The first author of the publication (Saara Laulumaa) performed most of the wet lab experiments. The author participated in the process of writing the paper by contributing to the methodology and results of the molecular dynamics simulations.
![Figure 1.1 A schematic drawing of the interior of a cell. Figure modified from Ref. [1].](https://thumb-eu.123doks.com/thumbv2/9pdfco/1889886.266052/22.748.149.553.90.342/figure-schematic-drawing-interior-cell-figure-modified-ref.webp)
Research objectives and scope of the Thesis
By collaborating with a group of experimental wet labs, our goal was to create a broad and thorough picture of the dynamics of P2 and several of its point-mutated variants in order to further research in diseases of the peripheral nervous system. Our second protein target discussed in this thesis is rhodopsin, a well-studied G protein-coupled receptor found in the retina of the human eye.
Structure of the dissertation
In the seventh chapter, we discuss the results of all four papers included in this thesis. The final chapter summarizes the research conducted for this thesis and offers ideas for future prospects.
Cellular life
Lipids
The first part of the name (PO) defines the tail group - the first hydrocarbon tail is a palmitic acid and the second an oleic acid. Oleic acid is also classified as an omega-9 fatty acid because of the double bond on the ninth bond from the tip of the tail.
![Figure 2.1 Schematic picture of a cellular membrane. Figure adapted from Ref. [23].](https://thumb-eu.123doks.com/thumbv2/9pdfco/1889886.266052/29.748.118.660.86.365/figure-schematic-picture-cellular-membrane-figure-adapted-ref.webp)
Proteins
Integral membrane proteins are mostly bound to the interior of the membrane with parts of the protein protruding from the surface of the membrane. These receptors can bind neurotransmitters—chemicals created in the synapses of the nervous system—that activate the protein.
Point mutations – an issue in protein encoding
There are three different stop codons that are recognized by the ribosome and signal the end of the protein. In this part of the thesis, we focus on the central part of the nervous system.
The nervous system
The basic elements of the nervous system are nerve cells or neurons (see Fig. 3.1). The PNS consists of all parts of the nervous system that are connected to the CNS.
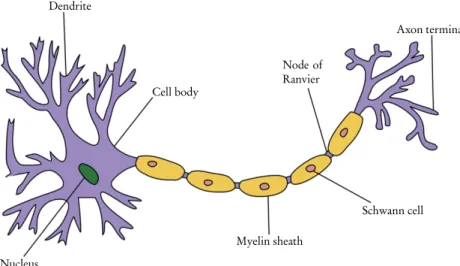
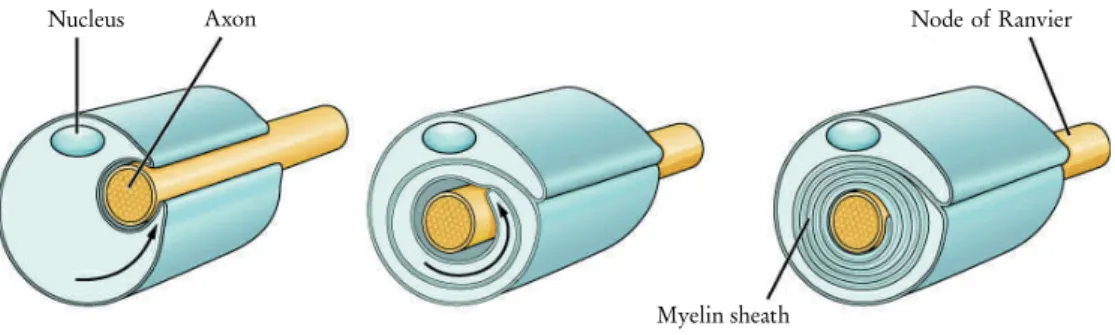
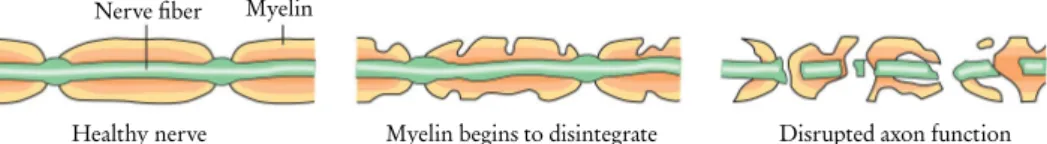
The myelin sheath
In the PNS, the myelin sheath is made by Schwann cells, which interact with only a single axon. As a result, both axon and myelin sheath damage in the CNS is much slower to heal.
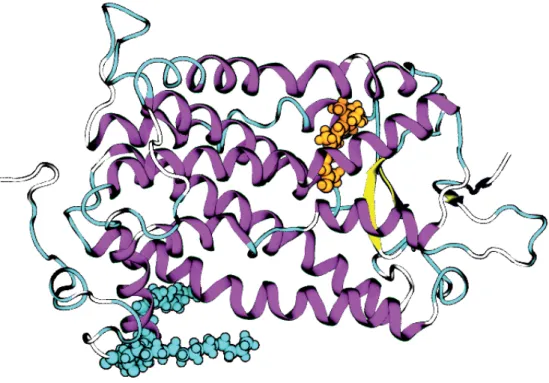
The peripheral membrane protein P2
The inability to degrade damaged myelin appears to be due to a lack of autophagy activation in the CNS[67]. Other factors that impede nerve healing in the CNS are due to oligodendrocytes inhibiting neuronal growth and the absence of growth factor chemicals [56].
Diseases of the nervous system
A common factor in MS patients is the destruction of the myelin sheath and the loss of oligodendrocytes [80]. Most variants of CMT cause demyelination in the myelin sheath, but some can also cause damage to the axon itself.
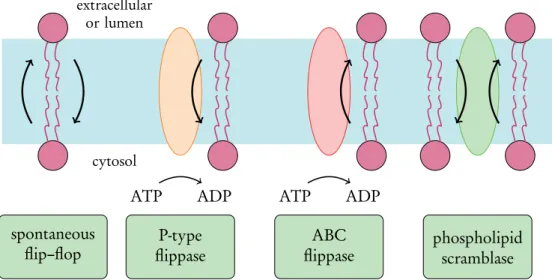
Transbilayer lipid motion
For the ER to grow properly, half of the synthesized lipids must flip-flop across the ER bilayer to the lumenal side[98]. Other relevant factors include the phase behavior of the bilayer[107] and the concentration of membrane packing elements such as cholesterol[108].
Scramblase proteins
Another recent study revealed a correlation between the decrease in POPS flip-flop free energy with increased concentrations of peroxidized POPC molecules in a similar system [115]. The second is the so-called credit card model, where the lipid head group enters the hydrophilic transmembrane channel of the scramblaza, possibly via a conformational change in the protein, and exits on the second leaflet [127].
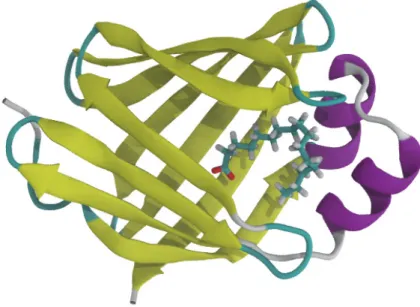
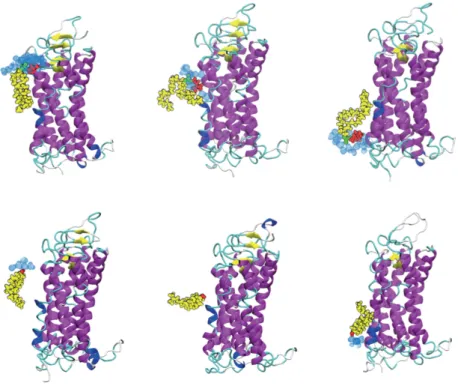
Rhodopsin
Two palmitate chains attached to cytosolic cysteine residues are also shown in cyan. The extracellular side is home to the N-terminal along with some short interhelical loops and beta sheets.
Scramblase dysfunction in diseases
Due to the high resolution with which the crystal structure of rhodopsin was obtained, it is possible to build a complete model of the protein by knowing the exact location of each individual atom. Suppression of apoptotic signals in the early stages of cancer due to mutations in scramblase or other tumor suppressor genes is often the reason that cancer cells can survive [133].
Building the model
First, we create a model of the system under study, then let it operate under environmental conditions of our choice for a certain time. Perhaps the easiest model to understand is the classical atomistic model, where each atom of the system is assigned a value for its mass and electric charge.
Force fields and topologies
Similarly, if they are on opposite sides of the bond, the dihedral angle is 180 degrees, corresponding to a trans configuration. The strength of the coupling can be adjusted by changing the time constant, which can be useful, for example, for a fast equilibration of the system. The pressure is directed to the reference pressure in advance by the matrix parameter W, which defines the strength of the coupling.
The pressure is presented as a 3×3 tensor, which allows non-isotropic handling of the system pressure, which is desirable in the case of lipid bilayers. To avoid artifacts caused by the sides of the box, periodic boundary conditions (PBCs) are often introduced in the simulations. Periodic boundary conditions place an infinite number of identical copies of the simulated system around themselves.
Analysis methods
Free energy and umbrella sampling
Thermodynamic free energy is a useful quantity for measuring thermal and chemical processes since every chemical reaction involves a change in the free energy of the system. Temperature is important in free energy reactions – the entropic term plays a larger role at higher temperatures and is not as significant at lower temperatures. The enthalpic component can be associated with the heat of the system and is defined as.
This can be performed with a centered difference method applied to the entropic part of the free energy profile. In the third paper, we investigated another point mutant of the P2 protein, F57A through atomistic MD simulations. The windows were equilibrated for 20 ns followed by a 50 ns data acquisition period—the equilibration period long enough for the system to reverse the effects caused by the forced direction of the lipid and the data period short enough for the system not to relax. completely in its current state.
Point mutation in P2 lid hinge increases protein dynamics
In Paper I, we constructed and simulated models of the P2 protein and its P38G mutant with and without a bound ligand found in the crystallized structure we used as a base model[11]. The color bar indicates the RMSF values, with bright magenta corresponding to the most flexible parts of the protein. By comparing the protein variants, we can see that the dynamics of the ligand-free P2 and its P38G mutant are quite similar, with perhaps a slight decrease in flexibility upon mutation.
Second, the data show that the wild type with the palmitate is significantly more stable than either of the two palmitate-free variants, especially in the loop regions. The increased flexibility of the P38G mutant could explain the previously observed increase in membrane binding activity of the variant. For this reason, we performed additional simulations where the main focus was the dynamics of the protein hydrogens, divided into hydrogens of both CH3 and CH2 groups.

CMT1–associated point mutations affect P2 dynamics
The empty protein is shown in the top graph and the P2 gel ligand at the bottom. In general, the number of water molecules found in the vessel was smaller in the presence of the ligand. The only exception was the T51P mutant, where the number of water molecules was much higher in the ligand form.
This was explained by the early opening of the T51P structure, allowing more water to enter the barrel (see Figure 7.5). The β5 and β6 strands of the barrel flip away from the protein creating a gap large enough for a ligand to pass through. This movement can also be seen in the RMSF plots in the area of higher fluctuations.

P2 barrel opening is regulated by a conserved residue in the portal
Bottom: The closed and open structures of the F57A mutant of P2 taken from the simulations. By comparing the dynamic cross-correlation matrices of the wild type P2 with its point mutants, we can see which interactions are disrupted by the point mutations. The dynamic cross-correlation maps (Figure 7.7) for the wild type show areas of significant anti-correlation between the β5-β6 loop (residues 74-77) and the lid region (residues 16-36) and the adjacent β3-β4 loop (residues 55-58) indicated by the two arrows.
We believe that this anti-correlation reflects the coordinated open-close movements of the β5-β6 flap upon ligand transfer. The DCCM analysis also shows that upon point mutation these regions of anti-correlation disappear, indicating that the mutation removes the ability of the protein to regulate its opening. Upon further examination of the previously studied point mutants (P38G and the CMT disease . mutants) with higher dynamics in the portal region, we also perceived similar changes through their dynamic cross-correlation maps.

Rhodopsin functions as a non-selective lipid scramblase
Molecular dynamics simulations confirmed the enhanced dynamics of the mutant variant, particularly within the portal region in the presence of bound fatty acid. Note how the main chain hydrogen bond (dashed lines) is preserved at the edge of the β-sheet also in the mutant. The most flexible parts of P2 can be observed to reside on the side of the lid, i.e.
The temperature dependence of the elastic scattering intensity, S(Q,ω= 0), of wtP2, normalized to the 20 K temperature data, is reported in Fig. 4 with fits using the bimodal distribution model (Eq 4) and the model Gaussian. (Ex. 1). Temperature dependence of MSD. Data from wtP2 (red) and P2-P38G (blue) were estimated by fitting data obtained on IN6 (top), IN13 (middle), and IN16 (bottom) using eq 1. Conversely, in proteins with bound palmitate, the hydrogens of the P38G mutant are more mobile than in wt-P2.
This agrees well with the clear change in the slope of the sum of elastic intensities (Fig. 5) at 240 K. P2wt and the I43N and I52T mutants all bound to membranes with affinities in the range of 2–6 μM, regardless of lipid composition (Table 3). Structure and dynamics of a portal region mutant of human myelin protein P2 suggests β-barrel opening in fatty acid-binding proteins.
Structure and dynamics of a mutant of the portal region of human myelin protein P2 shows opening of the β-barrel in the fatty acid.
Dynamics of the peripheral membrane protein P2 from human myelin
Molecular mechanisms of Charcot-Marie-Tooth neuropathy linked
Structure and dynamics of a human myelin protein P2 portal region