On the other hand, no phenotypic variation was observed in the generations exposed to CuSO4·5H2O and Cu(OH)2nano-pesticides. On the other hand, no phenotypic variation was observed in the generations exposed to CuSO4.5H2O and Cu(OH)2 nano-pesticides. On the other hand, the locomotor activity in the nanocapsules (2.5 mM CaCO3) used during the preparation of the validamycin nano-pesticide was determined.
On the other hand, the locomotor activity in the nanocapsules (2.5 mM CaCO3) used during the preparation of the nanopesticide validamycin was determined to be 89 ± 3.9%. Genotoxic effects of permethrin (A), nanopesticide permethrin (B), nanopesticide CuSO4.5H2O (C), nanopesticide Cu(OH)2 (D), acephate (E), nanopesticide acephate (F), validamycin (G), validamycin nanopesticide ( H) and CaCO3 (I) in the comet test. Genotoxic effects of permethrin (A), nanopesticide permethrin (B), CuSO4·5H2O (C), nanopesticide Cu(OH)2 (D), acephate (E), nanopesticide acephate (F), validamycin (G), nanopesticide validamycin (H ) ) and CaCO3(I) in the comet assay.
Statistically significant decreases were observed in the mRNA expression of Duox and Hml in larvae exposed to and 5 mM concentrations of Cu(OH)2nano-pesticides, compared to the larvae of the control group. A significant increase in the expression of these genes was observed in larvae exposed to 1 and 5 mM concentrations of acephate and acephate nano-pesticides (Figure 12E,F). On the other hand, dose-dependent reductions were observed in the expression of Duox, Hml, Muc68D and PPO2 after exposure to acephate nano-pesticides and 5 mM).
Significant decreases were observed in Duox and Hml mRNA expression in larvae exposed to and doses of 5 mM acephate nanopesticides. A significant increase in the expression of these genes was observed in larvae exposed to concentrations of 1 and 2.5 mM validamycin and validamycin nanopesticides (Figure 12G, H). In addition, significant decreases in Muc68D gene expression occurred at the highest concentration of validamycin nanopesticides (2.5 mM).

Discussion
However, in larvae exposed to and 2.5 mM concentrations of validamycin, dose-dependent decreases in the mRNA expression of the Oggg1 gene were observed compared to larvae in the control group, and statistically significant results were found at 0.1, 1 and 2.5 mM doses of validamycin (Figure 13G). Meanwhile, after exposure to validamycin nanopesticides and 2.5 mM), the overall expression of Duox, Hml, Muc68D and PPO2 decreased in a concentration-dependent manner. Statistically significant decreases were observed in the mRNA expression of Duox and Hml in larvae exposed to and 5 mM concentrations of validamycin nanopesticides compared to control group larvae, while the expression of the PPO2 gene was statistically significant in larvae exposed to the two highest concentrations of validamycin nanopesticides (1 and 2.5 mM).
Compared to the control group, larvae exposed to 2.5 mM concentrations of validamycin and validamycin nanopesticides generally showed a decrease in mRNA expression of the Ogg1 gene, in a concentration-dependent manner, while finding statistically significant results at doses of 0.1, 1 and 2.5 mM (Figure 13G, H). No statistical differences were observed in the expression of Hsp70, Hsp83, CAT, SOD2, p53, Ogg1, Duox, Hml, Muc68D or PPO2 for the nanocapsules (2.5 mM CaCO3) used during the preparation of the validamycin nanopesticides. One of the first steps involved in the first response is related to the Hsp gene family.
A significant increase was observed in the expression of SOD2, p53, Hsp70, Hsp83 and CAT genes of larvae after exposure to permethrin (0.06 and 0.1 mM), permethrin nanopesticides (1 and 2.5 mM), acephate and acephate (1 nanopesticides). ), and validamycin and validamycin nanopesticides (1 and 2.5 mM). In addition, exposure to CuSO4·5H2O (1 and 5 mM) caused an increase in the expression of CAT and p53 genes, while exposure to the nanopesticide Cu(OH) 2 resulted in a significant decrease in the expression of Hsp70, Hsp83, CAT, SOD2 genes. and p53. While no statistical differences were observed for CaCO3 and validamycin in nanocapsules, significant decreases were detected after exposure to permethrin nanopesticides, Cu(OH) 2 nanopesticides, acephate nanopesticides, and validamycin nanopesticides.
A non-statistically significant increase in Ogg1 gene mRNA expression was observed in larvae exposed to 1 mM concentration. Nanocapsules used to prepare nanopesticides did not induce significant changes in the expression of Hsp70, Hsp83, CAT, SOD2, p53, Ogg1, Duox, Hml, Muc68D or PPO2. In this project, a statistically significant concentration-dependent decrease in Hsp70 and SOD2 gene expression was detected, while a significant increase in CAT and p53 gene expression was detected at and 5 mM CuSO4·5H2O concentrations, consistent with literature studies.
Dose-dependent decreases in the expression of Duox, Hml, Muc68D, PPO2 and upd3 - the genes associated with midgut-hemocyte interaction - were detected in Drosophila larvae. Although there were no changes in the expression of the Muc68Dgene, a significant decrease in the expression of Duox, Hml and Upd3 was observed. Furthermore, a statistically significant effect was only recorded in the expression of the PPO2 gene at the high concentration (10 mM).
Methods 1. Chemicals
A significant loss of crop yield can occur indirectly due to a decline in the pollinator population. The residual organic solvent in the mixture containing acephate encapsulated with PEG-400 was removed by means of a vacuum evaporator. Changes in the crystalline structure of CaCO3 particles were investigated by XRD measurements performed both during the synthesis stages and after validamycin loading.
The characterization of the nano-pesticides was obtained by analyzes TEM (LEO 906 E TEM by ZEISS) (Austin, Texas, USA), SEM (LEO 1430 by ZEISS) (Hillsboro, Oregon, USA), DLS (Worcestershire, UK) and laser Doppler ve-locimetry (LDV) (Malvern Zetasizer Nano-ZS ZEN 3600) (Worcestershire, UK). Measurements were performed by Cu Kα radiation at 2θ in the angular range of 10–90◦ and at a scan speed of 0.01◦. The permethrin content in the prepared permethrin nano-pesticide was characterized using a Shimadzu Prominence High-Performance Liquid Chromatography (HPLC) system with a UV detector at 225 nm and an Inertsil ODS-3 C18 column.
Endotoxin content was measured by chromogenic Limulus amebocyte lysate (LAL) assay (Lonza (QCL-1000TM), Inc., Walkersville, MD, USA), according to the protocol described in the manual and in previous studies. Three different mutant strains were used in the study: wild Canton-S, flare-3 and multiple wing hairs. Two different flight strains were used in the SMART wing test: flare-3 with genetic constitutionflr3/In (3LR) TM3,Bds and multiple wing hairs with mwh/mwh genetic constitution.
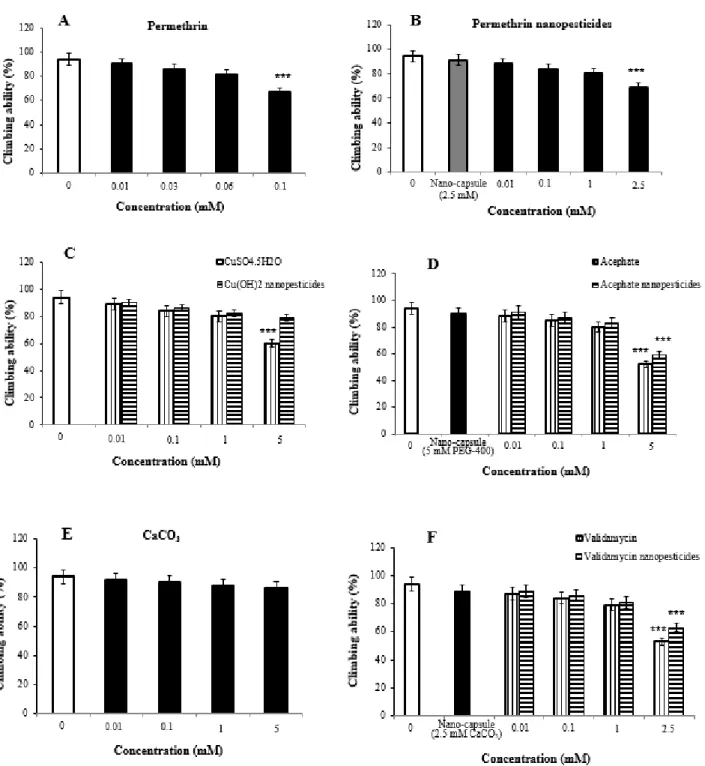
The F0 (parent), F1, F2 and F3 generation flies in the control groups, together with those in the study groups exposed to test chemicals, were carefully observed under a stereomicroscope to note any possible phenotypic variations in the head, thorax, eyes sign , mouth, wings, legs or abdominal region. In accordance with the procedures proposed in the previous work in the literature, the locomotor behavior of the flies was measured by climbing tests. The methods applied were mainly as follows: 40 larvae were used for each concentration in the control and study groups, and 5 replicate tests were performed in each concentration group.
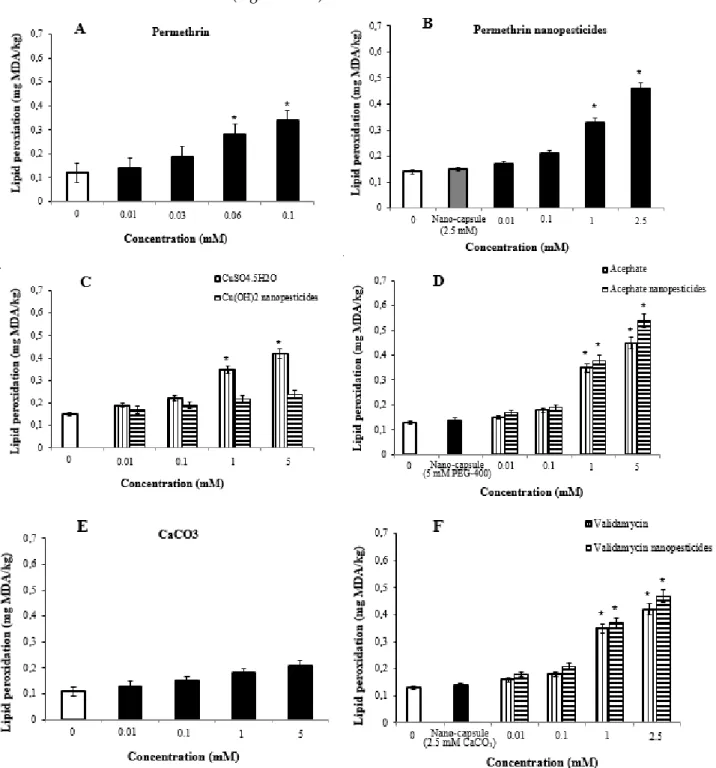
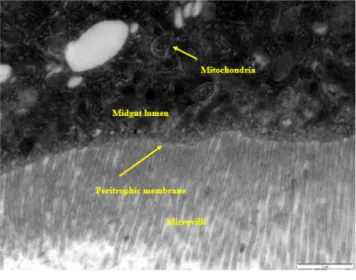
The lipid peroxidation tests were performed by measuring the amounts of malondialdehyde in the control and study groups. Third instar drosophila larvae (72 ± 4 hours) were exposed for approximately 24 hours to the compounds used as negative controls, nanopesticides, and various concentrations of microparticulates of the nanopesticides. This mixture was taken into a separate flask, rotated overnight at room temperature, and embedded in the mixture (1st araldite).
Conclusions
The effects of (nano)formulations on the fate of an insecticide in soil and implications for environmental exposure assessment.Environ. Detoxification and reproductive system-related gene expression following exposure to Cu(OH)2 nanopesticide in water fleas (Daphnia magna Straus 1820).Environ. Comparative cytotoxic and genotoxic effects of permethrin and its nanometric form on human erythrocytes and lymphocytes in vitro.Chem.
New insights into the toxic/genotoxic effects of CuO nanoparticles in the in vivo Drosophila model. Nanotoxicology. Influence of extracellular polymeric substances on the long-term fate, dissolution and speciation of copper-based nanoparticles.Environ. Demir, E.; Aksakal, S.; Turna, F.; Kaya, B.; Marcos, R. In vivo genotoxic effects of four different nano-sized forms of silica nanoparticles in Drosophila melanogaster.J.
Alaraby, M.; Demir, E.; Domenech, J.; Velázquez, A.; Hernández, A.; Marcos, R. In vivo evaluation of the toxic and genotoxic effects of exposure to cobalt nanoparticles in Drosophila melanogaster. Environment. Evaluation of the side effects of poly(epsilon-caprolactone) nanocapsules containing atrazine against maize plants.Front. On the safety of nanoformulations to non-target soil invertebrates - an atrazine case study. Environment.
Toxicity of engineered metal oxide nanomaterials mediated by nano-bio-ecological interactions: A review and perspective. The environment. Can atrazine-loaded nanocapsules reduce the toxic effects of this herbicide in the fish Prochilodus lineatus. Evaluation of interactive effects of UV light and nano-encapsulation on azoxystrobin toxicity in zebrafish. Nanotoxicology.
Hepatic effects of the clomazone herbicide in both its free form and associated with chitosan alginate nanoparticles in bullfrog tadpoles. Chemosphere. Evaluation of sublethal effects of nanoformulation of essential oils based on polymers on the German cockroach.Ecotoxicol. Oral administration of graphene oxide nanosheets induces oxidative stress, genotoxicity and behavioral teratogenicity in Drosophila melanogaster.Environ.