Chez la plante modèle Arabidopsis thaliana, leur rôle a été particulièrement démontré dans les voies de signalisation hormonale, le développement des fleurs et la réponse aux pathogènes. Le laboratoire OVGV (Organisation et Variabilité des Génomes Végétaux) de l'UMR Amélioration et Santé des Plantes, ASP travaille depuis longtemps sur la génétique moléculaire et la physiologie de la résistance du tournesol aux maladies, avec le développement d'approches génomiques, structurales et fonctionnelles.
LE BLE DANS LE MONDE
UTILISATIONS DU BLE TENDRE
CLASSIFICATION BOTANIQUE, ORIGINE ET CARACTERISTIQUES DU BLE
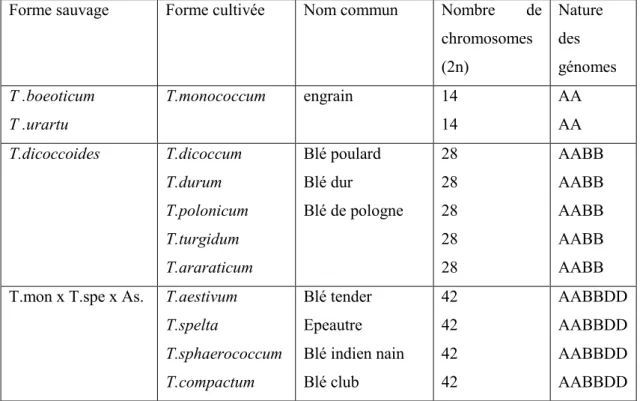
Chez le blé tendre (Triticum aestivum), la première étude d'hybrides F1 issus de croisements chromosomiques a montré que le blé tendre possède trois génomes AA, BB et DD, chacun constitué de sept paires de chromosomes homéologues, à savoir. un total de 42 chromosomes (Kihara et Nishiyama, 1930). Le blé tendre est une espèce hexaploïde, c'est-à-dire qu'il possède 6 jeux de chromosomes (2n = 42, AABBDD) ; issus d'hybridations successives entre espèces sauvages diploïdes.
AMELIORATION GENETIQUE DU BLE
La deuxième polyploïdisation s'est produite entre du blé dur tétraploïde et un autre blé diploïde, Aegilops tauschii (2n = 14, DD) pour donner Triticum aestivum, blé tendre panifiable (Feillet, 2000). Le blé est la source de protéines végétales la plus importante dans l’alimentation des humains et surtout des animaux.
GENERALITE SUR LA PROTEOLYSE
Voie calcium dépendante
Voie lysosomale
Voie ubiquitine-protéasome dépendante
LA VOIE DE PROTEOLYSE CIBLEE UIQUITINE/PROTEASOME 26S
L’ubiquitination des protéines cibles
- L’ubiquitine
- La liaison des ubiquitines aux substrats
- Enzyme activant l’ubiquitine, E1
- Enzyme conjugant l’ubiquitine, E2
- Ubiquitine protéine ligase, E3
Ensuite, l'ubiquitine est transférée de l'enzyme E1 au site actif de la cystéine de l'enzyme E2, qui la lie également via une liaison thiol-ester. Il se lie généralement au résidu lysine48 de l'ubiquitine préalablement fixé au substrat (Finley et al, 1994).
Dégradation sélective des protéines polyubiquitinées par le protéasome 26S
- Le protéasome 26S
- Reconnaissance des protéines polyubiquitinées par le protéasome 26S
- Deubiquitination et dégradation des substrats
On pense que la partie N-terminale des protéines HECT E3 est impliquée dans la reconnaissance du substrat (Ardley et Robinson, 2005). Chacune de ces 6 sous-unités est essentielle à la dégradation des protéines et peut jouer un rôle différent selon le substrat à dégrader (Rubin et al, 1998).
LE COMPLEXE SCF
Historique et Généralités sur le complexe SCF
La protéine Rbx (également appelée Roc1 ou Hrt1) est la dernière sous-unité identifiée du complexe SCF. Chez l'homme, la sous-unité Rbx a été isolée, un composant du complexe SCF et est associée à Cdc53.
Description du complexe SCF
- La sous-unité Skp1
- La sous-unité Cullin1
- La sous-unité Rbx
- La sous-unité F-box
Il a été démontré que ASK1 interagit avec la protéine CUL1 et est capable de former divers complexes SCF (Gray et al, 1999; Gagné et al, 2002). La protéine Rbx (également appelée Roc1 ou Hrt1) est la plus petite protéine du complexe SCF, composée d'environ 108 acides aminés.
Régulation du complexe SCF
- La protéolyse est dépendante de la phosphorylation
- Régulation par RUB1/NEDD8
- Régulation par CAND1
- Régulation par le complexe CSN
Deux d'entre eux, RUB1 et RUB2, sont impliqués dans la régulation de la voie de réponse auxine, de la production d'éthylène et de la croissance végétative (Bostick et al, 2004). De plus, ils ont montré le rôle de la protéine CAND1 dans la régulation positive de l'activité des complexes SCFUFO, SCFTIR1, SCFCOI1 et SCFSLY1 (Chuang et al, 2004 ; Feng et al, 2004).
IMPORTANCE DU COMPLEXE SCF CHEZ LES EUCARYOTES
Importance chez les animaux
- Régulation du cycle cellulaire
- Contrôle du cancer
La dégradation des cyclines via la voie de protéolyse dépendante de l'ubiquitine est essentielle pour l'inactivation de la CDK associée (Meijer et al, 2003; Pines, 1995). L'APC est actif de la fin de la phase G2 jusqu'au milieu de la phase G1.
Importance chez les plantes
- Régulation de la signalisation hormonale
- Voie de l’auxine
- Voie des gibbérellines
- Voie de l’acide jasmonique
- Voie de l’éthylène
- Régulations du développement floral
- Régulations en réponse à la lumière
- Régulations du Cycle circadien
- Régulations des mécanismes de défenses contre les pathogènes
Ces récepteurs sont probablement situés sur la membrane plasmique (Gilroy et Jones, 1994 ; Hooley, 1994). Comme OsGID2, le gène AtSLY1 est un régulateur positif de la voie de signalisation de l'acide gibbérellique chez Arabidopsis.
MATERIELS
- Matériel végétal et conditions de culture
- Souches de bactéries
- Souches de levure
- Plasmides
- Plasmide pGEM®-T Easy
- Plasmide pDONR™221
- Plasmide d’expression
- Acides nucléiques
- Milieux de culture
- Milieu de culture des bactéries
- Milieux de culture des levures
Le vecteur pDEST32 (12288 pb) porte le gène de résistance à la gentamicine et permet l'expression d'une protéine de fusion liée au domaine d'activation du gène GAL4. Nous avons utilisé le milieu LB pour la culture de bactéries dont la composition est la suivante.
METHODES
Etude bioinformatique
Méthodes générales de biologie moléculaire
- Extraction des acides nucléiques
- Extraction d’ADN
- Extraction d’ARN
- RT-PCR semi-quantitative
- Principe de la méthode
- Transcription inverse (RT) des ARNm en ADNc
- Amplification des ADNc
- Purification de fragments d’ADNc sur gel d’agarose
- Clonage
- Séquençage
- Analyse des séquences
Le milieu est incubé 60 minutes à 42°C (activation de la rétrotranscriptase) et 10 minutes à 75°C (dénaturation de l'enzyme). Une solution de lavage contenant du tampon Tris-EDTA (Tris-HCL 10 mM pH 8, EDTA 0,5 mM) complétée par de l'éthanol absolu permet le lavage de l'échantillon retenu par la fibre de verre.
Traitements et prélèvements
L'alignement des séquences et l'estimation du pourcentage d'identité sont réalisés à l'aide du programme Clustalx (Higgins et Sharp, 1989). Enfin, une analyse avec BLASTp (séquence protéique déduite de la séquence nucléotidique) permet de déterminer si l'on est en présence d'une séquence codante.
4 .1. Principe du clonage GATEWAY
- Préparation des fragments d’intérêt à cloner
- Dessin des amorces
- PCR recombinante
- Réaction de recombinaison BP
- Transformation criblage et purification
- Réaction de recombinaison LR
- Transformation, criblage et purification
- Etude de l’interaction protéine-protéine : technique du double-hybride
- Principe du double-hybride
- Construction des protéines de fusion « Appât » et « Proie »
- Préparation de cellules de levures compétentes
- Co-transformation des protéines de fusion dans la levure
- Criblage des transformants
- Analyse des résultats
- Détection spécifique des protéines : technique du western-blot
- Extraction de protéines totales à partir des levures
- Electrophorèse dénaturante
- Transfert sur membrane de nitrocellulose
- Détection immunologique
- RT-PCR quantitative
- Principe
- Prélèvement des échantillons
- Conditions de PCR
- Efficacité de PCR
- Quantification relative des PCR en temps réel et analyses statistiques
Il est possible de fusionner le domaine d'activation avec le domaine de liaison à l'ADN et de reproduire l'effet de la protéine entière dans des systèmes de transcription in vitro. Le système à deux hybrides d'Invitrogen tire parti des propriétés modulaires du facteur de transcription GAL4 de levure, qui consiste en un domaine de liaison à l'ADN et un domaine d'activation de la transcription. Il s’agit d’une approche simple basée sur la différence de valeur Ct entre le gène cible et notre gène d’actine de référence.
ETUDE PRELIMINAIRE DE L’EXPRESSION DE TROIS GENES SKP1, CULLIN1
RT-PCR, clonage et séquençage
- Le gène Skp1
- Le gène Cullin1
- Le géne RBX1
- Actine
L'amplification en ADNc est spécifique aux trois paires d'amorces testées ; trois bandes ont les tailles attendues (354, 437 et 568 pb), on peut donc supposer qu'il s'agit bien de séquences Cullin1. La séquence d'ADNc du gène de l'actine a été obtenue à partir de TIGR Wheat et des amorces spécifiques pour ce gène ont été générées à l'aide du logiciel Primer3. Des séquences partielles ont été clonées ; nous n'avons pas tenté d'amplifier l'intégralité de l'ADNc de l'actine.
Etude de l’expression des trois gènes sous l’effet de différentes hormones
- Profils d’expression des gènes TSK16, Cullin1 et Rbx1
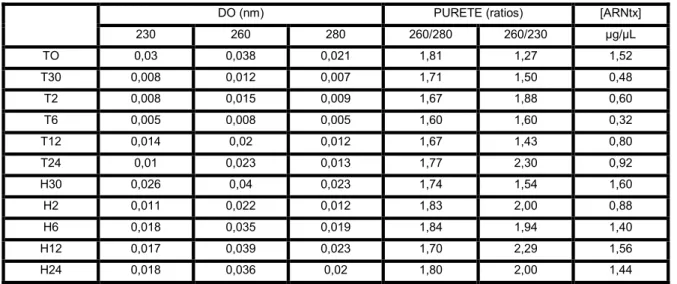
Etude de l'expression des trois gènes sous l'influence de différentes hormones. L'ARN total extrait des plantules présente une qualité satisfaisante (Fig. 22).
IDENTIFICATION DE DIFFERENTS GENES SKP1-LIKE CHEZ LE BLE T
Nomenclature des gènes SKP1-like chez le blé T. aestivum
Suite à notre analyse bioinformatique, nous avons identifié un total de vingt séquences codant pour des gènes de type SKP1. Nous avons comparé ces gènes avec d'autres gènes SKP1 identifiés dans d'autres organismes et avons découvert que 13 séquences SKP1 peuvent coder pour des séquences complètes. Nous avons nommé certains de ces gènes selon la nomenclature de Kong et al.
CLONAGE DES GENES TSKS, CULLIN1, F-BOX ET RBX1 CHEZ LE BLE
Le gène TSK16 est le gène SKP1 que nous avons identifié dans la première partie de notre travail et qui représente une délétion qui n'a pas encore été caractérisée. CLONAGE DES GÈNES TSKS, CULLIN1, F-BOX ET RBX1 DANS LE BLÉ TRITICUM AESTIVUM, À L'AIDE DE LA TECHNOLOGIE GATEWAY. Un total de 7 gènes TSK, 5 gènes F-box, un gène Cullin1 et un gène Rbx1 ont chacun été obtenus dans un clone d'entrée « pDONOR221 » et dans deux clones d'expression « pDEST22 » et « pDEST32 ».
RECHERCHE DE PARTENAIRES FONCTIONNELS POUR LES DIFFERENTES
Vérification de l’expression des différentes protéines de fusion TSKs-DBDGAL4
Malgré ce manque d'activité, les constructions TSK15 et TSK17 ont été testées dans l'espoir de découvrir des partenaires physiologiques. En effet, ces protéines de fusion peuvent être sujettes à des interactions avec d'éventuels partenaires et l'absence d'activité de liaison au motif GAL4-BD n'est pas un critère suffisant pour ne pas poursuivre cette étude.
Les protéines TSKs sont elles équivalentes ?
- La protéine TSK1
- La protéine TSK3
- La protéine TSK6
- La protéine TSK11
- La protéine TSK16
Nous avons également noté une forte interaction entre la protéine TSK1 et la protéine ATFbl5. Cependant, nous avons noté (Fig. 34) que la protéine TSK11 n'interagit que dans le sens dans lequel cette protéine se déplace vers la position de la proie. Ainsi, dans cette étude, nous n'avons identifié aucun partenaire cloné de la protéine TSK16 (Fig. 35).
ETUDE STRUCTURE/FONCTION DE LA PROTEINE TSK11
L’interaction et/ou l’autoactivation entre TSK11 délétée de son domaine en N-
Nous avons effectué une délétion respectant le cadre de lecture de la protéine TSK11 pour obtenir une séquence TSK11 tronquée à laquelle nous avons ajouté un codon d'initiation de traduction ATG (Fig. 37). Nous avons étudié l'interaction entre 2 protéines de fusion en surveillant l'expression du gène rapporteur LacZ de la souche hôte, et deux expériences indépendantes ont été réalisées. Parallèlement, nous avons vérifié par Western blot que la protéine TSK11 tronquée est exprimée dans la levure et que le retrait d'un fragment ne modifie en rien l'expression du gène, tant que le cadre de lecture est respecté.
L’interaction entre la protéine chimère N-domain-TSK6 et les protéines ZTL et
L'interaction entre la protéine chimérique TSK6 du domaine N et les protéines ZTL et ATFBL5 a été établie.
TSK11 forme-il un homodimère ?
Cependant, la protéine tronquée a perdu sa flexibilité et devient incapable de former un homodimère (Fig. 40b).
EXPRESSION DES GENES TSKS CHEZ LE BLE
Le but de ce projet de recherche était de vérifier l'interaction entre la sous-unité Skp1 et ses différents partenaires du complexe SCF. La sous-unité Skp1 agit comme un adaptateur entre l'assemblage Cullin1-Rbx1, la sous-unité catalytique du complexe SCF et la protéine F-box responsable de la reconnaissance spécifique du substrat (Bai et al, 1996; Skowyra et al, 1997). La caractérisation moléculaire et biochimique de la sous-unité SKP1 a donc été entreprise chez le blé hexaploïde.
LA FAMILLE SKP1 CHEZ LE BLE TRITICUM AESTIVUM
L'étude de la fonction d'une protéine implique souvent la connaissance de ses partenaires d'interaction afin d'obtenir des informations sur la ou les fonctions de cette protéine. Diverses études ont montré que l'expression du gène SKP1 varie selon les organes et les différents stades de développement. Cependant, pour notre étude, nous avons isolé les gènes TSK de plants de blé âgés de 2 à 3 semaines et, malheureusement, nous n'avons pas pu amplifier tous les gènes SKP1 du blé.
RECHERCHE DE PARTENAIRES FONCTIONNELS
En revanche, il est probable que les protéines TSK1 et TSK3 forment un complexe NON-SCF. Notre étude a révélé que les protéines TSK6 et TSK16 ne se lient ni aux protéines F-box ni à la protéine Cullin1. Au cours de notre étude, nous avons comparé les séquences d'acides aminés des 7 gènes TSK du blé avec la protéine SKP1 humaine et d'autres protéines Skp1 appartenant à différentes espèces (Tableau 7).
Etude structure-fonction de la protéine TSK11
Pour vérifier ce résultat, nous avons fusionné ces 29 acides aminés du N-Ter à la protéine TSK6, qui n'a interagi avec aucune des protéines du complexe SCF étudié. Il est également probable qu’en plus des 26 acides aminés clés, la structure 3D de la protéine entière contribue de manière significative à l’établissement de ces interactions. Cependant, au cours de ces travaux, nous avons montré qu'en plus de ces deux domaines conservés, il existe des domaines supplémentaires, soit 29 acides aminés pour la protéine TSK11, qui sont directement impliqués dans l'interaction des protéines SKP1 avec les protéines F-box.
L’expression relative des gènes du complexe SCF chez le blé
Members of the Arabidopsis SKP1-like gene family exhibit a variety of expression patterns and may play diverse roles in Arabidopsis. The F-box subunit of the SCF E3 complex is encoded by a diverse superfamily of genes in Arabidopsis. Subcomplex of the proteasome regulatory particle required for ubiquitin conjugate degradation and related to the COP9 signalosome and eIF3.
Loss of circadian clock-related protein 1 in Arabidopsis results in altered clock-regulated gene expression. A large complement of predicted Arabidopsis ARM repeat proteins are members of the U-Box E3 family of ubiquitin ligases. Members of the ASK gene family show different expression patterns and may play different roles in Arabidopsis.
Ubiquitin is a 76-residue protein that can be covalently attached to target proteins by an enzymatic conjugation cascade involving three enzymes indicated, E1, E2 and E3. The SCF complex is a type of ubiquitin protein ligase (E3) which as the specific factor responsible for substrate recognition and ubiquitination.