INTRODUCTION SUR LES VECTEURS DE GENES
L A THERAPIE GENIQUE
Principe et Résultats
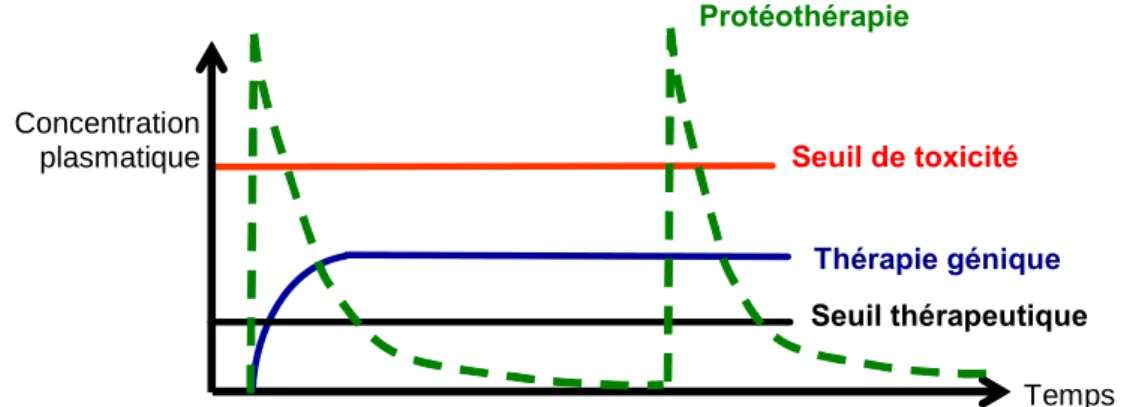
La thérapie génique est née du premier essai clinique sur l'homme, réalisé en 1989 par l'équipe de Rosenberg7 pour traiter des patients atteints d'un mélanome avancé. La plupart de ces essais ont montré la faisabilité et le fort potentiel de la thérapie génique, même si son efficacité n’est pas encore optimale.
Les vecteurs viraux
Soyez spécifique à un tissu ou à un type de cellule, surtout si ces cellules sont réparties dans tout le corps. Les différents types de virus utilisés en thérapie génique se multiplient.

Les vecteurs non viraux
Bien que cette méthode soit prometteuse pour transfecter in vitro des cellules difficilement accessibles avec les techniques classiques, comme les neurones ou celles du cerveau, son principal inconvénient est que sa pénétration dans les tissus in vivo n'est que faible. À l’aide d’un laser femtoseconde proche infrarouge, l’équipe de Zeira a obtenu une expression à long terme (> 100 jours) du gène de l’érythropoïétine sans dommage tissulaire ni effets secondaires51.
L ES VECTEURS CATIONIQUES
Lipides cationiques
1996) Guanidinium cationic cholesterol lipids: efficient vectors for transfection of eukaryotic cells, Proc. 1997) Structure of DNA-cationic liposome complexes: DNA intercalation in multilamellar membranes in different regimes of interhelical packing, Science. Nomenclature for synthetic gene delivery systems, Hum. 1997) Structure of DNA-cationic liposomal complexes: DNA intercalation in multilamellar membranes in distinct interhelical packing regimes, Science.

Les polymères cationiques
L'injection du gène de l'interleukine IL-2 associé à un vecteur de type DMRIE a donc été réalisée chez des patients atteints de différents cancers. Le traitement a été bien toléré et deux patients ont présenté une réponse antitumorale partielle pendant 16 à 19 mois.70 Les recherches actuelles se concentrent désormais sur l'amélioration de l'efficacité encore limitée de ces systèmes.
Barrières au transfert de gènes
Les vecteurs captés par ce mécanisme doivent s'échapper de l'endosome avant d'atteindre le lysosome. Cela conduit à une rupture osmotique de la vésicule, une rupture de l'endosome et une libération des complexes dans le cytosol.

L ES V ECTEURS NON CATIONIQUES
Vecteurs anioniques
Les vecteurs neutres existants
Roux a montré qu'il était possible d'insérer des oligonucléotides138 ou de l'ADN plasmidique139 et ainsi de les utiliser comme vecteurs de gènes in vivo, car ils sont non toxiques et ne s'associent pas aux protéines plasmatiques. Pour les améliorer, il faudra trouver une manière alternative d’interagir avec l’ADN en l’absence de charges positives.

Vers un nouveau type d’interaction avec l’ADN
Ces résultats montrent d'une part l'effet de la quantité de lipides sur l'accessibilité de l'ADN. Il est particulièrement remarquable que l’allongement de la chaîne améliore la rétention de l’ADN dans le gel et la transfection.
L ES LIPOPOLYTHIOUREES
La fonction thiourée
Les liaisons hydrogène intermoléculaires sont plus faibles que dans le cas de l’urée, car le soufre est un moins bon accepteur de liaisons hydrogène que l’oxygène153. Les liaisons hydrogène formées sont plus flexibles en raison de la polarisation plus forte du soufre par rapport à l’oxygène154.
Résultats du lipide DT3TU
L'incubation de ces liposomes DT3TU/DPPC 1/2 avec l'ADN donne naissance à des agrégats de 800 nm (Figure 21), démontrant que l'ADN est associé au liposome. Il est possible de retrouver une zone de stabilité colloïdale supérieure à celle des lipides (>15 nmol DT3TU/μg ADN).
Conclusion
Injecté par voie intraveineuse à des souris porteuses de tumeur, leur localisation a été étudiée 30 minutes après l'injection (Figure 23). Enfin, nous observons une accumulation passive dans les tumeurs 7 fois supérieure à RPR209120 à 30 minutes, ce qui semble particulièrement intéressant du point de vue du ciblage tumoral.
OBJECTIF DU TRAVAIL
Cependant, nous avons choisi de modifier légèrement la structure de la tête du lipide DT3TU. Nous avons étudié la formation de lipoplexes en mesurant la taille des particules (Figure 39) et par un test de retard de migration électrophorétique (Figure 40).

D EFINITION DU PROJET
R EALISATION
Les éléments fixes
L'élément essentiel qui doit être conservé dans toutes les structures lipopolythiourées est la présence d'unités thiourée dans la tête polaire, car celles-ci sont responsables de l'interaction avec l'ADN. Il était donc important d’évaluer le nombre minimum de fonctions thiourée nécessaires au maintien de l’interaction avec l’ADN.
Structure de la famille de lipopolythiourées
La tête polaire a été choisie ramifiée plutôt que linéaire, car cette modification permet d'améliorer le transfert de gènes dans le cas des lipopolyamines cationiques170. Une structure bi-antenne permet également l'introduction de deux extrémités au lieu d'une. , qui peut être varié. . Il est clair que l'influence du nombre de motifs thiourée n'est pas clairement définie, puisqu'une bonne corrélation avec l'ADN est observée avec 3 ou 4 motifs.
Les éléments variables
Le diamètre des particules de 11/CholPEG2000 fluctue en présence d'ADN et cette fluctuation dépend de la quantité de lipopolythiourée. Au niveau de la formulation, nous avons pu constater une grande amélioration de l’hydrophilie des composés par rapport au DT3TU.
SYNTHESE D’UNE FAMILLE DE LIPOPOLYTHIOUREES
G ENERALITES
Stratégie
Ensuite, nous avons modifié les extrémités de la tête polaire en ajoutant soit une ou deux unités hydroxyle, soit un groupe imidazole. Enfin, nous avons modifié l'ancre hydrophobe en réduisant la longueur de la chaîne ou en la remplaçant par un cholestérol.
Réactions « clé »
En bref, ces lipides sont capables de se lier à l'ADN de la même manière que le lipide DT3TU, confirmant qu'une tête ramifiée comportant deux unités thiourée permet une interaction avec l'ADN. Plus précisément, nous avons choisi de développer des vecteurs basés sur le modèle de lipopolitiurée DT3TU.
P REPARATION DES PREMIERES LIPOPOLYTHIOUREES
Rétrosynthèse
La fonction thiourée est finalement introduite par la réaction de l'amine avec le lipide isothiocyanate issu de l'amine.
Synthèse des précurseurs
Nous avons ensuite préparé l'acétal 4, qui a été obtenu en une étape par la réaction de l'acide 4-formylbenzoïque avec le chlorure de thionyle dans le méthanol.195 L'acétalisation de l'aldéhyde a été favorisée par la libération d'acide chlorhydrique résultant de la formation de l'ester méthylique (Schéma 7). Le traitement de 5 avec de l'azoture de sodium dans du DMF à 100°C197 a donné le di-azide 6, caractérisé par un signal fort provenant du carbone α de l'azoture à 51 ppm.
Assemblage du vecteur
Dans tous les cas, l’intensité de fluorescence représente 90% de la fluorescence Picogreen en présence d’ADN libre.
V ARIATION DE L ’ ESPACEUR ET DU REPARTITEUR
Espaceurs succinique et diglycolique
La préparation du composé 17 a commencé par la protection de la sérinol amine par le groupe tert-butoxycarbonyle (Boc). L'examen des spectres RMN confirme la réaction avec le déplacement à 4,15 ppm du proton vers α de l'azote espaceur.
Espaceur PEG
Le composé 30 peut subir une transacétalation avec le diol 3 conduisant à un mélange d'acétal 31 encore protégé par le THP (34 %) et d'acétal 32 comprenant un alcool libre (23 %). Nous avons choisi de former l'isothiocyanate sur le lipide 36 car la déprotection de l'isopropylidène, dans la méthode alternative proposée pour 28, entraînerait une hydrolyse du fragment acétal.
Répartiteur dissymétrique
Si ce composé s'avère intéressant lors de l'évaluation biologique, il faudra trouver une autre voie de synthèse plus rapide et plus efficace, par exemple à partir de PEG asymétriques commerciaux. Il conduit à l'amide 40 avec un rendement de 67%, identifié par un déplacement du signal protonique vers l'azote α.

V ARIATION DES TERMINAISONS
- Terminaison alcool
- Terminaison diol
- Terminaison méthyle
- Terminaison imidazole
Nous avons émis l'hypothèse que la thiourée dans un environnement acide se réorganise pour former un cycle à 5 chaînons. Enfin, nous avons envisagé de diversifier davantage la structure en introduisant des unités imidazole aux extrémités de la tête polaire.
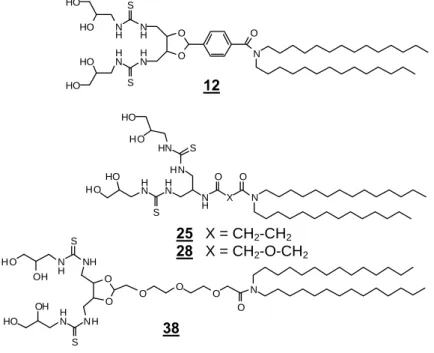
V ARIATION DE L ’ ANCRE HYDROPHOBE
Longueur des chaînes aliphatiques
Le spectre RMN montre le pic de thiourée carbonyle à 184 ppm, ainsi qu'un singulet à 3,01 ppm de proton, correspondant aux groupes méthyles terminaux. Le couplage de la lysine protégée 39 sur la didodécylamine ou la didécylamine est réalisé dans les mêmes conditions que 40.
Ancre cholestérol
Efficacité de la transfection après injection intratumorale de complexes lipopolitiourée/ADN à l'aide d'un gène rapporteur luciférase. La quantité de lipides nécessaire à cette interaction varie en fonction de la structure des composés.
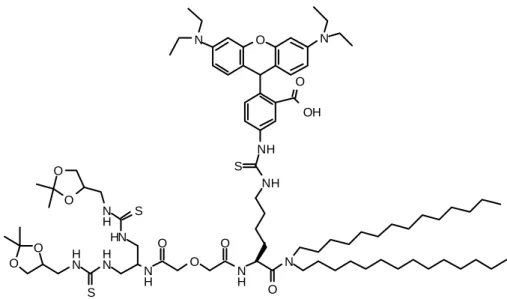
S YNTHESE D ’ UNE THIOUREE FLUORESCENTE
Synthèse
Le couplage de l'isothiocyanate de rhodamine B avec l'amine 73 est ensuite réalisé en présence de triéthylamine dans le dichlorométhane, dans des conditions analogues à la formation de la thiourée 74. La structure de la thiourée 76 a été établie par examen du spectre RMN, qui montre la présence de l'aromatique. protons du chromophore, et par analyse LC-MS, qui donne le pic de masse 786 ((M+2H)/2)+.
C ONCLUSION
Bilan des vecteurs synthétisés
Dans l'étape suivante, l'aminé latérale est déprotégée par reflux du méthanol en présence de carbonate de potassium221 (Schéma 32).
Conclusion
Les résultats de toxicité des lipopolyurées après injection intraveineuse sont présentés dans la figure 61. Des problèmes de reproductibilité importants sont survenus lors de la mesure du potentiel zêta des lipopolyurées.
ETUDE DES PROPRIETES PHYSICO-CHIMIQUES ET BIOLOGIQUES DES
S TRATEGIE ET METHODES
Méthodes physico-chimiques
Enfin, la formation de lipoplexes se produit spontanément par ajout d'une solution aqueuse d'ADN plasmidique à la formulation lipidique obtenue précédemment. Utiliser une sonde telle que Picogreen238 qui émet une fluorescence lorsqu'elle est insérée entre des bases d'ADN.

Critères d’évaluation biologique
B : Fluorescence de la Rhodamine et de la GFP par rapport au segment défini en A C : Internalisation des complexes 28. L'ajout de dextrane au milieu de culture n'a pas d'effet sur l'activité de transfection de la lipopolyurée 58 ou de la lipopolyamine RPR 209120.
E VALUATION DES FORMULATIONS DES LIPOPOLYTHIOUREES
Validité du modèle
Ces deux lipides ont ainsi démontré leur capacité à associer suffisamment fortement l'ADN pour le retenir sur gel. Les liposomes alors formés sont capables d'induire une interaction avec l'ADN, modifiant le diamètre des particules et retenant l'ADN sur le gel.
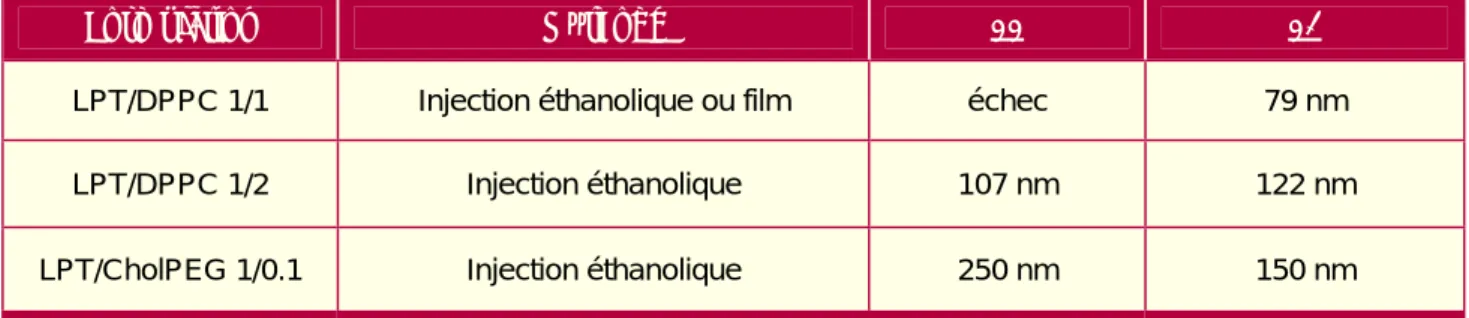
Evaluation systématique des lipopolythiourées
Cela indique que l'ADN reste accessible à l'intercalateur même lorsqu'il est associé à des composés lipopolythiourées car il est retardé sur le gel. Pour toutes les formulations testées, nous avons observé une rétention totale de l'ADN sur gel, mais à un rapport LPT/P variable.

Automatisation de la formulation
Dans cet exemple, l'ADN est ralenti à partir de 2,5 LPT/P, mais sa rétention n'est complète qu'à partir de 10 LPT/P. Pour toutes les formulations testées, nous avons observé une rétention totale de l'ADN sur gel, mais à un rapport LPT/P variable. Pour ce faire, nous avons créé un programme qui suit les étapes d'une injection d'éthanol (Tableau 11), et nous avons comparé les tailles des liposomes à des injections réalisées « à la main ».
E TUDES BIOPHYSIQUES
- Accessibilité de l’ADN
- Morphologie des complexes lipopolythiourée/ADN
- Conformation de l’ADN
- Protection de l’ADN par les complexes
Nous avons donc réalisé des mesures de dichroïsme circulaire de l'ADN plasmidique dans l'eau, puis en présence d'une quantité croissante de lipopolitiourée 59 (Figure 47). Nous avons montré que les lipopolyurées sont capables de former de petits complexes homogènes avec l'ADN.

E VALUATION IN VITRO
Efficacité de transfection
Il apparaît clairement qu'une incubation de 48 heures permet d'améliorer l'activité de transfection de 59 de 30% en moyenne. Nous avons également étudié la capacité de transfection de la lipopolythiourée sur d'autres types cellulaires, notamment les cellules Eahy humaines.

Etudes de ciblage
Nous avons ensuite considéré les lipopolythiourées biologiquement actives et étudié l'influence de l'ajout d'un élément de ciblage sur leur activité de transfection. Cette efficacité semble augmenter avec la quantité d'élément cible (de 1% à 5%), mais la cible semble moins active lorsque la quantité de lipide est importante (surtout à 40 LPT/P).

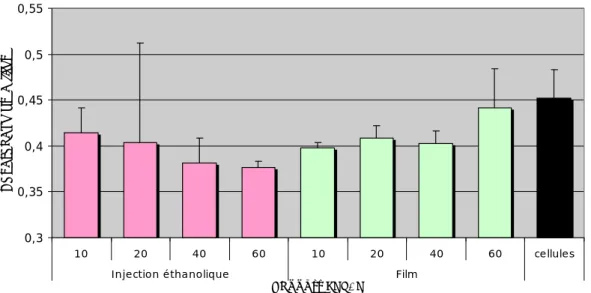
Toxicité in vitro
La formulation par hydratation d'un film lipidique permet de maintenir une quantité constante de protéines et, par conséquent, de réduire la toxicité des complexes. La toxicité des complexes peut également être modifiée par la composition lipidique de la formulation.
E VALUATION IN VIVO
Toxicité in vivo
Efficacité de transfection intratumorale
Biodistribution
Tout d'abord, on constate que deux heures après l'injection, entre 8 et 11 % de la dose injectée restent dans le sang pour respectivement 42/DPPC et 57/69, ce qui est comparable à un liposome neutre. Enfin, nous constatons malheureusement une certaine accumulation dans les tumeurs, mais nous n'avons introduit aucun élément de ciblage pouvant conduire à un tropisme particulier.

C ONCLUSION
DISCUSSION
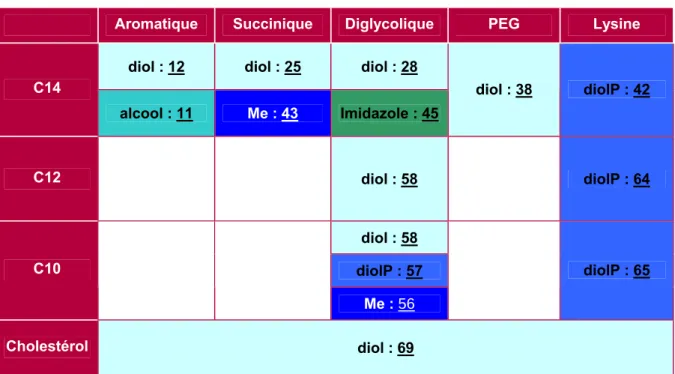
Etudions d'abord le cas des lipopolythiourées 42, 64 et 65 de la famille des lysines (Tableau 14). Il semble clair que ce mécanisme dépend largement de la charge des complexes ADN/vecteur.
L ES LIPOPOLYTHIOUREES
Stabilité des lipopolythiourées
Des difficultés ont été rencontrées lors de cette hydrolyse et elle doit être suivie par LC-MS pour éviter la dégradation de la thiourée. Il est possible que, malgré la congélation puis la lyophilisation du milieu réactionnel, de l'acide résiduel provoque une dégradation de la lipopolitiurée en début de stockage, l'analyse ELSD après lyophilisation étant appropriée.
Etude de la charge des complexes
Pour mieux comprendre le recrutement des ions par la lipopolyurée, nous avons étudié le potentiel zêta des particules en fonction du pH de la solution (Figure 65). Une fois la synthèse de chaque lipopolythiourée terminée, nous avons effectué une chromatographie sur couche mince détectée par la ninhydrine.

Toxicité
Par ailleurs, il est reconnu que l'efficacité des systèmes cationiques est généralement fortement réduite par la présence de sérum89. C'est pourquoi nous avons souhaité étudier l'influence du sérum sur la capacité de transfection des lipopolythiourées (Figure 68). Si la présence de sérum affecte également l'activité des lipopolythiourées 57 et 58, son influence est beaucoup moins importante, puisque l'activité est divisée par 4 au maximum.
Comparaison des lipopolythiourées avec le lipide DT3TU et les systèmes cationiques
On peut également étudier les lipopolythiourées de la famille des diglycols dont les résultats sont résumés dans le tableau 15. Ainsi, les terminaisons et l'espaceur affectent principalement la formulation des composés, et la longueur de chaîne semble plus déterminante pour l'activité de transfection.
E TUDES STRUCTURE / PROPRIETES
Influence de l’espaceur et du répartiteur
La variation espaceur ne permet pas de conférer une activité biologique à un lipide possédant des chaînes aliphatiques de 14 atomes de carbone à extrémités diol. Enfin, on constate que la nature de l'espaceur semble influencer principalement la formulation et ne semble pas être un paramètre déterminant pour la capacité de transfection.
Influence de la terminaison
De plus, les composés de la famille des lysines ne possèdent pas d'espaceur mais un diviseur asymétrique. En bref, on peut conclure que la nature de la terminaison n'a pas d'influence déterminante sur les propriétés biologiques, mais permet de moduler les propriétés physico-chimiques, puisque dans tous les cas la présence du diol semble faciliter la formulation de composés lipopolythiourées.

Influence de la chaîne : famille lysine
Influence de la chaîne : famille diglycolique
On obtient 65 mg d'alcool 32 libre, ce qui correspond à un rendement total de la réaction de 52 %. Après évaporation, le produit brut est purifié sur cartouche de silice pour obtenir 46 mg d'huile (63 %).
E TUDE MECANISTIQUE
Voie d’entrée dans la cellule
Premièrement, nous voulions examiner l’effet de l’ajout d’un excès de lipides sur la transfection des complexes 58/ADN. Pour ce faire, nous avons effectué une transfection de complexes 58/ADN à 10 ou 13 LPT/P auxquels un nombre spécifié d'équivalents lipides 58 ont été ajoutés.

Sortie de l’endosome
En revanche, l’ajout de sulfate de dextrane inhibe complètement la transfection du RPR 209120, démontrant que les lipoplexes doivent interagir avec les GAG pour être internalisés. On peut émettre l'hypothèse que les lipopolythiourées 57 et 59, de par leur chaîne à 10 carbones, permettraient un meilleur échange avec les lipides des membranes endosomales, ce qui améliorerait la délivrance d'ADN, et donc l'efficacité de la transfection.

C ONCLUSION
CONCLUSION GENERALE ET PERSPECTIVES
PARTIE EXPERIMENTALE
High efficiency gene transfer to skeletal muscle mediated by electrical pulses, Proc. 2004) DNA electrotransfer: its principles and an updated review of its therapeutic applications, Gene Ther., 11, S33-S42. Direct gene transfer by DNA-liposome complexes in melanoma: expression, biological activity and lack of toxicity in humans, Proc. 2005) Cellular delivery mediated by Tat peptides: back to basics, Adv.