Contrôle avec gel d'agarose de la fixation de l'ADN endommagé sur les billes magnétiques. 231 Chromatogrammes et spectres typiques obtenus lors de la quantification des lésions de l'ADN par HPLC-. Enfin, nous discuterons de l'utilisation des deux microsystèmes susmentionnés ainsi que d'autres approches disponibles dans notre laboratoire pour mener une étude pilote axée sur le rôle de la protéine DNA Damage-binding Protein 2 (DDB2) dans le traitement des lésions déformant l'ADN ( photoproduits et adduits de platine).
L ES AGENTS ANTICANCÉREUX DÉRIVÉS DU PLATINE
Généralités sur les principaux dérivés du platine
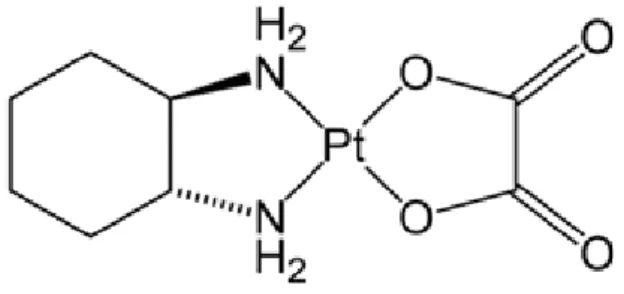
Le cisplatine, complexe coordonné du platine historique
Il est également fréquemment utilisé (avec moins de succès) contre le cancer des bronches à petites cellules, ORL, des ovaires et de la vessie (Kartalou & Essigmann, 2001b ; Rabik & Dolan, 2007). Pour le cancer ORL, le cisplatine est utilisé en association avec le 5-fluorouracile (5-FU, antimétabolite analogue de l'uracile qui interfère avec la synthèse de l'ADN et de l'ARN) dans le cadre de traitements néoadjuvants visant à préserver la fonction des organes affectés comme le larynx, le pharynx et langue.

Autres exemples de dérivés du cisplatine
Cependant, leur faible solubilité dans l’eau a conduit à des améliorations sur ce point précis, conduisant à l’oxaliplatine (Stordal et al., 2007). Pour les mêmes raisons, la molécule réagit peu avec les protéines plasmatiques, ce qui contribue à augmenter sa biodisponibilité (Bell et al., 2008).
Stratégies originales faisant appel à des dérivés du platine
L'activité de plusieurs isozymes de la GST est efficacement inhibée après exposition des cellules tumorales pulmonaires à l'étacraplatine, confirmant l'intérêt d'une telle molécule. Après stockage dans la particule (Figure 9), le cisplatine peut être spécifiquement délivré dans les cellules exprimant le récepteur des facteurs de croissance épidermiques humains 2 (HER2) reconnu par l'anticorps, et initier une cytotoxicité supérieure à celle du cisplatine (Cheng et al., 2009). .

Données pharmacologiques sur les dérivés du platine
- Données pharmacocinétiques
- Pénétration à l’intérieur de la cellule
- Activation intra-cytoplasmique
- Formation d’adduits sur l’ADN génomique
- Proportions des différentes lésions de l’ADN
- Répercussions des lésions sur la structure de l’ADN
- Cytotoxicité des différents types de lésions
Sept ans après la mise sur le marché de la molécule, différentes formes d'adduits du cisplatine sur l'ADN ont été identifiées (Fichtinger-Schepman et al., 1985). Des données déjà anciennes parlent d'un plus grand pouvoir mutagène de la lésion 1,2-d(ApG) par rapport à la lésion 1,2-d(GpG) (Burnouf et al., 1990).

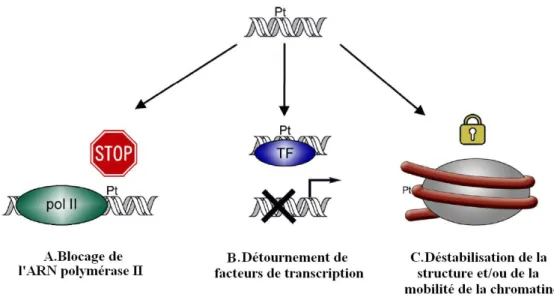
Conséquences biologiques de la présence d’adduits du platine sur l’ADN
Conséquences directes
Enfin, l'inhibition de la transcription peut entraîner une augmentation de la rigidité des nucléosomes suite à la formation d'adduits du platine sur l'ADN. La présence de lésions sur l'ADN inhibe l'activité de la protéine CDK, entraînant
Réparation des lésions
La présence d'un mésappariement en face de l'adduit (introduit par synthèse translésionnelle) amplifie la déformation de la double hélice et augmente l'efficacité de la réparation par le système NER (Moggs et al., 1997). L’un des mécanismes modulateurs de la réparation des adduits de platine est l’effet protecteur des lésions (repair blinding) par certains facteurs, notamment les protéines HMG box (high Mobility Box).

Induction de l’apoptose
Les modifications post-traductionnelles des histones (épigénétiques) affectent cette accessibilité et donc aussi le fonctionnement de la voie NER (Jung & Lippard, 2007). Nous verrons que les voies menant à l’induction de l’apoptose n’impliquent pas toujours des dommages à l’ADN, mais aussi une modification directe des mitochondries par des dérivés du platine.

Bases moléculaires de la résistance aux dérivés du platine
Mécanismes empêchant les dérivés du platine d’atteindre l’ADN
Le mécanisme exact d’entrée du cisplatine dans la cellule n’a pas encore été clairement élucidé, comme nous l’avons vu lors de la description de cette étape. L’existence d’autres rôles du glutathion dans la médiation de la toxicité du platine doit être envisagée.
Mécanismes intervenant après la formation des lésions
La contribution de la voie NER à la résistance au cisplatine est modeste (Siddik, 2003), mais peut néanmoins être suffisante pour provoquer l'échec du traitement chimiothérapeutique (Aebi et al., 1996). L'acquisition de capacités de résistance par les cellules tumorales ovariennes et colorectales pourrait être attribuée à la perte de MMR, et notamment de MLH1 (Aebi et al., 1996).

I NTERACTION DES ADDUITS PLATINÉS AVEC DES PARTENAIRES PROTÉIQUES
Interaction avec les protéines de réparation
- Interactions avec les protéines de la voie NER
- Interactions avec les protéines de la voie MMR
- Interaction avec les protéines de réparation des cassures simple-brin
- Interactions avec les protéines de la réparation des cassures double-brin
Les bases flanquantes de la lésion semblent n'avoir aucune influence sur la capacité de reconnaissance de DDB2 (Scrima et al., 2008). De plus, l'expression de DDB2 est maximisée pendant la transition G1/S, ce qui est cohérent avec un mécanisme de contrôle de l'ADN génomique avant sa réplication (Nag et al., 2001).
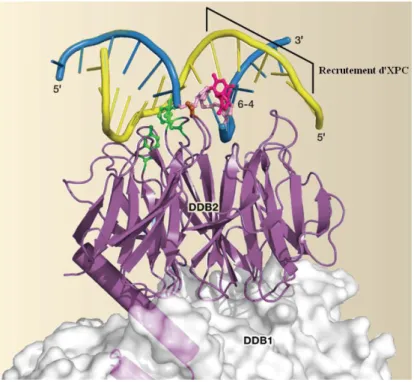
Autres acteurs de la reconnaissance des lésions platinées
- Le facteur de transcription p53
- Protéines à boîtes HMG
- Histone H1
- TBP
- YB-1
- M ÉTHODES D ’ ÉTUDE DES INTERACTIONS ENTRE ADN LÉSÉ ET PROTÉINES
XPC-HR23B Cisplatine, carboplatine, oxaliplatine, satraplatine Reconnaissance des lésions déformant l'ADN (Sugasawa et al., 1998). RPA Cisplatine Protection du fragment simple brin pendant la NER (Clugston et al., 1992).

Caractérisation d’une interaction simple : une protéine/une lésion
Retard sur gel (EMSA)
La confirmation de l'interaction peut être obtenue, par exemple, en ajoutant un concurrent spécifique qui annulera l'interaction. Enfin, l'introduction d'un concurrent spécifique non radiomarqué (piste 4) permet de supprimer partiellement l'interaction (barre rouge clair).

Autres techniques faisant appel à de l’ADN en phase homogène
Cela a par exemple permis de confirmer le mode de liaison du domaine HMG A de la protéine HMGB1 à la majorité des adduits cisplatine (Ohndorf et al., 1999). A titre d'illustration, la liaison de la protéine bactérienne MutS (analogue bactérien de hMutα) à divers dommages à l'ADN a été étudiée à l'aide d'une technique de fluorescence induite par transfert d'énergie (Lopez-Crapez et al., 2008).

Techniques faisant appel à de l’ADN immobilisé (analyses en phase hétérogène)
C'est le minimum de la courbe de réflectivité représentée en fonction de l'angle d'incidence du rayonnement (figure 24). La première est la visualisation en temps réel sur un écran de l’ensemble de la surface de travail.

Identification d’un interactome
- Ligand fishing : principe et exemples
- Ligand fishing : améliorations récentes
- Puces à protéines
- M ATÉRIEL & M ÉTHODES
Cependant, une certaine surreprésentation des protéines impliquées dans la réplication et la réparation de l'ADN (AGT, XRCC1, PARP-1, Ku70/80, Ref-1) a été notée, démontrant l'efficacité du piège. Seules quelques études de ce type ont encore été réalisées en ce qui concerne les dommages à l'ADN.

Construction et utilisation du système de ligand fishing
Principe général
Contrairement aux systèmes que nous avons décrits lors de l'analyse bibliographique, notre piège est composé d'ADN plasmidique. La partie responsable de l'activité exonucléase 5' → 3' est éliminée par traitement avec la sérine protéase subtilisine, ce qui donne une polymérase fonctionnelle qui est le fragment de Klenow.

Protocoles détaillés
Les premiers tests de platination de l'ADN impliquaient l'exposition des plasmides après fixation sur billes magnétiques à une concentration de 33 µM de cisplatine (poudre lyophilisée diluée à 15 mg/ml dans du DMSO, Sigma) pendant 2 h à 37 °C à l'abri de la lumière et sous agitation vigoureuse ( 1 100 tr/min). Un contrôle de la fixation efficace de l'ADN est réalisé en déposant les différentes solutions d'incubation et de rinçage sur un gel d'agarose.
Analyse des protéines
Coloration au nitrate d’argent
La préparation des échantillons destinés aux analyses protéomiques a suivi un protocole similaire, avec les variantes suivantes : l'échantillon représentatif de chaque condition est constitué de la combinaison de trois échantillons identiques préparés simultanément selon la méthode décrite ci-dessus pour augmenter la quantité de protéine ; l'élution par tampon dénaturant est réalisée à l'aide d'un tampon de type Laemmli (concentrations finales : Tris-HCl 25 mM pH 6,8 2% dodécylsulfate de sodium 4% glycérol 5% β-mercaptoéthanol 0,0008% bleu de bromophénol) préparé au Laboratoire d'Etude du Protéome Dynamique (LEDyP); L'élution par une nucléase suit le même protocole que décrit précédemment, les protéines étant finalement dénaturées à l'aide du tampon de type Laemmli préparé avec LEDyP.
Immunodétection (Western blotting)
La membrane est ensuite bloquée dans une solution de lait lyophilisé 0,5% TBS 0,5% Tween-20 5% pendant 1 heure et l'incubation avec l'anticorps primaire dilué dans la même solution est réalisée pendant une nuit à 4°C sous agitation douce. Tween-20, puis incubation avec l'anticorps secondaire couplé à la peroxydase de raifort est réalisée dans la même solution pendant 1 heure.

Protéomique
Protocole d'analyse des échantillons de protéines – Les échantillons sont concentrés par SDS-PAGE en bandelette de 2 mm (stack migration) sur gels NuPAGE préfabriqués (Invitrogen) par migration (durée < 2 minutes) à 200 V dans un MES 50 mM 50 mM Tris 0 tampon, 1 % SDS 1 mM EDTA. Les protéines sont ensuite fixées par un bain de 30 minutes dans un tampon acide (acide acétique 7,5%, EtOH 30%) et colorées par un bain de 30 minutes dans une solution de bleu de Coomassie (Bio-Rad).

Quantification des lésions de l’ADN
Protocole détaillé
Il s’agit généralement d’un des pics minoritaires indiquant une efficacité de réaction inférieure à celle du produit d’addition 1,2-d(GpG). Les transitions recherchées (description de la masse et des fragments) lors de l’analyse par spectrométrie de masse sont répertoriées dans le tableau 9.

R ÉSULTATS
Optimisations du système de ligand fishing
- Préparation des sondes plasmidiques
- Tests de capture de protéines choisies
- Évaluation de deux méthodes de relargage des protéines piégées
- Test préliminaire utilisant le mode de relargage total des protéines
Nous avons conclu que cela n’est pas dû à l’action directe du platine sur les billes, mais plutôt à l’ADN lui-même endommagé. Pour cela, deux endonucléases ont été évaluées sur le piège nu, c'est-à-dire l'ADN endommagé fixé sur les billes magnétiques (sans protéine capturée) : la nucléase P1 et la DNase I.

Détermination de l’interactome des lésions platinées
D ISCUSSION
Malgré les efforts déployés depuis une quarantaine d’années, de nombreuses zones d’ombre subsistent quant aux conséquences biologiques des dommages à l’ADN provoqués par les dérivés du platine. Depuis un peu plus de 10 ans, plusieurs approches de pêche aux ligands ont été mises en œuvre dans le but d'identifier des protéines qui interagissent avec des lésions de l'ADN, chacune reproduisant des résultats antérieurs et/ou donnant naissance à de nouveaux candidats.
Considérations sur le protocole de piégeage des protéines
- Structure de la sonde et nature des lésions
- Exhaustivité de la liste de protéines identifiées
- Limitation des protéines non pertinentes
- Aspect quantitatif
- Bilan
Cela peut aussi (dans une moindre mesure) être une conséquence de l’utilisation d’une sonde plasmidique. La sonde photoactivable n'est donc pas totalement représentative de l'ADN cellulaire endommagé par le cisplatine.
L’interactome des lésions platinées : protéines connues et candidats originaux
Membres de l’interactome déjà connus
L'absence d'ADN ligase III sur nos sondes cisplatine semble cohérente avec cette observation, qui montrerait le problème de représentativité de ces sondes photoactivables. Pour prendre une décision définitive, il serait donc intéressant de confirmer la capacité exacte de reconnaissance du complexe PARP-1/XRCC1/ADN ligase III.
Nouveaux membres de l’interactome
Une surexpression de la protéine a été observée dans certaines lignées tumorales (Ourliac-Garnier et al., 2009). ARF6 (ADP-ribosylation factor 6) est une GTPase qui participe à la mobilité cellulaire (recyclage endosomal de la E-cadhérine, élément des jonctions adhérentes), ainsi qu'à la régulation de la division (interaction avec le cytosquelette), notamment entre les cellules.
M ATÉRIEL ET M ÉTHODES
Description générale du dispositif
La biopuce SPRi
Le plan de dépôt A (5 x 5) a été utilisé pour déterminer les conditions expérimentales les plus favorables à la construction de biopuces puis aux études d'interactions protéiques. Toutes ces sondes ont été appliquées sur la puce à une concentration unique dans la solution d'application (10 µM), paramètre affectant la densité de greffage.
![Figure 53 : Assemblage de la séquence [sonde-cible lésée] à la surface du prisme SPRi](https://thumb-eu.123doks.com/thumbv2/1bibliocom/464979.70338/154.892.190.700.463.865/figure-assemblage-séquence-sonde-cible-lésée-surface-prisme.webp)
Dispositif expérimental
Une microstructuration de la surface peut alors être réalisée pour définir la zone de travail. Les signaux sont enregistrés et convertis pour produire des sensorgrammes représentatifs des événements se produisant à la surface de la biopuce (adapté de Scarano et al., 2010).

Protocoles détaillés
- Prismes
- Synthèse des séquences sondes et cibles
- Platination des cibles
- Dépôt des sondes sur le prisme par électropolymérisation
- Analyse SPRi
- Régénération totale de la biopuce
A la fin de la phase de dépôt, le prisme est rincé à l'eau, séché à l'argon et stocké à 4°C sous atmosphère d'argon. Les groupes de protéines commerciales ont des concentrations différentes (270nM - 5µM), ce qui implique une dilution de la protéine lors de la préparation de l'échantillon à injecter.

Optimisation des paramètres expérimentaux
Favoriser l’hybridation des sondes
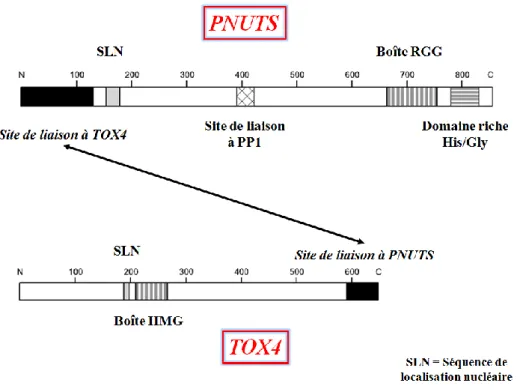
Cette fonctionnalité fait de TOX4 au sein du complexe PTW/PP l'élément potentiellement responsable de l'interaction avec les adduits. La figure 61 montre un exemple typique de signaux enregistrés lors de l'hybridation de cible sur le premier modèle de prisme avec ces conditions optimisées.
Analyse de l’interaction sondes/protéine
Comme la sonde cisplatine, la sonde simple brin présente une dissociation régulière conduisant à une perte d'1/3 du signal d'association. Cela pourrait impliquer que la dissociation observée était en réalité causée par une perte de protéines affectant les parties intactes de la sonde portant l'adduit.

Vérification de l’interaction d’HMGB1 avec les trois types d’adduits
Ensuite, différentes concentrations de la protéine HMGB1 ont été injectées séquentiellement sur la biopuce : 5 nM, 10 nM, 25 nM, 50 nM, puis à nouveau 10 nM pour assurer la reproductibilité des signaux en fin d'expérience. Grâce à la concentration en HMGB1 de 25 nM, nous avons réussi à atteindre le plateau de saturation pour toutes les sondes.
![Figure 63 : Conversion en image des signaux d’hybridation des constructions oligonucléotidiques cibles : (A) [ODN non lésé-Zip6C] ; (B) [ODN cisplatine-Zip7C] ; (C) [ODN oxaliplatine-Zip9C] ; (D) [ODN JM118-cisplatine-Zip7C]](https://thumb-eu.123doks.com/thumbv2/1bibliocom/464979.70338/172.892.289.727.627.1079/figure-conversion-hybridation-constructions-oligonucléotidiques-cisplatine-oxaliplatine-cisplatine.webp)
Évaluation des capacités de fixation de TOX4 sur les adduits platinés
Pour HMGB1, des signaux d'interaction très forts ont été enregistrés pour les sondes cisplatine (3,3 %) et JM118 (3,2 %). Le changement de réflectance observé pour les sondes oxaliplatine est le plus faible (2,7%), suivi de celui mesuré pour les sondes ODN intactes (2,4%).

Remarques générales sur la technique
Reproductibilité des interactions
L'hybridation des cibles sur les sondes greffées dépend de l'accessibilité de ces séquences, et est probablement réduite par l'existence de structures secondaires. De plus, réaliser l'étape d'hybridation à une température élevée (mais évidemment inférieure à la température de fusion des oligonucléotides) permet de favoriser la spécificité de l'ensemble sonde/cible et d'éviter les hybridations croisées.
Détermination des paramètres cinétiques de l’interaction
Les appareils commerciaux actuellement disponibles ne disposent pas de système de contrôle de la température. De ce fait, la concentration en protéines n’est pas égale après passage de la solution sur les sondes amont, ce qui altère l’interaction avec les sondes aval.
Utilisation de protéines recombinantes
Cependant, l'augmentation du débit a été obtenue lors de la dissociation des deux protéines, et en aucun cas nous n'avons observé de rupture de pente, caractéristique du phénomène de réassociation. En ce qui concerne la sélection de la protéine candidate, la sélection d’une protéine bloquante appropriée peut également s’avérer difficile.
Remarques générales sur les interactions observées
Affinité d’HMGB1 pour les sondes platinées
Les différences de déformation de la structure de l'ADN selon les adduits dues à l'encombrement stérique apporté par les fragments transporteurs oxaliplatine et cisplatine sont certainement responsables de cette affinité variable. Cependant, si une certaine affinité de la protéine (lot identique) pour la sonde simple brin a toujours été observée lors des expériences ultérieures (signaux supérieurs à ceux enregistrés pour les parcelles pyrrole), cette interaction ne s'est jamais révélée supérieure à celle observée pour le double brin. -sonde toronnée non endommagée par la suite.

Affinité de TOX4 pour les sondes platinées
Dans la troisième partie, nous avons voulu appliquer ces méthodes à un autre problème : la protéine DDB2 et son rôle en tant que membre de l'interaction avec les dommages à l'ADN. ÉTUDE DE LA PROTÉINE DDB2 EN TANT QUE MEMBRE DES INTERACTOMES ADDUITS CIPLATIN.
M ATÉRIEL ET MÉTHODES
Principe général et stratégie expérimentale
Cultures des lignées et tests de cytotoxicité
Lignées cellulaires
Le milieu est changé tous les trois jours et un passage hebdomadaire est réalisé en détachant les cellules à l'aide d'une solution de PBS 0,05% trypsine 10µM EDTA (Gibco Invitrogen) et en réensemençant à 20 000 cellules/ml (volumes utilisés : 5ml par boîte de 25cm² ; 15ml pour un Boite de 75 cm² ; 35 ml pour une boite de 175 cm²). Le comptage est effectué manuellement en colorant les cellules avec une solution de bleu trypan (Sigma) puis en les plaçant dans une cellule de comptage en verre Kova (Hycor Agilent Technologies).
Test de viabilité MTT
Après avoir retiré le PBS, 1,7 ml du milieu de culture d'origine sont replacés dans chaque boîte et les cellules sont incubées dans des conditions normales pendant 24 heures. Pour ce faire, 20 µl de réactif MTT sont ajoutés dans le puits et les cellules sont incubées pendant 2 heures.
Ligand fishing et protéomique
- Démarche expérimentale
- Exposition des lignées cellulaires au cisplatine
- Extraction des protéines nucléaires
- Préparation des sondes plasmidiques UV
- Capture et analyse des protéines
L'introduction de lésions photoinduites s'effectue par simple exposition de la solution d'ADN aux rayons UV. La vérification de l'efficacité de la fixation de l'ADN est à nouveau effectuée sur un gel d'agarose dont un exemple typique est visible dans la section.

SPRi
Mesure des capacités de réparation après un stress génotoxique
- Exposition des cellules au rayonnement UVB
- Extraction de l’ADN cellulaire
- Quantification des lésions UV
- Traitement au cisplatine
Le protocole de digestion de l'ADN est identique à celui déjà décrit pour la quantification des adduits de platine dans les plasmides sondes. La quantification des lésions a été réalisée de la même manière que pour les adduits de platine dans l'ADN plasmidique.

Mesure de l’expression de gènes
- Culture des cellules
- Extraction de l’ARN total
- Rétrotranscription
- PCR quantitative
La transcription inverse de l'ARN total en ADNc est réalisée à l'aide du kit SuperScript II Reverse Transcriptase (Invitrogen). La validation du gène domestique le plus adapté parmi les quatre disponibles est réalisée à l'aide de la matrice Excel BestKeeper© version 1.0 (Pfaffl et al., 2004).

Vérification de l’expression de DDB2 et sensibilité des lignées aux trois dérivés du platine
Détermination préalable de la sensibilité des lignées au trois dérivés du platine
Mise au point des sondes plasmidiques UV
Évaluation préalable par immunodétection de l’interaction de DDB2 avec les sondes plasmidiques
Interactome des adduits du cisplatine
Interactome des photoproduits UV
Vérification par SPRi de l’affinité de DDB2 pour les lésions des dérivés du platine
Évaluation des capacités de réparation des adduits de l’ADN selon le statut DDB2
Détermination de la dose d’exposition aux UV par test de cytotoxicité
Mesure de la cinétique de réparation des adduits
Réparation des photoproduits UVB
Réparation des adduits majoritaires du cisplatine
Suivi de l’expression de DDB2 au cours du temps
Tests de ligand fishing effectués à partir des extraits MDA MB 231
Interactome des lésions du cisplatine
Interactome des lésions engendrées par le rayonnement UV
Cinétique de réparation des lésions
Réparation des photoproduits
Réparation des pontages générés par le cisplatine
Cinétique d’expression de DDB2 suite aux traitements génotoxiques
Réponse de nos modèles cellulaires aux tests de cytotoxicité
Figures supplémentaires
Liste des communications et publications



![Figure 28 : Analyse par SPR de la dissociation de la protéine MutS avec un oligonucléotide portant un mésappariement de base (GG/CT) ou une lésion complexe [mésappariement + adduit 1,2-d(GpG) du cisplatine] (G*G*/CT) en l’a](https://thumb-eu.123doks.com/thumbv2/1bibliocom/464979.70338/83.892.153.691.111.367/analyse-dissociation-protéine-oligonucléotide-mésappariement-complexe-mésappariement-cisplatine.webp)

