LES DONNÉES PROVIENNENT DE LA BASE D'EXPRESSION GÉO (NOS D'ACCESSION : GSE33 ET GSE516)..122 FIGURE A.2.8 EXEMPLE D'UN PLAN DE COMPARAISON DIRECTE DE DEUX. FONCTION DES NIVEAUX D'EXPRESSION ESTIMÉS PAR ANALYSE DE VARIANCE..230 FIGURE C.3.9 REPRÉSENTATION DES FRÉQUENCES DE NIVEAUX.
La symbiose
- Généralités
- Une origine endocytobiotique pour la cellule eucaryote
- Endocytobiose et unité de sélection
- Impact sur l’hôte
- Impact sur le symbiote
- Impact sur le symbiocosme
- Le concept d’individu
Elle concerne l'ensemble de la population (contrairement aux précédentes), qui se caractérise par l'amélioration du fonctionnement de l'hôte et la localisation précise des symbiotes (symbioses racinaires de légumineuses). L’absence de recombinaison génétique dans les populations d’endocytobiote est, d’une part, une conséquence de la séquestration de symbiotes dans celles-ci.
Les deux partenaires de l’association puceron - bactérie.52
Caractéristiques biologiques
Le puceron du pois Acyrthosiphon pisum (voir Figure A.1.1) appartient à la famille des Aphididae et à l'ordre des Hémiptères. Ce cycle complexe lui permet d'exploiter de manière optimale les ressources de sa plante hôte (Baumann et Baumann, 1994).
Nutrition
Les pucerons utilisent principalement le saccharose comme source d'énergie (Febvay et al., 1995 ; Rhodes et al., 1996). Si le puceron utilise le saccharose comme source d'énergie, les acides aminés sont quant à eux réservés à la production de composés structuraux (Rhodes et al., 1996).
Localisation des symbiotes
Par conséquent, les pucerons parthénogénétiques qui portent des embryons dans leurs ovaires possèdent à la fois des bactériocytes dans l'hémolymphe maternelle et des bactériocytes dans les embryons. De la naissance jusqu'au milieu de la période de reproduction du puceron, le nombre de bactéries d'origine embryonnaire dépasse même celui d'origine maternelle et peut représenter jusqu'à 75 % de la population totale (Humphreys et Douglas, 1997 ; Whitehead et Douglas, 1993a). .

Du côté de la bactérie Buchnera aphidicola
Un génome avec une organisation originale
Cette étude suggère que la taille du génome de Buchnera continue de diminuer, contrairement à l'hypothèse précédente de stase génomique (Silva et al., 2001). Le génome de Buchnera avec un taux de GC de seulement 26,3 % est l'un des génomes présentant la plus grande distorsion en bases A et T (Clark et al., 1999).
Un métabolisme essentiellement dédié à la biosynthèse
Voies essentielles à la biosynthèse des acides aminés : L'enzyme ilvE (en vert) est absente des voies de biosynthèse de la valine, de l'isoleucine et de la leucine, mais une autre aminotransférase se charge sans aucun doute de ces étapes. Dans la voie de biosynthèse de la lysine, le gène dapC (en bleu) n’a été identifié dans aucun organisme. Enfin, la dernière étape de la biosynthèse de la phénylalanine est catalysée par TyrB chez Escherichia coli mais peut être remplacée par HisC chez Buchnera.
L'opéron NADH déshydrogénase (nuo) et l'opéron du cytochrome C (cyo) sont parfaitement conservés, mais la voie de biosynthèse de l'ubiquinone est absente. Cependant, seuls 18 gènes, soit 3 % du génome bactérien, sont annotés comme transporteurs (voir tableau A.1.1).

Une régulation de l’expression des gènes plutôt
Les conséquences de la perte de gènes régulateurs devraient également être atténuées en fonction du rôle de l'hôte dans la relation endocytobiotique. C’est pourquoi des études théoriques ont été réalisées pour étudier de manière plus spécifique la régulation de la synthèse des acides aminés. De plus, les gènes de biosynthèse des acides aminés sont fortement exprimés chez Buchnera (Baumann et al., 1999).
Ainsi, le contrôle de l’expression des gènes de biosynthèse des acides aminés reste encore un mystère chez Buchnera. Ce surenroulement joue un rôle important dans les modifications de l'expression des gènes chez Escherichia coli au cours de la croissance (Kusano et al., 1996) ou du stress osmotique (Higgins et al., 1988).
Le couple du point de vue de la problématique du métabolisme
Des études nutritionnelles
La première étude réalisée sur le puceron Myzus persicae (Mittler, 1971) a montré que les pucerons symbiotiques tolèrent toutes les carences sauf l'absence de méthionine en raison de ses propriétés phages-stimulantes. En revanche, les pucerons aposymbiotiques ont besoin de tous les acides aminés essentiels, démontrant le rôle des bactéries symbiotiques dans la fourniture d’acides aminés essentiels. La croissance des larves de pucerons a ensuite été étudiée chez le puceron Acyrthosiphon pisum sur un milieu alors déséquilibré en acides aminés essentiels.
Les résultats montrent que les larves aposymbiotiques croissent deux fois plus vite sur un milieu équilibré (c'est-à-dire sur un milieu dont la composition assure une croissance normale des pucerons) tandis que les larves symbiotiques ont une croissance identique sur les deux milieux (Prosser et Douglas, 1992). Cependant, même sur un milieu parfaitement équilibré, les performances des pucerons aposymbiotiques et symbiotiques ne sont jamais comparables, suggérant que la compensation du milieu nutritif ne peut remplacer Buchnera (Prosser et al., 1992).
Des études métaboliques
Une expérience réalisée sur un milieu contenant du saccharose marqué au 14C montre que chez les pucerons symbiotiques, la radioactivité est incorporée dans tous les acides aminés qui composent les protéines. Or, la même étude réalisée sur les pucerons aposymbiotiques montre que les acides aminés essentiels ne contiennent pas de 14C radioactif. Les résultats complets de cette étude indiquent que, chez les pucerons symbiotiques, le saccharose participe à la synthèse de presque tous les acides aminés (sauf l'histidine et l'arginine) via les voies de glycolyse (Febvay et al., 1999).
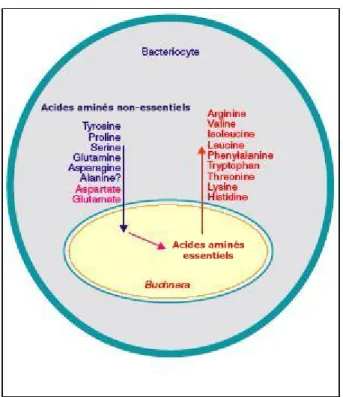
La bactérie est capable de synthétiser des acides aminés essentiels à son hôte, mais elle a besoin de plusieurs acides aminés non essentiels, principalement le glutamate et l'aspartate, qui sont probablement importés du cytoplasme de l'hôte (Zientz et al., 2001). Buchnera est capable de synthétiser des acides aminés essentiels à son hôte (rouge) et semble nécessiter plusieurs acides aminés non essentiels (bleu).
Un couple ou un mariage à trois ?
Parmi ces acides aminés, les deux principaux sont l'aspartate et le glutamate (en rose) (d'après Zientz et al., 2001). L'infection par PASS améliore la reproduction des pucerons dans des conditions de stress thermique (Montllor et al., 2002), ce qui suggère que des bactéries secondaires sont capables de compenser l'absence de Buchnera lorsque la température est élevée. La fonction des bactéries secondaires serait alors de favoriser la survie des pucerons durant la saison estivale (Koga et al., 2003).
Des études ont montré que les pucerons rendus aposymbiotiques présentent une extraordinaire prolifération de micro-organismes (Nakabachi et al., 2003) et que l'infection par des symbiotes secondaires permet la survie et la reproduction des pucerons aposymbiotiques sur plusieurs générations (Koga et al., 2003). Ils ont présenté comme preuve la grande quantité de symbiotes secondaires dans ces lignées de pucerons (Chen et al., 2000).

Les objectifs de la thèse du point de vue biologique
Il est tentant de comparer ce phénomène à celui de la duplication génique, qui confère au gène dupliqué la capacité d'évoluer vers de nouvelles fonctions car les contraintes fonctionnelles restent assumées par le gène d'origine. De même, dans le système super symbiotique, les symbiotes secondaires ouvrent une ouverture vers de nouvelles fonctions biologiques tandis que le symbiote primaire est préservé. Cependant, les progrès du séquençage associés à la diminution des coûts de l'expérimentation (Gibson, 2002) ont permis l'apparition récente de puces à ADN dans tous les domaines de la biologie, notamment dans ceux de la physiologie et de l'écologie (Ye et al., 2001).
Dans ce contexte récent, la problématique biologique de cette thèse est de résoudre une partie de l'énigme de la régulation des gènes chez Buchnera. Compte tenu du rôle des bactéries dans l’association symbiotique avec les pucerons, l’étude du métabolisme global des acides aminés à l’aide de la technologie des puces à ADN semble fournir un cadre idéal pour poser cette question.
Introduction
Historique
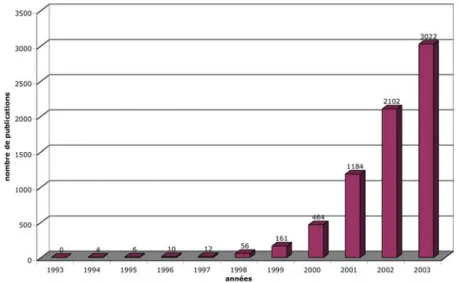
2 Contexte méthodologique Puces à ADN de A à Z (ou presque). thèse in situ), analyse d'images (acquisition de données) et informatique (interprétation de données). Depuis leur apparition, les puces à ADN suscitent un intérêt inversement proportionnel à leur taille, comme en témoigne l'explosion du nombre de publications qui leur sont consacrées depuis 2001 (voir figure A.2.1). Historiquement, les macroarrays, les microarrays et les « vraies » puces à ADN correspondent à trois méthodes d’analyse différentes (Lagoda et Regad, 2000).
Enfin, de « vraies » puces à ADN reliaient le gène de chaque organisme à un ensemble d’oligonucléotides synthétisés in situ. Aujourd’hui, ces trois distinctions ne s’appliquent plus vraiment, d’autant que ces techniques sont utilisées de manière transversale, comme le montre l’exemple des puces à ADN utilisant des produits de PCR et des cibles radioactives.
Principe des puces à ADN et analyse du transcriptome
Or, pour toutes les puces classiquement utilisées (et qui font l'objet de cette partie), l'étude du niveau d'expression des gènes repose sur la détermination des variations du niveau d'expression d'un organe. -nisme dans deux conditions différentes. Une étude expérimentale montre même que l'intensité du signal obtenu pour un fluorochrome n'est pas affectée par la présence d'une cible marquée avec l'autre fluorochrome ('T Hoen et al., 2004). Actuellement, les chercheurs tentent de concevoir une puce universelle contenant des sondes représentant toutes les combinaisons possibles de séquences d’ADN.
Il contient toutes les combinaisons possibles d'hexamères comme sondes, et son utilisation combinée à une étape de fractionnement enzymatique de la cible permettrait de générer des profils d'expression pour n'importe quel organisme (Roth et al., 2004). Les sondes synthétisées sont déposées par un robot sur la surface de la pale selon un plan défini.

Des applications variées pour les puces
Des puces à ADN
Une première expérience a été réalisée avec des ADNc (Ziauddin et Sabatini, 2001), et plus récemment une autre étude a été réalisée avec de petits ARN interférents (si-ARN) (Baghdoyan et al., 2004). D'un point de vue qualitatif, aucune différence de sensibilité n'a été observée en comparant l'utilisation de sondes oligonucléotidiques (50 mers) et de sondes PCR (Kane et al., 2000). La normalisation est une étape essentielle dans l'analyse des données de puces à ADN (Bilban et al., 2002).
Il est donc important de les tirer au sort au moment de leur utilisation (Oleksiak et al., 2002). Pour ces valeurs, le risque d'hybridation non spécifique n'est en effet pas significatif (Richmond et al., 1999). L'énergie libre est une mesure de la stabilité de l'hybridation entre deux brins, qui dépend de la température (T) (Griffin et Smith, 1998) et de la concentration en sel du milieu (Peyret et al., 1999).
Concernant les aspects quantitatifs, des études récentes suggèrent l'utilisation du modèle d'adsorption de Langmuir pour décrire l'hybridation sur puce basée sur la séquence de sondes (Hekstra et al., 2003). La présence de la première de ces bases libres (extrémité pendante) stabilise significativement l'hybride formé (Lane et al., 1992). Ces valeurs montrent que la meilleure stabilisation est obtenue avec une base A située en position 5' de la cible (Southern et al., 1999).