After interaction of the extracellular domain of the receptor with the ligands, a proteolytic step is activated, leading to the release of the active intracellular part of the N receptor (Nic). In addition to transcription factors, a variety of proteins post-transcriptionally control the precise activation of the pathway in time and space; these include proteases, glycosyltransferases, and E3 ubiquitin ligases. In this work, the role of N-ligands, Dl and Ser, as well as their two associated ubiquitin ligases, Neuralized (Neur) and Mind bomb1 (Mib1), was studied in N-pathway activation in Drosophila.
We showed that Neur is an intracellular membrane-bound protein that targets the N Dl ligand for endocytosis and degradation in a RING-dependent manner. In nerve loss-of-function mutations, Dl endocytosis is blocked, and the opposite is true when nerve is ectopically expressed (increased endocytosis). Next, the role of the two N ligands was studied during the lateral inhibition process and it was shown that, contrary to previous belief, Dl and Ser act synergistically during this process (Dl was thought to be the only active ligand there), and the regulation their transcription is not important for this function.
We observed that NeurΔR is active in signal sending but inactive in signal reception. This leads us to interesting findings regarding the function of the wild-type Neur protein; Neur may have a dual function during N signaling: it may activate signaling in a RING-independent manner, and it may also allow cells to receive the N signal in a RING-dependent manner (e.g., by reducing the cis- negative effect of the high concentration of ligands).
ΤΟ ΣΗΜΑΤΟΔΟΤΙΚΟ ΜΟΝΟΠΑΤΙ NOTCH
ΡΥΘΜΙΣΗ ΤΗΣ ΣΗΜΑΤΟΔΟΤΗΣΗΣ NOTCH 1. H πρωτεόλυση στην σηµατοδότηση Notch
Ουβικουϊτινυλίωση και σηµατοδότηση Notch (βλέπε “Συζήτηση”)
ΤΡΟΠΟΙ ΧΡΗΣΗΣ ΤΗΣ ΣΗΜΑΤΟΔΟΤΗΣΗΣ NOTCH
Επειδή το Ν έχει ανασταλτική επίδραση στη μοίρα του κυττάρου σε αυτή την περίπτωση, το μοναδικό κύτταρο που επιλέγεται ως πηγή σήματος είναι το μόνο που διαφεύγει της αναστολής (Heitzler and Simpson, 1991; Kopan and Turner, 1996; Heitzler et al., 1996). ). PNC: Proneural Cluster, SOP: Sensory Organ Precursor, PAO).
ΤΟ ΓΟΝΙΔΙΟ neuralized
Εκτός από το D. melanogaster neur, ομόλογα του γονιδίου έχουν κλωνοποιηθεί και σε άλλα έντομα, τόσο στην οικογένεια Drosophilidae όσο και στο κουνούπι Anopheles gambiae, τη μέλισσα Apis melifera και επιπλέον στον άνθρωπο (Nakamura et al., 1998). στο ποντίκι (Pavlopoulos et al., 2002) και στο νηματοειδές C.elegans (Wilson et al., 1994).
ΤΟ ΓΟΝΙΔΙΟ mind bomb
Aποµόνωση πλασµιδιακού DNA
Παραγωγή αντισωµάτων έναντι της πρωτεΐνης Neuralized 4α. Συνθήκες επαγωγής χιµαιρικής πρωτεΐνης His-Neur1050
Για την έκφραση των πρωτεϊνών Neur και NeurAR, τα πλασμίδια pUAST-neur ή pUAST-EGFPneur και pUAST-neurARGFP, αντίστοιχα, χρησιμοποιήθηκαν παρουσία του πλασμιδίου pMt-Gal4 και η επιμόλυνση ακολουθήθηκε από επαγωγή με 0,7 mM CuSO4 (στέλεχος 70 mM) CuS04).
Aνάλυση Western
Ανάλυση RNA
Eκχύλιση RNA και Aντίστροφη Mεταγραφή ακολουθούµενη από PCR α. Eκχύλιση RNA από προνύµφες Δροσόφιλας
Υβριδοποίηση in situ µε RNA ανιχνευτή
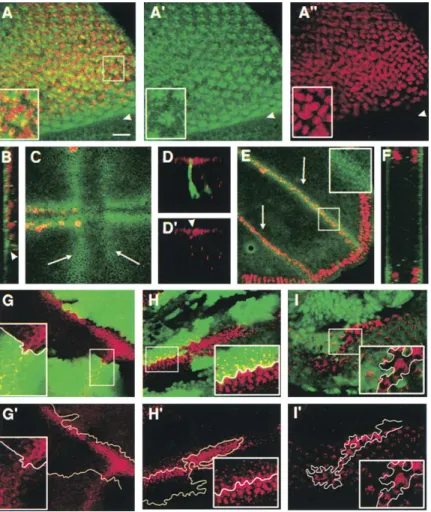
Ο φαινότυπος που παρουσιάζεται από το Nec μετά την υπερέκφραση του NeurΔR παρατηρείται επίσης σε άλλες περιπτώσεις όπου οι συνδέτες υπερεκφράζονται στην πλασματική μεμβράνη (Εικ. 3.30), όπως σε μιτωτικούς κλώνους mib1- (Εικ. 3.30A), mib1-+ UAS-neurΔR (Εικ. 3.30Ε), mib1- +UAS-Dl UAS-neurAR (Εικ. 3.30D, F), mib1-+UAS-Dl (Εικ. 3.30C) ανεξάρτητα από την παρουσία του NeuAR. Μιτωτικοί κλώνοι του Dl-Ser-δέσμευσης (A, B) υπερεκφράζοντας νευρώνα (C, D) ή του ανεπαρκούς neurAR (E, F) επισημαίνονται με πυρηνική GFP. Α) κλώνοι mib1, (Β) κλώνοι υπερέκφρασης UAS-neurΔR (υπό έλεγχο του actGal4), (C) κλώνοι mib1 που υπερεκφράζουν UAS-DlV5, (D και F) κλώνοι mib1 που υπερεκφράζουν UAS-DlV5, UAS -neurΔR με D που αντιστοιχεί σε περισσότερα κορυφαίες τομές, (Ε) κλώνοι mib1 που υπερεκφράζουν το UAS-neurAR.
Neuralized και Ενδοκύττωση
Πλευρική Αναστολή - Ο ρόλος των δεσµευτών του Notch
Ουβικουϊτινυλίωση και σηµατοδότηση Notch
Ο ρόλος του δακτύλου RING
Activation and function of Notch at the dorsoventral boundary in the Drosophila wing imaginal disc. Cytosolic interaction between Deltex and Notch ankyrin repeats implicates Deltex in the Notch signaling pathway. The Notch signaling pathway is required for enhancer of split bHLH protein expression during neurogenesis in the Drosophila embryo.
The identity of the sense organs in the Drosophila antenna is specified by the expression of the pronural atonal gene. Ligand-receptor interactions and transendocytosis of Delta, Serrate, and Notch: members of the Notch signaling pathway in Drosophila. Reversible blockage of membrane repair and endocytosis in the garland cell of the Drosophila melanogaster temperature-sensitive mutant, shibirets1.
Two different E3 ubiquitin ligases have complementary functions in the regulation of Delta and Serrate signaling in Drosophila. Identification of a human homolog of the Drosophila neuralized gene within the 10q25.1 malignant astrocytoma deletion region. Regulation of achaete-scute gene expression and sensory organ patterning in the Drosophila wing.
The Serrate locus of Drosophila and its role in imaginal wing disc morphogenesis: control of cell proliferation.
The interplay between DSL proteins and ubiquitin ligases in Notch signaling
Similarly, during embryonic neuroblast lateral inhibition, Ser is not expressed, making the process exclusively Dlaf-dependent (Gu et al., 1995). We and others have previously shown that Dl activity is augmented by its association with Neuralized (Lai et al., 2001; . Pavlopoulos et al., 2001). In fact, recent data suggested that Neur may act primarily on Dl, whereas Mib1 may act primarily on Ser (Le Borgne et al., 2005).
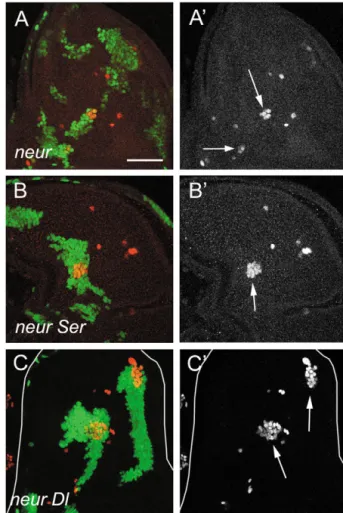
In both cases the phenotype improved (Fig. 2B,C; Table 1), suggesting that either Dl alone (in Ser clones) or Ser alone (in Dl clones) is more active in the presence than in the absence of neur+. This response of endogenous and overexpressed Ser to Neur is identical to that previously described for Dl (Lai et al., 2001; Pavlopoulos et al., 2001). The Drosophila gene disruption project ( Bellen et al., 2004 ) has generated a P element insertion, EY9780, that disrupts the mib1 gene in the 5⬘ UTR.
When we co-expressed Neur with either DSL protein in mib1 clones, the ligands regained their ability to induce Wg (Fig. 6C,F); indeed, Dl was hyperactivated, as it is when co-expressed with Neur in a wild-type background (Pavlopoulos et al., 2001). Therefore, Neur can substitute for the lack of Mib1 activity during wing DV boundary specification, consistent with recent reports (Le Borgne et al., 2005; Wang and Struhl, 2005). Dl transcriptional upregulation in the absence of Notch signaling in proneural fields occurs (Koelzer and Klein, 2003; Schweisguth and Posakony, 1994), but this modulation does not appear to be a prerequisite for the specification of the wild-type neural precursor, at . at least in the case of macrochaetes (this work) and embryonic neuroblasts (Seugnet et al., 1997b).
It is possible that the genetically detected N-Dl negative feedback loop may reflect Dl and N activity rather than transcription, although a transcriptional input has been documented (Heitzler et al., 1996). Although Neur was known to affect Dl localization and function in some cases (Lai et al., 2001; In the present work, we added the last pair, Neur-Ser, using all of the above assays.
In the ubiquitin-dependent endocytosis pathway, many of the adapter proteins are themselves ubiquitinated, possibly favoring the formation of interconnected cargo-adaptor complexes ( Polo et al., 2002 ); An even more puzzling observation in light of our model is that some DSL proteins in C. secreted mutants of Drosophila Dl and Ser act as Notch antagonists (Mishra-Gorur et al., 2002; . Sun and Artavanis-Tsakonas, 1997 ), consistent with a requirement for endocytosis in DSL signaling.