RÖVIDÍTÉSEK JEGYZÉKE
BEVEZETÉS
- A gamma-hidroxi vajsav (GHB) története, felhasználása, szervezetre
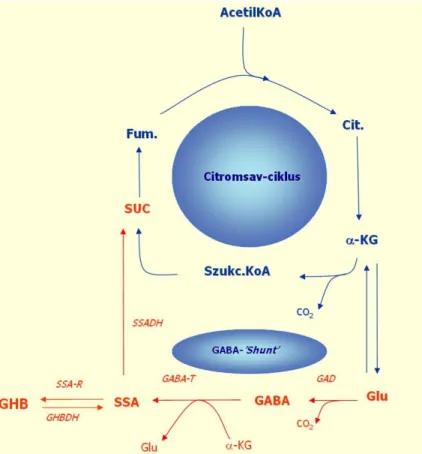
- A GHB és borostyánk ő sav (SUC) szerepe a szervezetben,
- A GHB és a GABA B rendszer
- A NAc agyszöveti struktúra jellemzése, feltételezett szerepe a GHB
A GHB-vel szedált áldozatokkal szembeni erőszakos cselekmények száma világszerte növekszik (ElSohly és Salamone, 1999; .. Jones, 2001), bár a használatot nehéz utólag megmondani a vérből (Wood et al. 2004; Carbon atoms szürkék, az oxigénatomok pirosak, míg a hidrogének a fehér hippocampusban 1,2 µM, a striatumban 1,8 µM, a kéregben 0,4 µM Vayer et al. 1988). A GHB mitokondriumoktól független katabolizmusát a NADPH-függő citoplazmatikus aldehid-reduktáz (ALR-1) enzim végzi (Kaufman és Nelson, 1981; Kaufman és mtsai. 1983; Kaufman és Nelson 1987).
A GHB-vel (100-300 mg/kg i.v.) végzett kezelés az ischaemiás állapot előidézése előtt és/vagy után patkányokban nagymértékben növelte a túlélő neuronok számát, az állatok mozgás- és tanulási képességét a kezeletlen állatokhoz képest (Ottani et al. 2004) A GHB-t sokáig emlegették a szakirodalomban, mint gyenge GABAB receptor agonistát (Hechler és mtsai 1997; Mathivet és mtsai 1997; Lingenhoehl et al. 1999; Castelli és mtsai 2003). Wu és mtsai. 2004) Olyan eredmények születnek, amelyek kétségbe vonják a GABAB receptortól független GHB hatásmechanizmus létezését (Jensen és Mody, 2001; Ren és Mody, 2003; Cortez és mtsai. 2004; Barbaccia és mtsai. 2005).
A klónozott, GABAB-receptor-független, GHB-receptor-izoform-struktúrák jelenlétét humán frontális kéreg-cDNS-könyvtárból is leírták (Andriamampandry et al. 2007). A kokain beadása következtében a connexinek (Cx36), a neuronok közötti réskapcsolatokat építő fehérjék sejtfelszíni expressziója (Bennett és mtsai 1999; McCracken és mtsai 2005), valamint az MSN-ek dendrit tüskék száma és mérete a kokain hatása alatt is jelentősen megnő.
CÉLKITŰZÉSEK
KÍSÉRLETI MÓDSZEREK
Kísérleti állatok, humán agyminták
Felhasznált oldatok és vegyszerek
számú határozata alapján a gamma-hidroxi-vajsav (GHB) a rendőrségi nyilvántartásban szereplő pszichotróp anyag (P2). Az Egészségügyi Engedélyezési és Közigazgatási Hivatal Gyógyszerigazgatási Hivatalának 4551/2005/KÁB rendelete alapján került felhasználásra. számú engedélye alapján. KAB.
Szubcelluláris frakciók el ő állítása patkány el ő agyból, humán NAc és
A Nac- és GP-tisztított szinaptoszomális membránfrakciók előállításához humán agymintákat dolgoztunk fel a patkány előagyi szubcelluláris frakcióinál leírtakkal megegyező módon (Molnár et al. 2006; Molnár et al. 2008a).
Szubcelluláris frakciók köt ő dési és transzport folyamatainak
- GHB és SUC telítési kötődés
- Jelzett [ 3 H]GHB és [ 3 H]SUC helyeken specifikusan kötődő
A Nac- és GP-tisztított szinaptoszómális membránfrakciók előállításához emberi agymintákat dolgoztunk fel az egér előagyi szubcelluláris frakcióinak előállításánál leírtakkal megegyező módon (Molnár et al. 2006). A minták radioaktivitását HiSafe3 (0,9 ml) szcintillációs keverékben (LKB-Wallac, Bromm, Svédország) mértük folyadékszcintillációs műszerrel (Wallac 1414 WinSpectral, Perkin-Elmer Life Sciences, Turku, Finnország). kétszer, az adatok legalább három különböző készítményből származnak.
Az egyensúlyi kötési paramétereket (a kötőhelyek maximális száma: BMAX; az egyensúlyi disszociációs állandó: Kd) az OriginPro 7.5 programba beépített egy- és kétkötőhelyes telítési görbék illesztésével határoztuk meg. Az egyes ligandumok féltelítési koncentrációját (IC50) az OriginPro 7.5 programmal határoztuk meg, egy vagy két kötőhelyes gátlási görbére illesztve. A [3H]SUC-felvételt a következőképpen határoztuk meg (Molnár et al. 2008a; Héja et al. 2004): a szinaptoszóma készítmény mintáit (550 µl) előinkubáltuk 'PBSS' pufferben (550 µl GHB) oldott különböző vegyületekkel, SUC vagy CIT) (10 perc, 30 °C).
Ugyanennek a vegyületnek a [3H]SUC-felvételre gyakorolt hatásának meghatározására kísérletsorozatot végeztünk legalább 3 preparátummal, minden inkubációt kétszer megismételve. Az adatokat az alapvonali emisszióhoz illesztett exponenciális görbével normalizáltuk, átlag ± S.D. (OriginPro 7.5), 5-7 különböző készítményből megismételve.
Ca 2+ ion tranziensek detektálása patkány NAc agyszeletben konfokális
- GHB-vel kiváltott Ca 2+ ion fluktuáció
- ATP-vel kiváltott Ca 2+ ion-hullámok
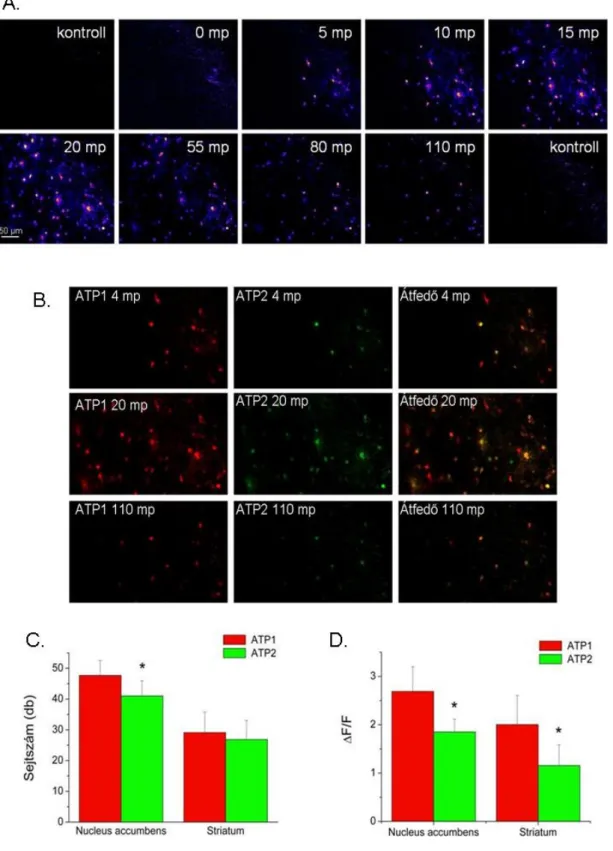
Ca2+ ion tranziensek kimutatása patkány NAc agyszeletekben lézeres konfokális képalkotó technikával és Ca2+ ionérzékeny fluoreszcens festékkel. A Fluo-4 AM intenzitása arányos az intracelluláris Ca2+ ionok koncentrációjával, de az intenzitás mértéke függ a sejtbe jutott festék koncentrációjától is. Ennek érdekében ATP-indukálta Ca2+-ion hullámokat generáltunk, amelyek gap junctionokon keresztül terjednek egér NAc agyszeletekben, majd követtük a SUC hullámokra gyakorolt hatását.
Az ATP által kiváltott Ca2+ ionhullámok kimutatására a 4.4.1. pontban leírtak szerint előkészített és megfestett egér agyszeleteket használtunk, amelyeket konfokális lézer pásztázó mikroszkóppal (Olympus BX50WI, Olympus, Budapest, Magyarország) követtünk. A sejtek Ca2+-ionjainak intracelluláris koncentrációjának változását az idő függvényében két területen, a NAc-ben vagy az azonos agyszelet striatumának mediodorsalis caudate putamen területén (továbbiakban striatum) követtük (2 mp/ felvétel). A Ca2+ ion hullámokat 100 µM 'modACSF'-ben oldott ATP váltotta ki, amelyet 2-5 MΩ ellenállású üveg mikrokapillárisból adtunk a szelet felületére, mindkét felvétel 3 .
Egyes méréseknél propidium-jodidos (PI) festést is alkalmaztak az ATP által kiváltott Ca2+ ionhullámok során fellépő sejthalál mértékének meghatározására. Sulforhodamin 101 (SR101) festéssel azonosítottuk az ATP-indukált Ca 2+ ion hullámokban részt vevő sejteket.
Immunhisztokémiai sejtazonosítás
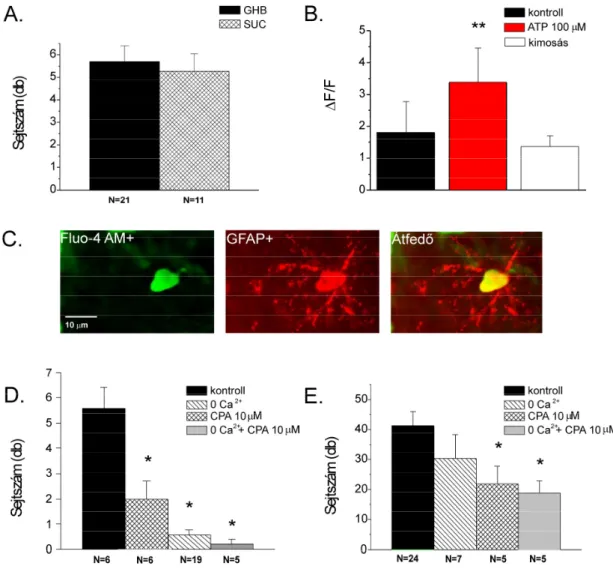
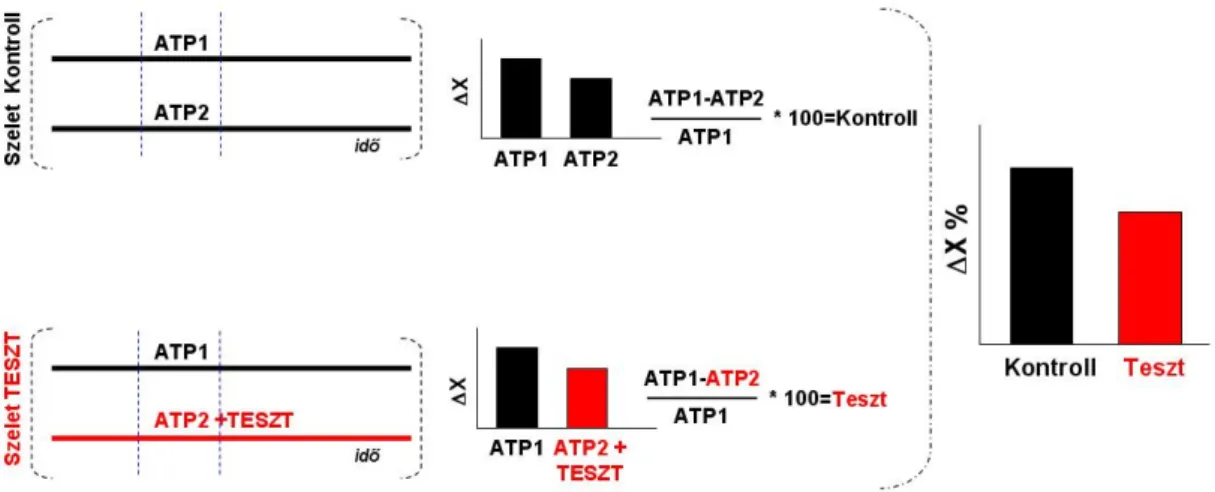
Az SR101 specifikusan jelöli a protoplazmatikus asztrocitákat, amelyek bizonyos glutaminsav transzportereket (EAAT) expresszálnak (Matthias et al. 2003; Kafitz és mtsai 2008). A Fluo-4 AM és SR101 festődés kolokalizációjának meghatározásához az 1. és 2. csatornában észlelt jeleket egymásra helyeztük (ImageJ Free Image Analysis Program 1.41, NIH, Amerikai Egyesült Államok). A szignifikancia meghatározásához az OriginPro 7.5 program független (kontroll vs. teszt) vagy páros (ATP1 vs. ATP2) t-tesztjét használtuk.
Másnap, 3x10 perces PB-ben való mosás után a lemezeket 1 órán át 0,5%-os Triton-X 100-ban tartottuk a membrán permeabilitásának növelése érdekében. PB-vel történő mosás után a metszeteket primer antitestet (nyúl anti-GFAP, 1:4000 azid PB-ben) tartalmazó szérumban inkubáltuk 48 órán keresztül, folyamatos enyhe rázatás mellett 4°C-on. A harmadik napon, 3x10 perces „PB” mosás után, a lemezeket a fluoreszcensen jelölt másodlagos antitestet (anti-nyúl IgG Alexa Fluor 594 konjugátum, 1:1000 PB-ben) tartalmazó szérumba helyeztük 1 órára, folyamatos gyengéd kezelés mellett. rázva, szobahőmérsékleten.
Az utolsó 'PB' mosás után a szeleteket egy tárgylemezre helyeztük, és a fedőlemezeket (Menzel-Glaser 24x40 mm) Aqua PolyMounttal szereltük fel. A korábban mért Fluo-4 AM és GFAP immunpozitivitás kolokalizációjának meghatározásához az 1. és 2. csatornában észlelt jeleket egymásra illesztjük (ImageJ Free Image Analysis Program 1.41, NIH, Amerikai Egyesült Államok).
Mez ő potenciál detektálása patkány NAc agyszeletben
Statisztikai kiértékelés és ábrakészítés
- SUC és GHB hatása ATP-vel kiváltott Ca 2+ ion-hullámra
- Az ATP-vel kiváltott Ca 2+ ion-hullámot jellemző paraméterek
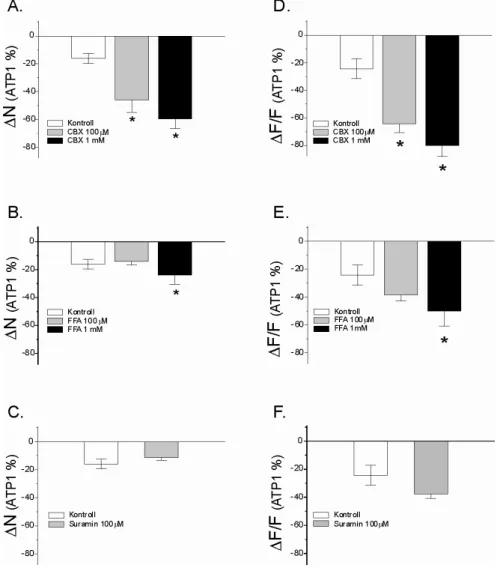
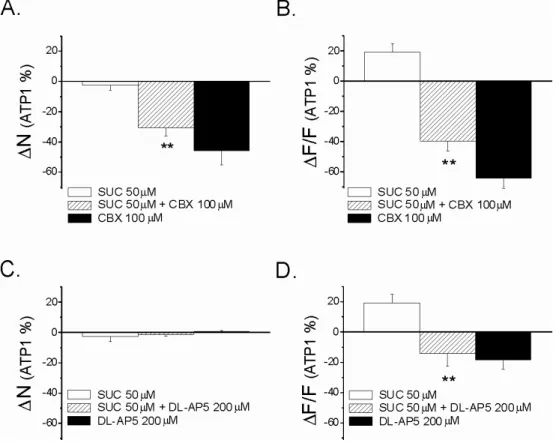
Nagy felbontású (2048x2048 pixel) képet készítettünk 2000-szeres nagyítással egy olyan sejtről, amely részt vesz a mindkét ATP dózis által generált Ca2+ ionhullámban (20.A ábra). Ezért megvizsgáltuk a CBX, az FFA és a suramin hatását az ATP-indukált Ca2+ ion hullámokra NAc agyszeletekben. A sejtszám (N) és a fluoreszcencia (∆F/F) változása, amely a két ATP-indukált Ca2+ ionhullám között következik be, az első Ca2+ ionhullám.
A sejtszám (N) és a fluoreszcencia változása (∆F/F) a két ATP-indukált Ca2+ ion hullám között az első Ca2+ ionhullám %-ában mutatkozik meg a kontroll mérésekhez képest (független t-teszt, p< 0,05). A 200 µM DL-AP5 nem befolyásolta az ATP által indukált Ca2+ ionhullámokban részt vevő sejtek számát, de csökkentette a résztvevő sejtek fluoreszcenciáját. Néhány agyszeleten a Ca2+ ionhullám kimutatása után a GFAP gliamarker immuncitokémiai reakcióját végeztük el a hullámban részt vevő sejtek azonosítására (N=4).
A Ca 2+ ion készletek hatását a Ca 2+ ion hullám terjedésére a 4.4.2 fejezetben leírt protokoll szerint végeztük.
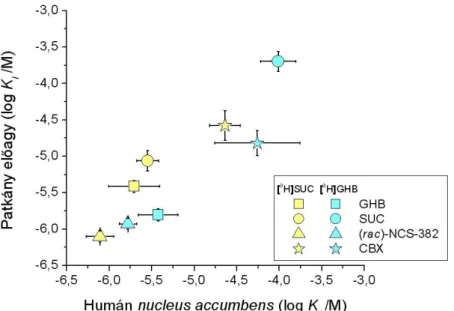
![1. táblázat. Humán NAc és GP tisztított szinaptoszómális membránfrakcióban mért, különböző ligandumok [ 3 H]GHB leszorítási adatainak összehasonlító táblázata](https://thumb-eu.123doks.com/thumbv2/9dokorg/19309715.0/37.892.160.765.265.700/táblázat-tisztított-szinaptoszómális-membránfrakcióban-különböző-leszorítási-összehasonlító-táblázata.webp)
MEGBESZÉLÉS
- A SUC-szenzitív GHB köt ő hely
- A GHB-szenzitív SUC köt ő hely
- A GHB köt ő hely és a nagy-affinitású SUC köt ő hely egy és ugyanaz
- A GHB GABA B receptor független hatása NAc asztrociták
- SUC hatása ATP-indukálta Ca 2+ ion-hullámokra
Ennek kiderítésére egér NAc agyszeletekben követtük a sejt Ca2+ ion koncentrációjának változását a GHB beadása során. A farmakológiai szerek által kiváltott asztroglia Ca2+-ion-tranzienseket korábban in vitro (Parri et al. 2001; Newman, 2003; Fields, 2004) és in vivo (Hirase és mtsai, 2005) is leírták. A Ca2+ ion oszcillációjának egyik oka lehet az asztrocita felszabaduló transzmitterek (ún. gliotranszmitterek, pl. Glu) idegsejtekre kifejtett stimuláló hatása (Haydon, 2001; Volterra és Meldolesi, 2005; Santello és Volterra Bergersen, 2009); és Gundersen).
Ennek ellenére az asztrociták Ca2+-ion-koncentrációjának növekedése nem feltétlenül befolyásolja a neuronok aktivitását (Fiacco et al. 2007), sőt eredményeink azt is mutatják, hogy a neuronok aktivitása nem változik a NAc-ben a GHB hatására (Molnár et al. 2009a). Munkánk során GABABR1 KO egerek NAc agyszeleteiben demonstráltuk a GHB hatását az asztrocita intracelluláris Ca2+ ion oszcillációjára. A terjedő Ca2+-ionhullámokat in vitro modellben, asztrocita (Cortina et al. 1998; Arcuino és mtsai 2002), agy fehérállományában (Schipke és mtsai 2002) és szürkeállományban (Shaltz et al. 2002) mutatták ki. ) ; Somjen et al. 2001; Peinado, 2001). szeletekben is.
Ilyen esetben a kiváltó inger (pl. mechanikai inger, elektromos inger, ATP) megnöveli a Ca2+ ionok intracelluláris koncentrációját, ami ATP felszabadulását váltja ki a gap junctionalis félcsatornákon keresztül (Hassinger et al. 1996; Arcuino et al. 2002; Bennett et al. 2003). Kimutatták, hogy a Ca2+-ionhullámok terjedési módjai egymás mellett vagy párhuzamosan is jelen lehetnek (Paemeleire et al. 2000). A SUC és a gap junctions közötti lehetséges funkcionális kapcsolat meghatározásához megvizsgáltuk a SUC hatását az ATP-indukált Ca2+ ionhullámokra.
Megállapítottuk, hogy a SUC koncentrációfüggő módon befolyásolja a Ca2+ ionhullámban résztvevő sejtek számát és intenzitását. A Ca2+ ionhullám terjedésének irányát nem befolyásolta sem az ATP szállítás iránya, sem a perfúziós rendszerben áramló folyadék (modACSF) iránya. Mindezek alapján elmondható, hogy bár a saját rendszerünkben mindkét Ca2+ ion hullámterjedési mód jelen lehet, modellünkben az intracelluláris terjedési mód követhető.
Ugyanakkor a GHB-nek nagyobb koncentrációban nem volt jelentős hatása a Ca2+ ionhullámokban résztvevő sejtek számára és azok intenzitására. A Ca2+ ionhullámot jellemző paraméterek összehasonlító vizsgálata kimutatta, hogy a SUC (50-200 µM) és a GHB (50 µM) alacsony koncentrációja jelentősen eltérő hatással van a hullámban résztvevő sejtek számára és intenzitására. A nagy mennyiségű felszabaduló glutaminsav nemcsak a Ca2+-ionok intracelluláris koncentrációját befolyásolja az asztrocitákban, hanem a neuronok aktivitását is.
ÖSSZEFOGLALÁS
SUMMARY
IRODALOMJEGYZÉK
Brancucci A, Berretta N, Mercuri NB, Francesconi W. 2004) Gamma-hydroxybutyrate and ethanol suppress spontaneous excitatory postsynaptic currents in dopaminergic neurons of the substantia nigra. Cash CD, Rumigny JF, Mandel P, Maitre M. 1981) Enzymology of the alternative reductive pathway of GABA catabolism leading to the biosynthesis of gamma-hydroxybutyrate. Castelli MP, Mocci I, Pistis M, Peis M, Berta D, Gelain A, Gessa GL, Cignarella G. 2002) Stereoselectivity of NCS-382 binding to gamma-hydroxybutyrate receptor in the rat brain.
1999) Prevalence of drugs used in alleged assault cases. 1996) Gamma-hydroxybutyric acid reduces thalamic sensory excitatory postsynaptic potentials through an action on presynaptic GABA B receptors. 1996) Gamma-hydroxybutyrate is not a GABA agonist. 2007) Astrocytes regulate neuronal excitability in the nucleus accumbens. 1977) Kinetics of in vivo conversion of gamma-[3H]aminobutyric acid to gamma-[3H]hydroxybutyric acid by rat brain. Kaufman EE, Nelson T, Miller D, Stadlan N. 1988) Oxidation of gamma-hydroxybutyrate to succinic acid semialdehyde by a mitochondrial pyridine nucleotide-independent enzyme.
Evidence for the participation of a cytosolic NADP+-dependent oxidoreductase in gamma-hydroxybutyrate catabolism in vivo. Laborit H, Jouany JM, Gerard J, Fabiani F. 1960) Generalities regarding the experimental study and clinical use of gamma-hydroxybutyrate. Levy MI, Davis BM, Mohs RC, Trigos GC, Mathé AA, Davis KL. 1983) Gamma-hydroxybutyrate in the treatment of schizophrenia.
Comparison of the effects of gamma-hydroxybutyrate and gamma-butyrolactone on cerebral carbohydrate metabolism. 1998 ) Gamma-hydroxybutyrate is a GABAB receptor agonist that increases potassium conductance in rat ventral tegmental dopamine neurons. The role of gamma-hydroxybutyric acid in the treatment of alcoholism: from animal to clinical studies. Snead OC 3rd, Bearden LJ, Pegram V. 1980) Effect of acute and chronic administration of anticonvulsants on endogenous gamma-hydroxybutyrate in rat brain.
In vivo conversion of gamma-aminobutyric acid and 1,4-butanediol to gamma-hydroxybutyric acid in rat brain. Vayer P, Ehrhardt JD, Gobaille S, Mandel P, Maitre M. 1988) Distribution of gamma-hydroxybutyrate and turnover rates in specific regions of the rat brain. New cyclic analogues of gamma hydroxybutyrate (GHB) with high affinity and stereoselectivity of binding to GHB sites in rat brain.
SAJÁT PUBLIKÁCIÓK JEGYZÉKE
Az értekezés témájához kapcsolódó közlemények
Az értekezés témájához szorosan nem kapcsolódó közlemény
KÖSZÖNETNYILVÁNÍTÁS
![13. ábra. A SUC, a GHB és a CIT [ 3 H]SUC felvételre és [ 14 C]SUC kibocsátásra gyakorolt hatásai](https://thumb-eu.123doks.com/thumbv2/9dokorg/19309715.0/42.892.154.772.595.819/ábra-suc-ghb-cit-felvételre-kibocsátásra-gyakorolt-hatásai.webp)