ordinal Macrohabitat Número de sobreiros
Sobreiros Número de sobreiros numa faixa de 50 metros em redor da armadilha
Contínua Macrohabitat
Número de pinheiros
Pinheiros Número de pinheiros numa faixa de 50 metros em redor da armadilha
Contínua Macrohabitat
Anos de isolamento Anos Anos de isolamento ao pastoreio Categórica nominal
Macrohabitat
Outras espécies Outras espécies Capturas de outras espécies de pequenos mamíferos na armadilha
Contínua Relações
Outros Apodemus Apodemus Outras capturas de Apodemus sylvaticus da armadilha
Contínua Relações
Risco de predação Predação Risco de predação no local de amostragem Categórica nominal
12
3. Resultados
3.1. Capturas
No total dos 15 locais amostrados foram capturados 185 indivíduos de 3 espécies diferentes: dois roedores, Apodemus sylvaticus (N=151, Tabela II) e Mus spretus (N=10, Tabela II); e um soricomorfo, Crocidura russula (N=24, Tabela II). Nos locais de controlo foram detectados um total de 59 indivíduos, enquanto que os restantes se encontravam distribuídos equitativamente pelos locais isolados ao pastoreio: 30 indivíduos nos locais isolados desde 2004, 38 nos de 2008 e finalmente 24 indivíduos nos locais isolados desde 1998. A razão-entre-sexos estimada para
Apodemus sylvaticus foi de 1,22 machos para cada fêmea, enquanto para Mus spretus foi de 4
machos para cada fêmea e para Crocidura russula de 0,17 machos para cada fêmea.
Tabela II - Capturas das diferentes espécies de pequenos mamíferos por local de amostragem. Os acrónimos Ctr, 2008, 2004 e 1998 correspondem aos locais de controlo e aos locais isolados ao pastoreio desde 2008, 2004 e 1998, respectivamente.
Devido a este reduzido número de capturas de Mus spretus e Crocidura russula, bem como o desequilíbrio na razão-entre-sexos, apenas se calculou a condição corporal para Apodemus
sylvaticus.
3.2. Índice de condição corporal da população de Apodemus sylvaticus
Os valores médios de condição corporal de A. sylvaticus por local de amostragem, sugerem a existência de dois grupos distintos (Figura 2)(é importante salientar que as figuras demonstram as medianas e não as médias). Por um lado, os locais onde o pastoreio está ausente desde 1998 e 2008 possuem indivíduos com condições corporais mais elevadas, com médias bastante próximas, 26,76 e 26,51 respetivamente, e sem diferenças significativas entre eles (p >0,05). No sentido
Apodemus sylvaticus Mus spretus Crocidura russula
machos fêmeas machos fêmeas machos fêmeas não determinado
Ctr-01 4 3 0 0 1 0 0 Ctr-04 5 5 5 0 0 2 0 Ctr-06 8 4 0 0 0 0 0 Ctr-P01 4 6 1 0 0 0 0 Ctr-P02 4 6 0 1 0 0 0 Ctr-P03 5 5 0 0 0 0 0 2008-01 6 4 0 0 0 0 0 2008-04 11 5 2 1 0 1 0 2008-07 7 5 0 0 0 0 0 2004-01 5 5 0 0 1 4 2 2004-03 4 6 0 0 0 0 0 2004-04 6 4 0 0 1 9 2 1998-01 5 5 0 0 0 0 0 1998-02 2 1 0 0 0 1 0 1998-05 7 4 0 0 0 1 0
13 oposto, os locais não visitados pelo gado desde 2004 e os locais controlo, ou seja, com pastoreio albergam indivíduos com condições corporais mais reduzidas, sendo as médias destes locais de 22,51 e 22,79, respetivamente, onde igualmente também não há diferenças significativas entre os grupos (p>0,05).
Figura 2 - Boxplot representando a mediana da condição corporal por tipo de local de amostragem. Os acrónimos 1998, 2004 e 2008 correspondem aos locais isolados ao pastoreio desde 1998, 2004 e 2008, respectivamente.
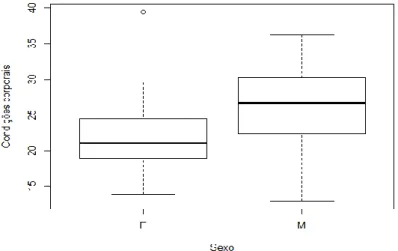
No total das capturas, a condição corporal média foi diferente consoante o sexo do indivíduo, com a condição corporal média dos machos de 26,23 enquanto a das fêmeas foi de 21,98 (Figura 3). Uma vez que a condição corporal era diferente consoante o sexo e existia diferenças significativas entre estes (p<0,05), as hipóteses foram testadas em separado para os machos e para as fêmeas.
Figura 3 - Boxplot representando a mediana da condição corporal por sexo. Os acrónimos F e M correspondem ao sexo feminino e masculino, respectivamente.
14
3.3. Hipóteses explicativas da variação da condição corporal
Nenhuma das variáveis apresentou valores de VIF maiores que 5, razão pela qual todas as variáveis foram usadas nos modelos (Anexo 1 e 2). Para cada sexo foram criados cento e vinte e oito modelos para cada uma das hipóteses do microhabitat e do macrohabitat e oito modelos para a hipótese das relações.
Para os machos, das três hipóteses formuladas sobre as condicionantes à condição corporal de A.
sylvaticus (incluindo apenas variáveis do mesmo tipo: microhabitat; macrohabitat e relações) foi
verificado que aquela composta apenas por variáveis associadas ao microhabitat apresentava suporte estatístico (Tabela III; Anexo 3). De entre este grupo de variáveis, o melhor modelo para esta hipótese (i.e., ΔAICc mais reduzido) inclui as variáveis percentagem de cobertura de arbustos, a percentagem de cobertura de árvores de fruto e a altura média das gramíneas. Os outros cinco modelos com ΔAICc<2 (que podem ser considerados modelos igualmente bons) incluem ainda a altura média de arbustos, a percentagem de cobertura de gramíneas e a percentagem de cobertura de sobreiros (Tabela III).
Para os modelos associados ao macrohabitat foram estimados valores de AICc mais elevados do que os calculados para os modelos de microhabitat. No entanto, existem três modelos da hipótese de macrohabitat com um ΔAICc<2, nos quais apenas o número de sobreiros é comum a todos, sendo que as restantes variáveis incluídas foram as relacionadas com a homogeneidade da vegetação e os anos de exclusão (Tabela III; Anexo 4). Finalmente os modelos da terceira hipótese – Relações – possuíram valores de AICc ainda maiores que aqueles estimados para os modelos da hipótese anterior, sendo que nesta hipótese existem apenas dois modelos com um ΔAICc<2, um dos quais apenas incluí a presença de outras espécies de pequenos mamíferos e o outro sendo o modelo nulo desta hipótese, ou seja modelo que não inclui nenhuma variável (Tabela III; Anexo 5).
Tabela III - Modelos lineares generalizados para as diferentes hipóteses explicativas da variação da condição corporal dos machos. “DF” graus de liberdade, “LogLik” probabilidade logarítmica do modelo, “AICc” Akaike Information Criteria corrigido para amostras pequenas, “ΔAICc” diferença entre o AICc do modelo e o AICc mais baixo, “W” peso do modelo. Os acrónimos desta tabela estão explicados na Tabela I.
DF LogLik AICc ΔAICc W
H1 - Microhabitat Altgramíneas+%Arbustos+%Fruto 5 -237,463 485,7 0,00 0,09 Altgramíneas+%Arbustos+%Fruto+%Sobreiro 6 -236,346 485,8 0,10 0,086 Altarbustos+ Altgramíneas+%Arbustos+%Fruto 6 -236,376 485,9 0,16 0,083 Altarbustos+ Altgramíneas+%Arbustos+%Fruto+%Sobreiro 7 -235,738 487,0 1,28 0,048 %Arbustos+%Fruto+%Gramíneas+%Sobreiro 6 -236,946 487,0 1,30 0,047 %Arbustos+%Fruto+%Gramíneas 5 -238,462 487,0 2,00 0,033 H2 - Macrohabitat Denso+Disperso+Sobreiros 5 -239,858 490,5 0,00 0,205 Sobreiros 3 -242,344 491,0 0,49 0,160 Anos+Denso+Disperso+Sobreiros 6 -239,208 491,5 1,03 0,122 H3 - Relações Modelo nulo 2 -250,065 504,3 0,00 0,299 Outras espécies 3 -249,088 504,5 0,20 0,271
15 O modelo médio para a hipótese com mais suporte para os machos – microhabitat - revelou que as variáveis relevantes e com influência positiva (intervalo de confiança a 95% apenas com valores positivos) eram a percentagem de cobertura de arbustos, a percentagem de cobertura de árvores de fruto e a altura média das gramíneas (Tabela IV).
Tabela IV – Modelo médio para a hipótese com mais suporte para os machos – microhabitat – com intervalos de confiança de 95% para cada variável. “β” coeficiente covariado da variável, “SE” erro padrão, “Z-value” estatística de Wald para testar a hipótese do coeficiente ser 0, “p” p-value, “95% IC” intervalo de confiança de 95 %. Os acrónimos desta tabela estão explicados na Tabela I.
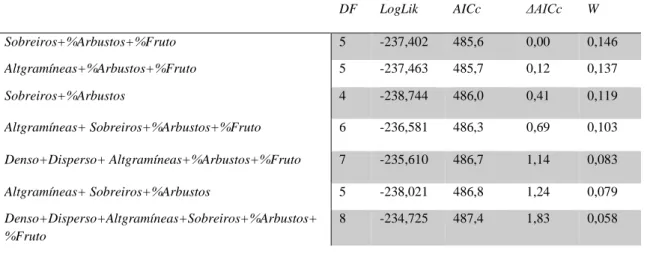
Numa última fase de análise dos machos, para testar a quarta hipótese – Hipótese mista, utilizou-se as variáveis incluídas nos melhores modelos de cada uma das três hipóteutilizou-ses iniciais, mas cujo intervalo de confiança dos seus coeficientes não incluía o zero, para garantir que o seu efeito era direccional. Assim, foram construídos trinta e dois modelos com as combinações de cinco variáveis: percentagem de cobertura de arbustos, número de sobreiros, percentagem de árvores de fruto, altura média de gramíneas e densidade do subcoberto (Tabela III). Destes, apenas sete (Tabela V; Anexo 6) foram considerados melhores modelos (i.e., ΔAICc<2), atingindo um AICc menor que aquele estimado para os melhores modelos das três hipóteses anteriores (Tabela III e V). Assim, esta hipótese foi considerada como tendo maior suporte estatístico. O likelihood ratio
test indicou que o melhor modelo respondia a 26,6% da variação da condição corporal dos
machos.
Tabela IV - – Modelos lineares generalizados para a hipótese mista para os machos. “DF” graus de liberdade, “LogLik” probabilidade logarítmica do modelo, “AICc” Akaike Information Criteria corrigido para amostras pequenas, “ΔAICc” diferença entre o AICc do modelo. Os acrónimos desta tabela estão explicados na Tabela I.
β SE Z-value p 95% IC
Intersecção 20,20 1,88 10,61 <0,001 16,468 23,927
%Arbustos 0,12 0,03 4,45 <0,001 0,067 0,174
%Fruto 0,28 0,12 2,32 0,020 0,045 0,523
Altgramíneas 0,04 0,03 1,39 0,164 0,006 0,089
DF LogLik AICc ΔAICc W
Sobreiros+%Arbustos+%Fruto 5 -237,402 485,6 0,00 0,146 Altgramíneas+%Arbustos+%Fruto 5 -237,463 485,7 0,12 0,137 Sobreiros+%Arbustos 4 -238,744 486,0 0,41 0,119 Altgramíneas+ Sobreiros+%Arbustos+%Fruto 6 -236,581 486,3 0,69 0,103 Denso+Disperso+ Altgramíneas+%Arbustos+%Fruto 7 -235,610 486,7 1,14 0,083 Altgramíneas+ Sobreiros+%Arbustos 5 -238,021 486,8 1,24 0,079 Denso+Disperso+Altgramíneas+Sobreiros+%Arbustos+ %Fruto 8 -234,725 487,4 1,83 0,058
16 Das cinco variáveis que compõem os melhores modelos da hipótese mista (Tabela V) apenas a percentagem de cobertura de arbustos apresentou um intervalo de confiança dos seus coeficientes não incluía o zero. Assim, apenas esta variável foi considerada influente, uma vez que apenas para ela foi possível aferir a direção (neste caso positiva) do seu efeito (Tabela VI).
Tabela V - Modelo médio para a hipótese mista para os machos, com intervalos de confiança de 95% para cada variável. β” coeficiente covariado da variável, “SE” erro padrão, “Z-value” estatística de Wald para testar a hipótese do coeficiente ser 0, “p” p-value, “95% IC” intervalo de confiança de 95 %. Os acrónimos desta tabela estão explicados na Tabela I. Β SE Z-value P 95% IC Interseção 22,72 3,31 6,815 <0,001 16,188 29,257 Sobreiros -0,01 0,01 1,065 0,287 -0,039 0,001 %Arbustos 0,09 0,03 2,880 0,003 0,029 0,154 %Fruto 0,17 0,15 1,126 0,260 -0,018 0,497 Altgramíneas 0,02 0,02 0,914 0,360 -0,009 0,087 Denso 0,20 0,95 0,210 0,833 -2,838 4,928 Disperso 0,52 1,31 0,396 0,692 -0,794 6,154
A percentagem de cobertura de arbustos era maior nos locais com maior condição corporal (Figura 4). Por outro lado, a média de percentagem de cobertura de arbustos era mais elevada nos locais excluídos ao pastoreio desde 1998 (60%) e nos locais excluídos desde 2008 (52,32%), ao passo que nos outros locais a média era mais baixa, sendo de 43,15% para os locais de 2004 e de apenas 31,29% para os locais de controlo (Figura 4).
Figura 4 - Boxplot representando a mediana da variação da percentagem de cobertura de arbusto por tipo de local de amostragem. Os acrónimos 1998, 2004, 2008 e Controlo correspondem aos locais isolados ao pastoreio desde 1998,
17 Para o caso das fêmeas verificou-se que novamente a hipótese composta apenas por variáveis associadas ao microhabitat apresentava suporte estatístico (Tabela VII). De entre o grupo de variáveis, o melhor modelo para esta hipótese (i.e., ΔAICc mais reduzido) inclui as variáveis altura média das gramíneas e a percentagem de cobertura de gramíneas. Os outros dois modelos com ΔAICc<2 incluem ainda a percentagem de cobertura de arbustos e a percentagem de cobertura de manta morta (Tabela VII; Anexo 7).
Para os modelos associados ao macrohabitat foram estimados valores de AICc mais elevados do que os calculados para os modelos de microhabitat. No entanto, existem dois modelos da hipótese de macrohabitat com um ΔAICc<2, nos quais o número de sobreiros é comum a todos, sendo que as restantes variáveis incluídas foram as relacionadas com a topografia (Tabela VII; Anexo 8). Finalmente os modelos da terceira hipótese – Relações – possuíram valores de AICc ainda mais elevados que aqueles estimados para os modelos da hipótese anterior (Tabela VII; Anexo 9), sendo que nesta hipótese existem apenas dois modelos com um ΔAICc<2, um dos quais apenas incluí a presença de outros indivíduos de Apodemus sylvaticus e o outro sendo o modelo nulo desta hipótese.
Tabela VI - Modelos lineares generalizados para as diferentes hipóteses explicativas da variação da condição corporal das fêmeas. “DF” graus de liberdade, “LogLik” probabilidade logarítmica do modelo, “AICc” Akaike Information Criteria corrigido para amostras pequenas, “ΔAICc” diferença entre o AICc do modelo e o AICc mais baixo, “W” peso do modelo. Os acrónimos desta tabela estão explicados na Tabela I.
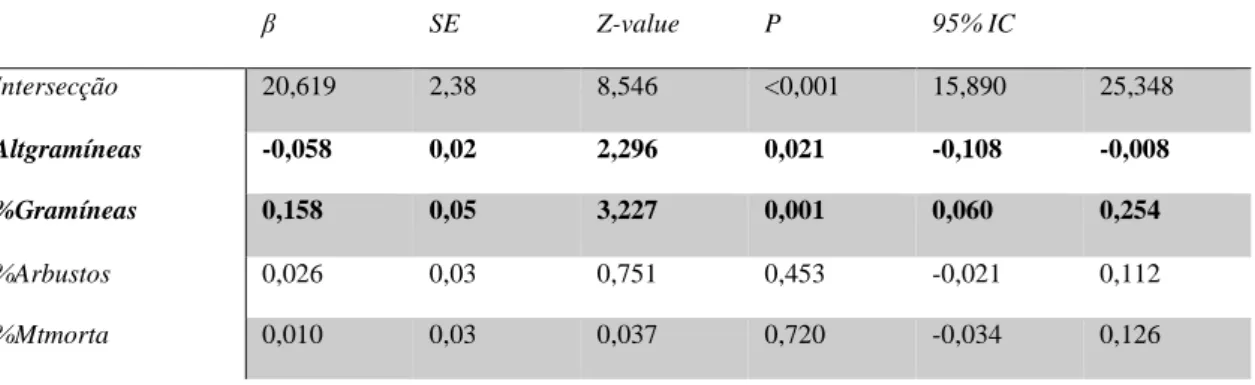
O modelo médio para a hipótese com mais suporte para as fêmeas – microhabitat - revelou que a variável relevante e com influência positiva (intervalo de confiança a 95% apenas com valores positivos) era a percentagem de cobertura de gramíneas e que a variável relevante mas com influência negativa (intervalo de confiança a 95% apenas com valores negativos) era a altura média das gramíneas (Tabela VIII).
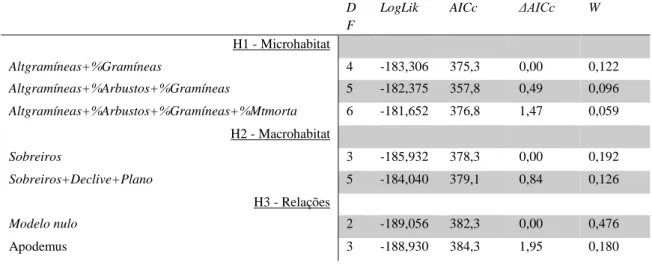
D F
LogLik AICc ΔAICc W
H1 - Microhabitat Altgramíneas+%Gramíneas 4 -183,306 375,3 0,00 0,122 Altgramíneas+%Arbustos+%Gramíneas 5 -182,375 357,8 0,49 0,096 Altgramíneas+%Arbustos+%Gramíneas+%Mtmorta 6 -181,652 376,8 1,47 0,059 H2 - Macrohabitat Sobreiros 3 -185,932 378,3 0,00 0,192 Sobreiros+Declive+Plano 5 -184,040 379,1 0,84 0,126 H3 - Relações Modelo nulo 2 -189,056 382,3 0,00 0,476 Apodemus 3 -188,930 384,3 1,95 0,180
18
Tabela VII - Modelo médio para a hipótese com mais suporte para as fêmeas – microhabitat – com intervalos de confiança de 95% para cada variável. β” coeficiente covariado da variável, “SE” erro padrão, “Z-value” estatística de Wald para testar a hipótese do coeficiente ser 0, “p” p-value, “95% IC” intervalo de confiança de 95 %. Os acrónimos desta tabela estão explicados na Tabela I.
À semelhança do referente aos machos, para testar a quarta hipótese – Hipótese mista, foram construídos oito modelos com as combinações de três variáveis: altura média de gramíneas, percentagem de cobertura de gramíneas e número de sobreiros (Tabela VII). Destes, apenas dois (Tabela IX; Anexo 10) foram considerados melhores modelos (i.e., ΔAICc<2), atingindo um AICc menor que aquele estimado para os melhores modelos das três hipóteses anteriores (Tabela 7 e 9). Assim, esta hipótese foi considerada como tendo maior suporte estatístico. O likelihood
ratio test indicou que o melhor modelo respondia a 21,5% da variação da condição corporal das
fêmeas.
Tabela VIII - Modelos lineares generalizados para a hipótese mista para as fêmeas. “DF” graus de liberdade, “LogLik” probabilidade logarítmica do modelo, “AICc” Akaike Information Criteria corrigido para amostras pequenas, “ΔAICc” diferença entre o AICc do modelo e o AICc mais baixo, “W” peso do modelo. Os acrónimos desta tabela estão explicados na Tabela I.
As três variáveis integram os melhores modelos da hipótese mista (Tabela X) e todas apresentaram um intervalo de confiança dos seus coeficientes não incluía o zero. Assim, as variáveis altura média de gramíneas e número de sobreiros foram consideradas influentes, com influência negativa, enquanto que a percentagem de cobertura parece influenciar positivamente (Tabela X). β SE Z-value P 95% IC Intersecção 20,619 2,38 8,546 <0,001 15,890 25,348 Altgramíneas -0,058 0,02 2,296 0,021 -0,108 -0,008 %Gramíneas 0,158 0,05 3,227 0,001 0,060 0,254 %Arbustos 0,026 0,03 0,751 0,453 -0,021 0,112 %Mtmorta 0,010 0,03 0,037 0,720 -0,034 0,126
DF LogLik AICc ΔAIC
c
W
Altgramíneas+Sobreiros+%Gramíneas 5 -181,189 373,4 0,00 0,601
19
Tabela IX - Modelo médio para a hipótese mista para as fêmeas, com intervalos de confiança de 95% para cada variável. β” coeficiente covariado da variável, “SE” erro padrão, “Z-value” estatística de Wald para testar a hipótese do coeficiente ser 0, “p” p-value, “95% IC” intervalo de confiança de 95 %. Os acrónimos desta tabela estão explicados na Tabela I. Β SE Z-value P 95% IC Interseção 23,937 1,65 14,334 <0,001 20,664 27,210 Altgramíneas -0,063 0,02 2,620 0,009 -0,110 -0,016 Sobreiros -0,013 0,01 1,161 0,246 -0,035 -0,0002 %Gramíneas 0,137 0,04 2,937 0,003 0,046 0,228
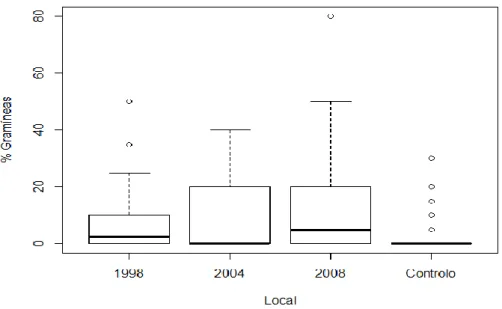
Para a percentagem de cobertura de gramíneas (Figura 5), os locais com maior percentagem eram os sem pastoreio sem 2008, com uma média de 13 %, seguidos dos de 2004, com uma média de 10 %, e dos de 1998, com 8.5 % de média. Nos locais de controlo, as gramíneas eram praticamente inexistentes, cobrindo apenas uma média de 2.5 %.
Figura 5 - Boxplot representando a mediana da variação da percentagem de cobertura de gramíneas por tipo de local de amostragem. Os acrónimos 1998, 2004, 2008 e Controlo correspondem aos locais isolados ao pastoreio desde
1998, 2004 e 2008 e aos locais de controlo, respectivamente.
Para a altura média de gramíneas (Figura 6), os locais de controlo eram claramente aqueles com a altura média mais baixa, de apenas 5,58 centímetros. Por outro lado, os locais isolados desde 2008 eram aqueles com a maior altura média, de 27,09 centímetros. Os locais isolados desde 1998 e 2004 possuíam gramíneas com alturas médias de 17,77 centímetros e 14,13 centímetros, respectivamente.
20
Figura 6 - Boxplot representando a mediana da variação da altura média de gramíneas por tipo de local de amostragem. Os acrónimos 1998, 2004, 2008 e Controlo correspondem aos locais isolados ao pastoreio desde 1998,
2004 e 2008 e aos locais de controlo, respectivamente.
Em relação ao número de sobreiros (Figura 7), os locais de controlo são aqueles com a média mais elevada (168 sobreiros), enquanto que em sentido oposto, os locais isolados desde 1998 possuíam a menor quantidade de sobreiros, com uma média de apenas 47 sobreiros. Os locais com isolamento desde 2008 e desde 2004 possuíam médias de 109 e 134 sobreiros, respetivamente.
Figura 7 - Boxplot representando a mediana da variação do número de sobreiros por tipo de local de amostragem. Os acrónimos 1998, 2004, 2008 e Controlo correspondem aos locais isolados ao pastoreio desde 1998, 2004 e 2008 e aos
21
4. Discussão
4.1. Condição corporal vs exclusão pastoreio
Apesar de não terem sido detectadas diferenças significativas entre os locais com diferentes histórias de pastoreio, algumas tendências parecem emergir. Os grupos de locais com isolamento ao pastoreio desde 1998 e desde 2008 são aqueles em que registou-se maiores condições corporais, enquanto que os locais de controlo e os locais isolados ao pastoreio desde 2004 são aqueles onde foram encontradas as menores condições corporais. O facto do aumento do período de exclusão do pastoreio não ser directamente proporcional ao aumento da condição corporal, sugere que esta última poderá depender de algo externo à escala temporal associada à ausência do impacto do pastoreio. A complexidade do habitat poderá não depender do tempo de isolamento ao pastoreio mas sim de outro factor mais ecológico, como o tipo de solo ou outra variável não avaliada que influencie o crescimento da vegetação, tornando o habitat mais ou menos adequado para os pequenos mamíferos. O facto de os conjuntos de locais estarem emparelhados dois a dois também pode sugerir que a variação das condições corporais é uma variação cíclica. Assim, esta variação aparenta não depender do período de exclusão, como poderia ser expectável, mas sim de alguma variação climática ou ecológica cíclica que influencia a disponibilidade de recursos, influenciando assim a condição corporal dos indivíduos.
4.2. Hipóteses explicativas da variação individual da condição corporal
Das 3 primeiras hipóteses apontadas como possíveis explicações para as variações detectadas na condição corporal, tanto para os machos como para as fêmeas, a hipótese associada às variáveis de microhabitat parece ser aquela melhor suportada pelos dados. Porém, os modelos integradores, provaram ser os que melhor explicam a variabilidade detetadas na condição corporal. Tal sugere em efeito sinergístico do habitat e da estrutura das comunidades na condição corporal em
Apodemus sylvaticus. Este resultado do modelo misto contribui para o debate sobre qual a escala
de habitat mais importante, em termos de conservação, para os pequenos mamíferos. Alguns autores sugerem que a estrutura do microhabitat é o que determina os padrões de distribuição e abundância (Bellows et al. 2001; Jorgensen 2004; Traba et al. 2009; Tarjuelo et al. 2011) enquanto que outros apoiam a hipótese do macrohabitat ser o principal responsável por estas variações (Morris 1984, 1987; Orrock et al. 2000; Corbalán 2006). Uma vez que as condições corporais de
Apodemus sylvaticus são influenciadas por variáveis tanto de microhabitat como de macrohabitat,
estes roedores aparentam depender igualmente das diferentes escalas do habitat para satisfazer os seus requisitos ecológicos. Estes resultados indiciam que, em termos de condição corporal, o habitat deve ser considerado a várias escalas, pois as variáveis associadas podem estar a codificar diferentes recursos ou estratégias ecológicas tais como procura de alimento e refúgio (para o caso do microhabitat, Traba et al. 2009; Tarjuelo et al. 2011) ou patrulhamento do território (para o caso do macrohabitat, Wolton 1985; Rosalino 2011).
4.3. Variáveis importantes para a condição corporal
Segundo estudos anteriores, a selecção de habitat pelos pequenos mamíferos, em particular no caso de Apodemus sylvaticus, é maioritariamente dependente da abundância de alimento e da disponibilidade de abrigo (Traba et al. 2009; Tarjuelo et al. 2011), maioritariamente associados à presença de arbustos (Boitani 1985; Khidas et al. 2002; Torre et al. 2002; Michel 2007). No entanto, esta espécie possui hábitos cursoriais e arborícolas, podendo mesmo estabelecer as suas