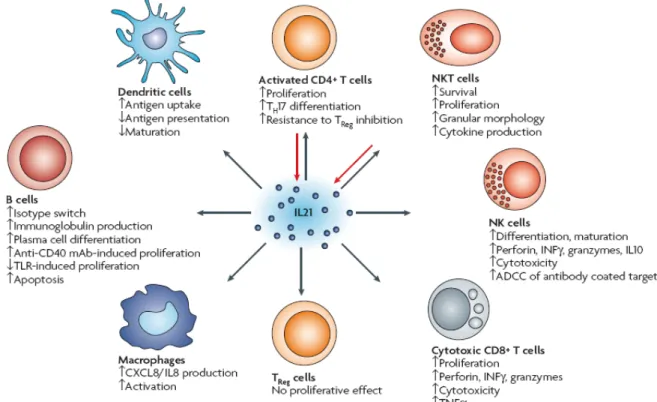
The increased number of plasma cells that occurred in the presence of IL21 and TLR9 agonists may reflect their contribution to the aberrant function of B cells and the large numbers of autoantibodies detected in SLE patients. In addition, the increase in the expression of CD86 may indicate the improved ability of B cells to activate naïve T cells. Presumably, the overproduction of IL21 and the enhanced TLR9 activation may act synergistically in the pathogenesis of SLE by inducing the differentiation of naïve B cells into plasma cells and enhancing B cell antigen-presenting capacity.
Innate and Adaptive Immunity
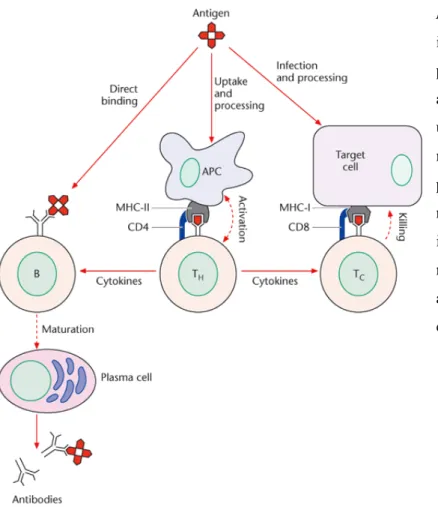
The cells of the immune system
Most cells of the body express MHC class I molecules and they are capable of presenting antigens to CD8+ T cells. The type of T cell response (tolerance versus immunity, Th1 versus Th2, Tregs versus Th17) that will occur following antigen presentation to T cells is related to the type and maturation stage of the dendritic cells, and the environmental signals they receive [53, 54]. After recognition of the pathogen, B cells differentiate into antibody-secreting plasma cells or memory B cells.
The cytokines
Analysis of the intracellular expression of TLR9 receptor in monocyte-derived dendritic cells from SLE patients and healthy individuals. IL21 and TLR9 synergistically promote the expression of antigen-presenting capacity markers on B cells from active SLE patients. IL21 and TLR9 may have synergistic stimulatory effect on plasma cell differentiation from naïve B cells into plasma cells in SLE patients with active disease.
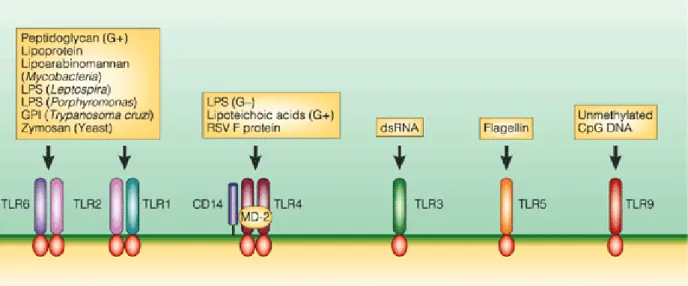
The Toll- Like Receptors
Self Tolerance and Autoimmunity
When this leads to tissue damage and deregulation of the immune system, the appropriate term is autoimmune disease. These diseases can be divided into two distinct categories, the organ-specific and the systemic autoimmune diseases. Apart from the involvement of the adaptive immune system in the breaking of tolerance, there are indications for the innate immune system participation.

Systemic Lupus Erythematosus
Pathophysiology of the disease
In the first phase, antigen presentation by APC is increased, and in the second phase, abnormal activation of autoreactive B and T cells follows. The final stage of development of pathogenesis involves the effects of autoreactive lymphocytes on target tissues. Toll-like receptors, which normally detect pathogens and trigger innate and sometimes adaptive immune responses, are one of the possible suspects for the pathogenesis of autoimmune diseases.
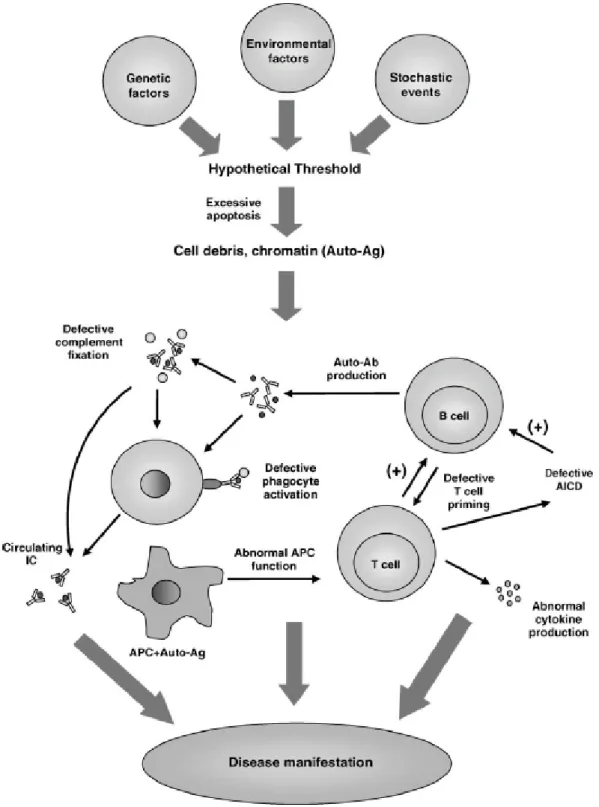
Defects in the action of phagocytic cells contribute to the prolonged availability of cell debris that can be phagocytosed and interpreted as foreign by the antigen-presenting cells. An abnormal titer of ANA by immunofluorescence or a similar assay at any time in the absence of drugs known to induce ANAs. Include cytoid bodies, retinal hemorrhages, severe exudates or choroidal hemorrhages, or optic neuritis.
One possible explanation is that intracellular autoantigens become 'visible' to the immune system during apoptotic cell death caused by environmental factors. The defective clearance of apoptotic cell debris may be a reason for the high abundance of autoantigens. T cells from SLE patients promote the activation of B cells to produce IgG in the absence of antigen or mitogen.
All of these mechanisms that have been described contribute to the perpetuation of autoimmune responses and tissue damage in SLE.
The role of TLR9 in the pathogenesis of SLE
This study confirms that TLR9 plays a key role in the production of antibodies against DNA and nucleoproteins [11]. In contrast to studies suggesting a negative role for TLR9 in murine lupus-like pathogenesis, there are studies suggesting the opposite [65]. Genetic loss of TLR9 signaling has been shown to result in accelerated disease progression and high mortality in lpr mice [ 62 , 31 ].
A prior error in the induction of tolerance and the clearance of apoptotic bodies results in the formation of DNA containing immune complexes (ICs). In response, plasmacytoid dendritic cells produce IFNα, which induces the expression of MHC and costimulatory molecules that enhance the antigen-presenting capacity of professional APCs. The antibodies that are produced perpetuate the formation of IC and create a vicious cycle in the pathogenesis of SLE.
SLE macrophages have a reduced ability to engulf apoptotic cells, which may contribute to the pathogenesis of lupus since newly presented intracellular antigens become apparent [59]. Recent research from our laboratory has shown that there is increased expression of TLR9 in CD14+ macrophages in active lupus patients compared to inactive patients and healthy controls [41]. Due to the small number in the peripheral blood and other accessible tissues and the high variability of the DC population, its manipulation is difficult.
Typically, the dendritic cells used in these studies are developed in vitro by monocytes that differentiate in the presence of GM-CSF and IL4 and mature in the presence of various stimuli such as lipopolysaccharide (LPS) and TNFα.

The role of IL21 in the pathogenesis of SLE
This decrease appears to be caused by cell migration into peripheral lymphoid tissues and sites of inflammation. It was recently suggested that human moDCs express TLR9 and they can mature in response to TLR9 stimulation using CpG ODN D19 [ 21 ]. The study of DCs in autoimmunity is of great importance as they have a central role in both the generation of immunity and tolerance.
The inability of DCs to clear autoreactive T cells, induce regulatory T cells, and their ability to present autoantigens and overproduce inflammatory cytokines is associated with the development of autoimmunity. Additional evidence for the involvement of IL21 in the pathogenesis of SLE was provided by the sanroque mutant mouse. These mice have enhanced levels of a population of follicular helper T cells that produce increased amounts of IL21.
The biological question and study hypothesis
The investigation of TLR9 and IL21 contribution in the pathogenesis of SLE was the aim of this study. The increased numbers of TLR9-expressing B cells and monocytes, as well as the increased IL21 levels in active SLE patients, found during this study raised many questions. Based on these observations, it was decided to investigate the potential synergistic effect of IL21 and TLR9 on several aspects of the biology of B cells, monocytes and dendritic cells, all antigen-presenting cell types, in SLE patients.
In addition, the effect of IL21 and TLR9 on the antigen-presenting capacity and differentiation of B cells into plasma cells and memory cells was assessed. The effect of TLR9 stimulation on monocytes was also evaluated and TLR9 expression was examined in SLE patients and healthy control moDCs. Isolation of human peripheral blood mononuclear cells (PBMCs) and measurement of IL21 and IL21R mRNA levels, from SLE patients and healthy individuals.
Comparative analysis of the B-cell differentiation markers (CD19, CD27 and CD38) for naive, memory and plasma cells after stimulation with combinations of IL21 and TLR9 ligand (ODN2006). Investigation of the antigen-presenting capacity of B cells from SLE patients and healthy individuals after culture with combinations of IL21 and ODN2006 and additional stimuli. Investigation of the effect of TLR9 and other stimuli on the antigen-presenting capacity of monocytes.
PATIENTS, MATERIALS AND METHODS
- Peripheral Blood
- Materials and Methods
- Cell isolation methods and cell cultures
- Flow cytometric analysis
- RNA isolation from peripheral blood mononuclear cells
- Reverse Transcription- Polymerase Chain Reaction (RT- PCR)
- Measurement of IL21 and IL21R mRNA levels expressed by SLE patients
- Active SLE patients display enhanced differentiation of naïve B cells into plasma cells
- IL21 and TLR9 stimulation promote the expression of co-stimulatory molecules on B
- TLR9 stimulation of monocytes
- TLR9 is not expressed in monocyte- derived dendritic cells
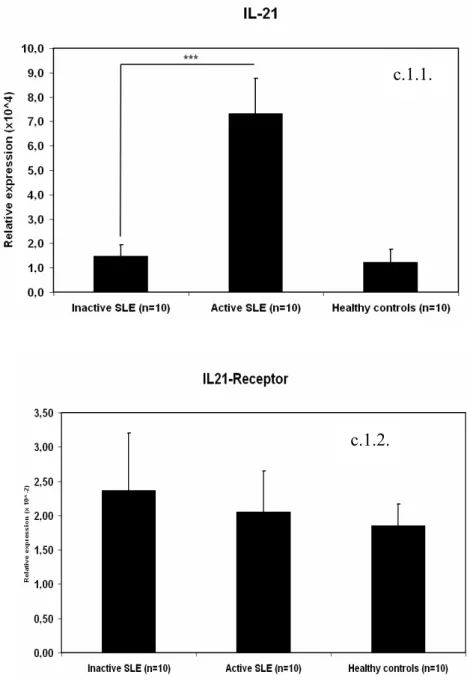
Subsequent real-time PCR showed that IL21 mRNA expression was significantly higher in patients with active SLE compared to SLE patients with inactive disease (p=0.002) and healthy controls (p=0.003) (Figure c.1.1 ). In contrast, no significant differences in IL21R expression were evident between SLE patients and healthy controls, as shown in Figure c.1.2. C2. Patients with active SLE show increased differentiation of naïve B cells into plasma cells after stimulation with IL21 and TLR9 ligand.
Isolated B cells from SLE patients with active and inactive disease were cultured for 7 days with IL21, TLR9 and TLR3 ligands and their combinations and the phenotype of B cells were finally assessed by flow cytometry. The differentiation of naïve B cells from active SLE patients to plasma cells is remarkable (Figure c.2.2.) compared to the almost zero induction of plasma cells by inactive SLE patients. With either stimulus, the percentages of memory B cells generated are higher in SLE patients with active disease.
Notably, the combination of TLR9 and IL21 stimulation results in a significantly higher induction of memory B cells in patients with active SLE (10%) compared to cells from inactive patients (5%). The combination of both stimuli promotes enhanced induction of plasma cells by naïve B cells in patients with active SLE. CD86 expression appears to increase dramatically upon IL21 and TLR9 stimulation, more so than other stimuli or combinations of stimuli, on B cells from patients with active SLE.
Monocytes from SLE patients and healthy controls were magnetically isolated from PBMC and flow cytometry was performed after 24 h of culture. TLR9 is not expressed on monocyte-derived dendritic cells in both SLE patients and healthy controls. To assess TLR9 expression in moDCs, monocytes from SLE patients and healthy controls were isolated and differentiated into dendritic cells after 7 days in cultures with GM-CSF and IL4.
DISCUSSION
- IL21 expression in SLE patients
- IL21 and TLR9 promote differentiation of human B cells
- IL21 and TLR9 affect the antigen presenting capacity of B cells from active SLE
- TLR9 stimulation of monocytes
- TLR9 expression in moDCs
- Conclusions and future plans
In summary, based on the findings of this study, IL21 expression is increased in SLE patients with active disease. The increased expression of TLR9 and IL21 in SLE patients with active disease is proposed to have functional consequences. The increased differentiation in plasma cells in the presence of IL21 and TLR9 agonists may be responsible for the aberrant function of B cells and the large numbers of autoantibodies detected in SLE patients.
The fact that IL21 expression is significantly increased in SLE patients with active disease may reflect a causal role of this cytokine in the aberrant function of some cell types. Given that IL21 and TLR9 induce the differentiation of naïve B cells into memory B cells and plasma cells in healthy individuals, the effect of these two stimuli on B cells from SLE patients was assessed. Here it is shown that IL21, in combination with TLR9 stimulation can lead to robust differentiation of B cells into plasma cells in active SLE patients.
This may mean that tolerance breakdown and changes in the immune system in active SLE patients make naïve B cells more vulnerable to the effect of IL21 and TLR9 on plasma cell differentiation. The antigen-presenting capacity of B cells from SLE patients after stimulation with TLR9 and IL21 was assessed. The fact that IL21- and TLR9-treated B cells from SLE patients have increased capacity to activate and induce proliferation of T cells may explain the increased activation of T cells in this autoimmune condition.
TLR9, which is upregulated upon BcR involvement, is increased in SLE patients, likely due to the high abundance of autoantigens.