K jejich největší radiaci došlo krátce po masovém vymírání na přelomu křídy a paleogénu (Ksepka et al., 2020). Někteří autoři jsou dokonce toho názoru, že může existovat až 18 tisíc druhů, které lze popsat (Barrowclough et al., 2016). Přesto stále nepanoval konsenzus o homologii jednotlivých částí v ptačím a savčím mozku (přehled Jarvis et al., 2005).
Dále hvězdicové a košíčkové buňky v kůře mozečku (Weyer a Schilling, 2003) a některé neurony v suprachiasmatickém jádře (Morin et al., 2011). První práce zabývající se nesavčím obratlovcem, krokodýlem nilským (Crocodylus niloticus), byla publikována v roce 2016 (Ngwenya et al., 2016).
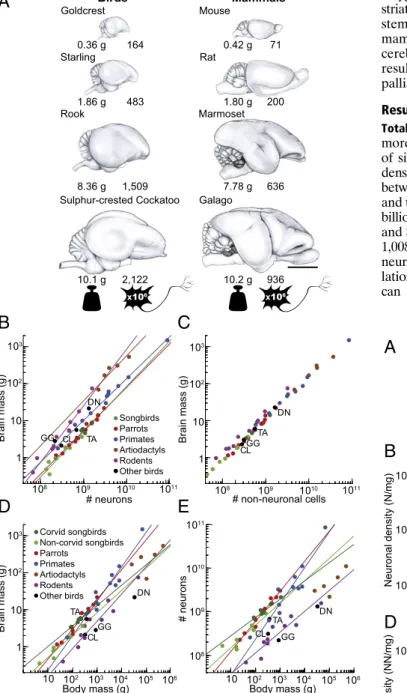
Birds have primate-like numbers of neurons in the forebrain
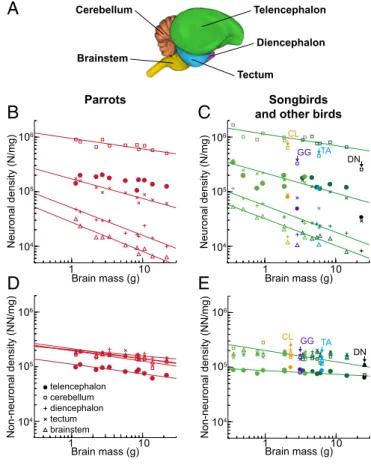
Thus, unlike mammals, the larger brain of songbirds and parrots contains increased neuron sizes in the telencephalon, and correspondingly decreased neuron sizes in the cerebellum and other brain regions (Fig. 4CandD). Neuronal densities in the avian pallium exceed those observed in the primate pallium by a factor of 3–4 (Fig. 3A). Note that the density of neurons is higher in parrots and songbirds than in mammals (for statistics, see SI Results). D–F) Average sizes of neurons included in the pallium (D), cerebellum (E), and rest of the brain (F).
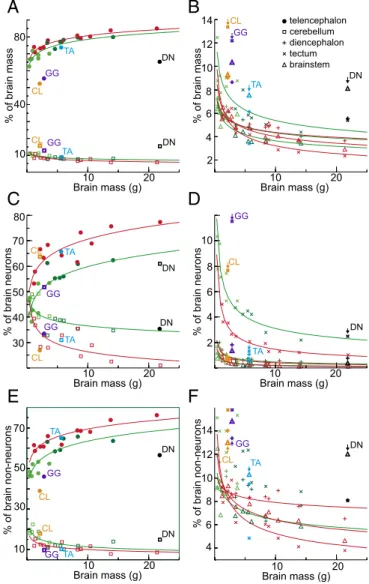
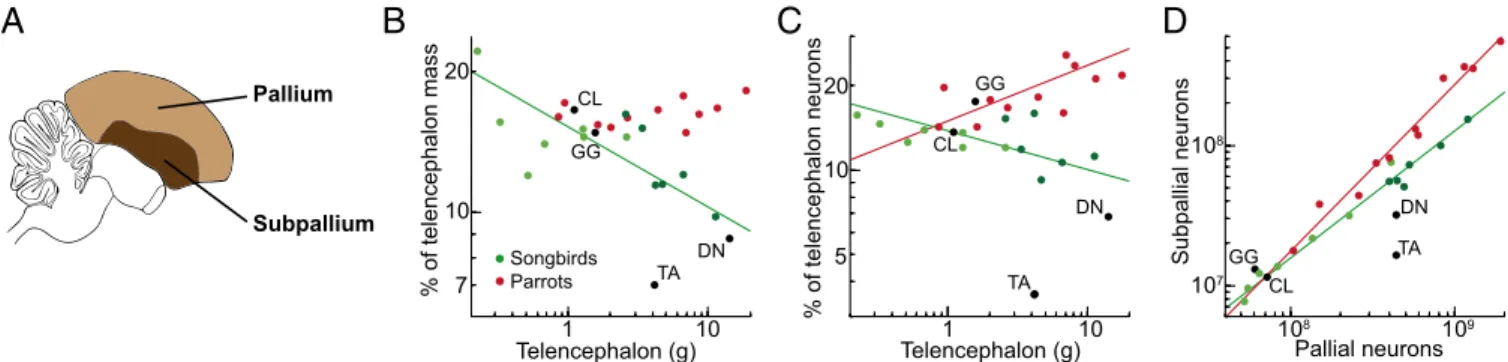
Note that the increase in the proportion of brain neurons in the rest of the brain in parrots can be attributed to a specifically increased number of neurons in the subpallium (Figure 5). Pie charts show the proportions of neurons in the pallium (green), cerebellum (red), and the rest of the brain (yellow).
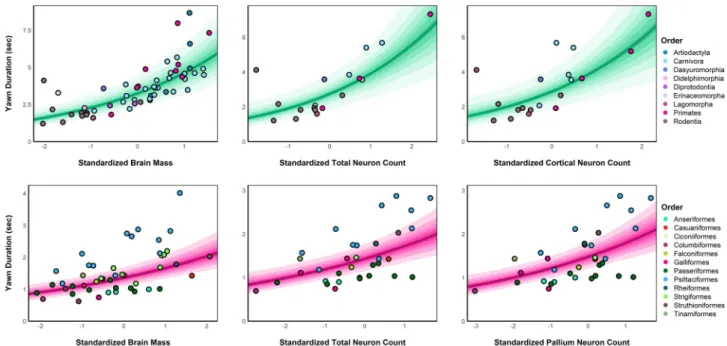
Brain size and neuron numbers drive differences in yawn duration across
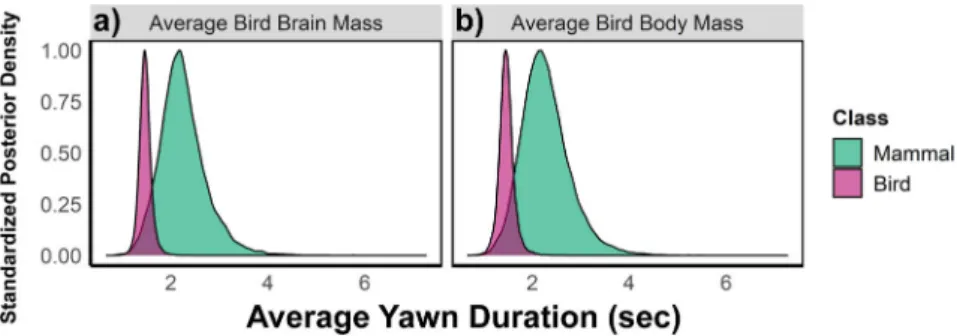
Phylogenetically controlled analyzes revealed robust positive correlations between yawn duration and (1) brain mass, (2) total neuron number, and (3) cortical/pallial neuron number in both mammals and birds, which cannot be solely attributed to allometric scaling rules. In support of this prediction, an initial study of 24 mammalian species found strong correlations between average yawn duration and the brain mass and number of cortical neurons for these species26. The aim of this study was to investigate how absolute brain mass and neuron number drive differences in yawn duration across mammals and birds, but models that do not control for body mass cannot rule out.
Consistent with the findings using more robust Bayesian methods, significant associations between residual yawn duration and all residual brain measures were observed in mammals and birds (see Supplementary Note 2). Our data therefore do not support a difference in the slopes of yawn duration on brain size or complexity between mammals and birds (see also Supplementary Fig. S1). Thus, despite common scaling patterns, our data suggest that the difference in mean yawn duration between mammals and birds cannot be attributed solely to allometric scaling with size.
Nevertheless, we still find clear effects of adjusted brain size on yawn duration in both mammals and birds, as well as for body size adjusted neural counts (both total and pallial) in birds, demonstrating the robustness of our findings. The overall pattern between yawn duration and neurological measures was robust across both vertebrate classes, as there was no clear difference in the slopes between mammals and birds (see Figure 3). Therefore, we first examined the general relationship between yawn duration and our brain measurements without correction for body size.
We used Bayesian multilevel phylogenetic models to examine the associations between yawn duration and the brain measurements, while taking into account the species' phylogenetic history. In addition, we estimated random slopes models to examine whether associations between yawn duration and the brain measures differ between mammalian and avian orders. In addition to estimating associations between yawn duration and neurological measures within our mammalian and avian samples, we also formally compared the discrimination of yawn duration and slopes of the brain size effects between these samples.

Neuron numbers link innovativeness with both absolute and relative brain size in
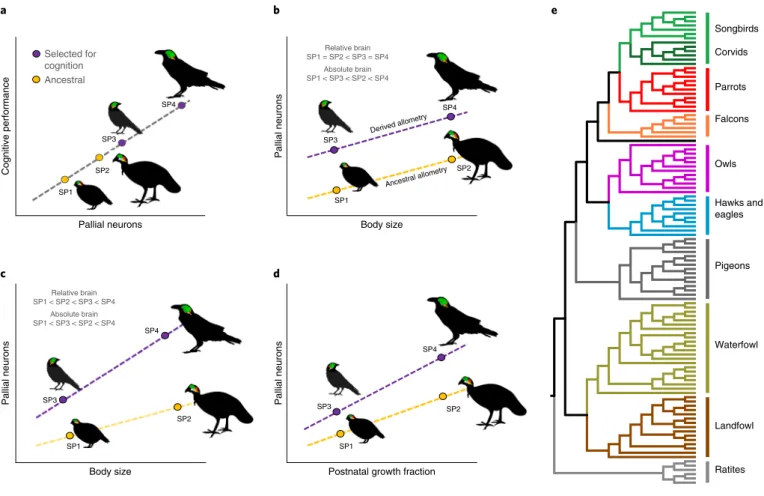
Encephalization - the evolutionary increase in the brain beyond what is expected for a given body size1 - has long been considered an important factor in the evolution of intelligence2,3. Based on neuron number estimates for 111 bird species, we show here that the number of neurons in the pallial telencephalon is positively associated with an important expression of intelligence: propensity for innovation. Finally, we test whether the accumulation of neurons in the pallium is related to an extension of later developmental stages.
Cognitive performance has long been assumed to depend on the number of neurons in the brain46,47, but this idea is currently supported by surprisingly little empirical evidence15. While the number of neurons in the whole brain and pallium increases similarly with both absolute and relative brain size (Fig. 4), the number of neurons in the cerebellum is more strongly related to absolute brain size alone; those in the brainstem follow no clear pattern. We found that longer developmental time leads to a greater accumulation of the number of neurons in the pallium of clades with high gradients than in those showing low gradients (Fig. 5 and Supplementary Fig. 8).
First, we provide firm support for the intuitively appealing idea that cognitive performance is limited by the number of neurons in the pallium and, to a lesser extent, the cerebellum. Second, we show that an increase in the number of neurons in the areas most closely involved in cognitive performance, the pallium, increases brain size, both in absolute and relative terms. The reason why the dual role of absolute and relative brain size in cognition has been undervalued in the past probably reflects the common practice of removing the allometric effects of body size.
Our study is based on an updated database that quantifies neuron numbers in the whole brain and three brain areas—the pallium (which includes the hyperpallium, mesopallium, nidopallium, entopallium, acropallium, and hippocampus), the cerebellum, and the brainstem (which includes the medulla oblongata and involving midbrain). tegmentum) for bird species. To be included in the RF together with absolute brain size, we estimated relative brain size by means of the normalized scaled brain index73 (NSBI). Isotropic fractionator: a simple, rapid method for quantifying total cell and neuron numbers in the brain.
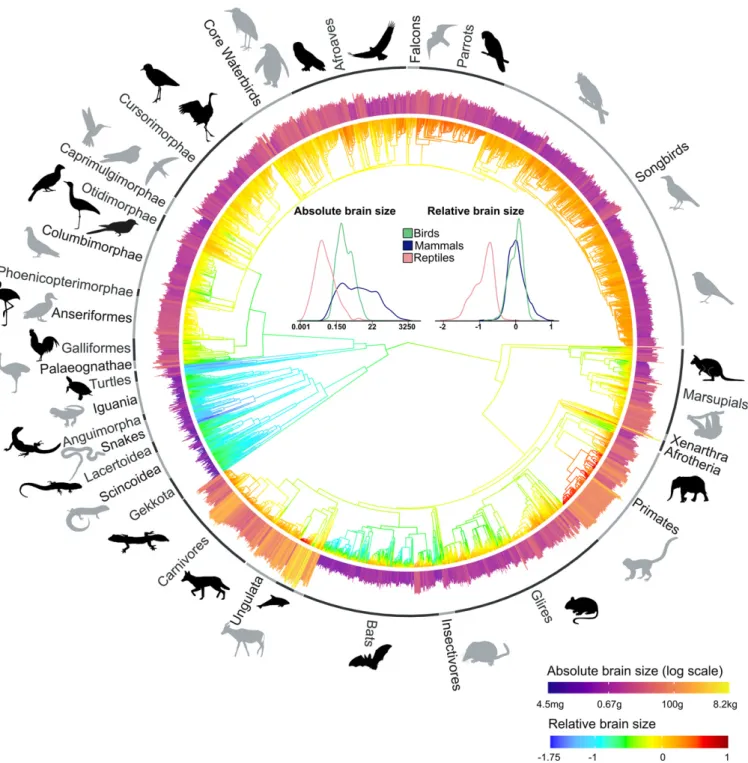
The evolution of brain neuron numbers in amniotes
The other two major increases in the number of neurons occurred in land birds and anthropoid primates, two groups known for their cognitive prowess. This capacity is better determined by the number of neurons in the brain or specific brain regions (although their relative importance is still debated), their connections, interneuronal distance, and axonal conduction velocity (8, 9). Not only can brains of the same size vary in the number of neurons, but also the total number of neurons can be allocated to different brain parts.
Similar changes were uncovered for the scaling of neurons with body mass ( SI Appendix , Fig. S7 and Table S3 ), where an additional decrease in the number of cerebellar neurons was found in snakes. The resulting changes in the number of neurons for body mass follow different pathways in different brain parts ( Fig. 4 and SI Appendix , Fig. S8 ). The gray scale bars indicate the proportion of brain neurons found in the telencephalon, rest of the brain and cerebellum.
These effects are then compounded in taxa that exhibit a positive relationship between relative brain size and relative neuron density, leading to a disproportionately higher number of neurons in species with relatively large brains. We suggest that these convergent increases in neuron number represent stepping stones in the evolution of avian and mammalian intelligence. Perfusions. Animals were killed by anesthetic overdose (intramuscular administration of ketamine and xylazine for reptiles; inhalation of halothane for birds, except for ostrich, rhea and emu, which were overdosed by intramuscular injection of anesthetics containing midazolam, detomidine, butordeamine, ketamine, butordeamine ).
The rest of the brain refers to the rest after the separation of the telencephalon and cerebellum, that is, the diencephalon, mesencephalon and medulla oblongata. Starting with the full model, the putative degrees were successively aggregated with the ancestral degree to confirm that they were significantly different, and the identified unrelated degrees were then aggregated to identify convergence. The models were compared with the kernel R function ANOVA, and the simplified model (with) was adopted for excess value 0 .05. Evolution rates. To compare the evolutionary rates between the different brain parts ( SI Appendix , Table S5 ), we used an approach specifically devised to compare evolutionary rates for multiple phenotypic traits on a phylogeny ( 80 ). a model where all traits evolve at a common rate.
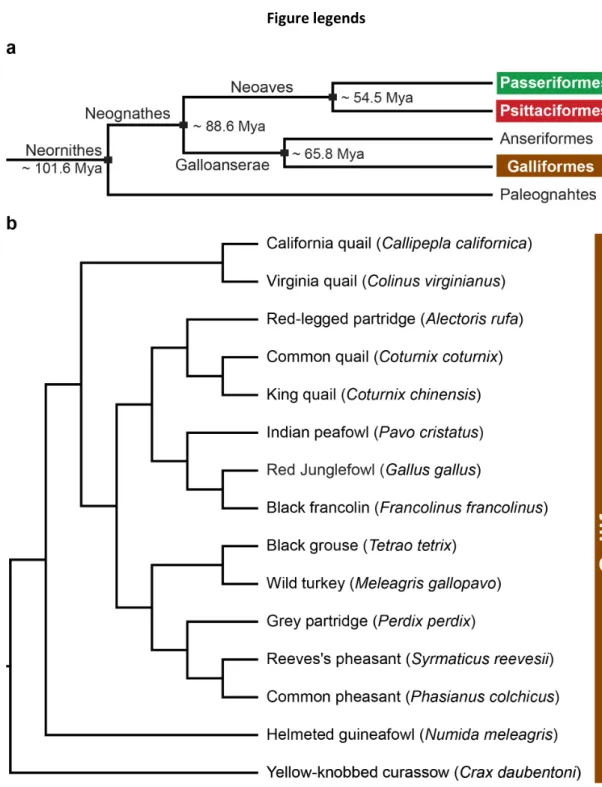
Cellular scaling rules for brains of the galliform birds (Aves, Galliformes)
Recently, corvid and parrot brains have been shown to contain vast numbers of neurons packed in high densities in the telencephalic pallium (Olkowicz et al. 2016), particularly in the associative regions (Ströckens et al. 2022). In addition, the numbers of pallial neurons scale with body size at a steeper slope compared to other avian clades (Sol et al. 2022). Brains were dissected into different components using the Olympus SZX 16 stereomicroscope as previously described (Olkowicz et al., 2016).
Similar to mammals (Herculano-Houzel et al. 2015), in galliform birds the cerebellum contains the majority of cerebellar neurons, and the telencephalon typically contains less than 35% of cerebellar neurons. The opposite pattern was observed in songbirds and parrots (Olkowicz et al. 2016; Kverková et al. 2022). They have significantly fewer neurons in the associative parts, especially in the nidopallium and mesopallium (Ströckens et al. 2022).
Ve skutečnosti se ukázalo, že velikost asociačních oblastí (Lefebvre et al., 1997; Timmermans et al., 2000) a počet neuronů v telencephalonu (Sol et al. 2022) jsou dobrými prediktory sklonu k inovaci v ptáci.. Dále se zdá, že individuální variabilita v počtu neuronů v mozečku je vyšší než v jiných částech mozku (Herculano-Houzel et al. 2015). Tato data jsme následně porovnali s již publikovanými daty savců získanými stejnou metodou (Herculano-Houzel et al., 2015a).
89 množství zbytkového tepla, i když ptáci se srovnatelně velkým mozkem mají v palliu více neuronů (Olkowicz et al., 2016). Analýza počtu neuronů u nově vylíhnutých kuřat byla následně použita k ověření metody parciální izotropní homogenizace (Ngwenya et al., 2017). Podařilo se nám prokázat asociaci inovace jak s počtem neuronů v celém mozku, tak v palliu a mozečku (Sol et al., 2022).
Podle některých studií mají ryby žijící v teplé vodě větší relativní mozkovou hmotu (Yu et al., 2014). Svou roli by mohl hrát zobák, o jehož termoregulační funkci se dlouho spekulovalo a nedávno zdokumentovala (Tattersall et al., 2016).