In particular, I would like to express my gratitude to Pavlína Daňková, Ph.D., and Martin Rossmeisl, MD, Ph.D. Many other heartfelt thanks go to my partner Marek, who 'came' into my life just when I was about to complete my Ph.D. to complete, but was all the more supportive and helped me complete my studies.
W HITE AT
- D ISTRIBUTION AND F UNCTION
- H ETEROGENEITY
- Anatomical and functional heterogeneity
- Cellular heterogeneity
- W HITE ADIPOCYTES
- Heterogeneity and plasticity
- Endocrine properties
- L IPID T RANSPORT , S TORAGE , AND U TILIZATION
- Lipogenesis
- Lipolysis
Even the SAT located in the upper limbs (hands) has not been systematically investigated and only individual studies dedicated to this depot are available [44-46]. 24 diacylglycerol O-acyltransferase (DGAT) 2, sterol regulatory element-binding transcription factor (SREBF) 1 genes involved in lipid biosynthesis.

L YMPHATIC VASCULAR FUNCTIONS
The volume of ISF is five times greater than the plasma in blood capillaries, and therefore ISF represents approximately 15% of total body weight [140]. Moreover, thanks to new discoveries, it is clear that the LS is essential for the circulation of cerebrospinal fluid and aqueous humor, response to cancer treatments, liver function or the prevention of neurodegenerative and neuroinflammatory diseases, which further increases our understanding of the importance of proper LS functioning.
L YMPHATIC VASCULAR STRUCTURE
L YMPHATIC ENDOTHELIAL CELLS
H ETEROGENEITY AND P LASTICITY
Together we can hypothesize that the plasticity of LECs is driven by a response to a change in the different types of microenvironments. The remarkable heterogeneity and plasticity of LECs has been a challenge for researchers before the discovery of reliable lymphatic markers, which are described in the next chapter.
M ARKERS OF LYMPHATIC ENDOTHELIAL CELLS
Although PDPN is also a gene known primarily as a podocyte marker, it has become very useful for the detection of LECs [177]. LEC identity may also be characteristic for the expression of specific VEGF receptors, which are described in detail in the chapter below.
L YMPHANGIOGENESIS
VEGF FACTORS
During early development, VEGFR-3 is expressed in all endothelia but is later restricted to LECs and certain vascular ECs [186–188]. VEGF-A is considered the strongest pro-angiogenic activator involved in angiogenesis under physiological and pathological conditions [ 191 , 192 ].
VEGF-C
- Biosynthesis and activation
- Molecular regulation: VEGF-C/VEGFR-3 signaling
The minor form is likely a plasmin cleavage product that is approximately nine amino acids longer compared to the major form of VEGF-C [186]. When AKT is phosphorylated, activation of mammalian targets of rapamycin and Rac 1 is achieved [1].
![Figure 6: Schematic illustration of biological activation of VEGF-C. Adapted from [186]](https://thumb-eu.123doks.com/thumbv2/9pdfinfo/2882272.438435/40.892.208.746.306.651/figure-schematic-illustration-biological-activation-vegf-c-adapted.webp)
As discussed above, both AT and LS have an impact on metabolic maintenance through the regulation of lipid homeostasis [211]. However, due to extremely evolving molecular and genetic approaches and modern imaging technologies, the fascinating anatomical, cellular and molecular interplay between the structure and components of AT and LS is slowly being revealed.
L YMPHATIC DISTRIBUTION IN AT
A cross between LS and AT was neglected, as there were almost no means to visualize LS in vivo and to manipulate experimental lymphatic drainage [220]. The known facts about the interaction between LVs, LNs, LS components (especially ICs) and AT are described in the next chapter.
A DIPOSE - LYMPHATIC FUNCTIONS
It was shown that the abnormal accumulation of AT in the mesentery observed in Crohn's disease was associated with defects in intestinal lymphatic drainage [235, 236]. Therefore, we can suggest that leaky lymph cells in the intestinal area can contribute to the accumulation of VAT.
A DIPOSE - LYMPHATIC DYSFUNCTIONS
O BESITY
Therefore, one of the aims of my thesis was to elucidate the role of different degrees of lymphatic drainage on the lipolytic activity of femoral SAT in women. These facts may support the idea that accumulation of VAT may be linked to subtle nuances of intestinal lymphatic drainage efficiency.
L YMPHEDEMA
Despite significant adjustments in surgical strategies and modern molecular and genetic therapy options, complete recovery from secondary lymphedema is still unattainable [255]. Causality between defects of LS and accumulation of AT has been described in mouse models of primary and secondary lymphedema.
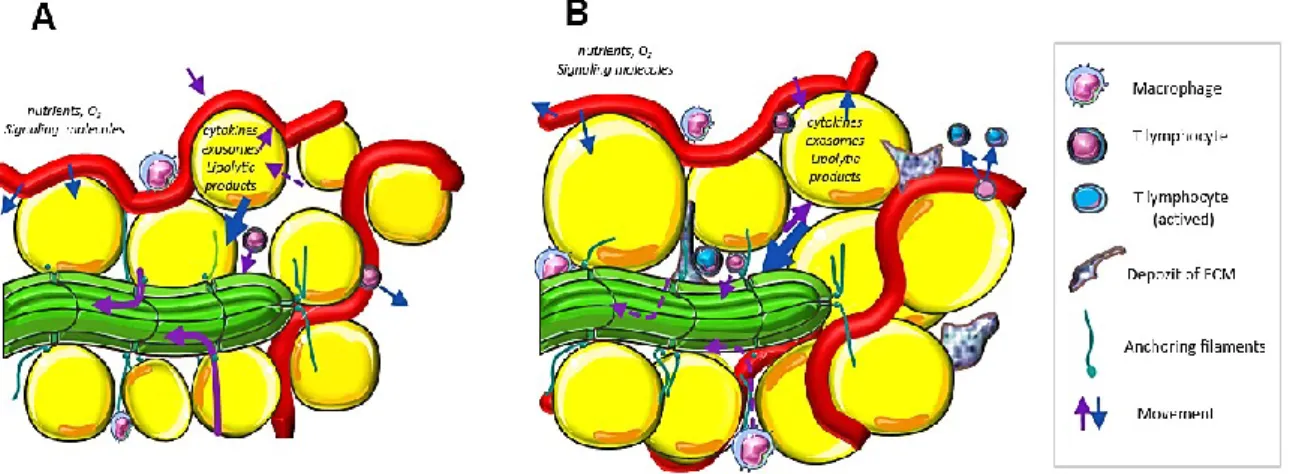
P ROADIPOGENIC - LYMPHATIC STIMULI
Therefore, the researchers hypothesized that lymph/ISF might contain proadipogenic stimuli, which is described in the next chapter. The impairment of lymphatic drainage (B) is then associated with an abnormal accumulation of ICs, manifestations of sterile inflammation and fibrosis.
C ELLULAR CROSSTALK OF ADIPOCYTES AND LEC S
Nevertheless, clear evidence of this type of lipid transport between LECs and adipocytes is still lacking. 284] showed in a thromboxane-deficient (Tp-/-) mouse model that lipopolysaccharide-induced lymphangiogenesis was inhibited, leading to impaired lymphatic drainage.

W OMEN WITH NORMAL AND WORSENED LYMPHATIC DRAINAGE (A IM 1)
H EALTHY WOMEN AND BREAST CANCER SURVIVORS WITH / WITHOUT NON - PITTING
B ODY FLUID SAMPLE
ISF COLLECTION
- Blister ISF collection
- Microdialyzate collection
LAT FLUID COLLECTION
B ODY FLUID ANALYSES
A DIPOSE TISSUE
L YMPHEDEMA AT
H EALTHY AT
AT PROCEDURES
C OLLAGENASE DIGESTION OF AT
AT EXPLANTS
I N VITRO CELL CULTURE
E STABLISHMENT OF AN IN VITRO CULTURE OF PREADIPOCYTES
C ONVERSION OF PREADIPOCYTES INTO ADIPOCYTES
H UMAN D ERMAL L YMPHATIC E NDOTHELIAL C ELLS
I N VITRO EXPERIMENTS
E FFECT OF BODY FLUIDS ON ADIPOGENESIS
S INGLE AND COCULTURE MODELS
- Single culture: Application of ACM on HDLECs
- Single culture: Application of LCM on white adipocytes
- Indirect coculture system
- Direct coculture system
Collection of Fat Conditioned Medium (CM) - 12 day differentiated adipocytes were washed twice with DPBS +Mg 2+ and Ca 2+ and insulin starved in EBM-2 medium supplemented with transferrin (0.1 ug/ml) for 16-19 h. The medium was aspirated and cells were exposed to EBM2 (supplemented with transferrin (0.1 µg/ml), 0.5% free FFA and endotoxin low BSA) with or without 1 mM dibutyryl-cAMP (lipolysis inducer) for 30 min . Then the medium was discarded and the cells were washed twice with DPBS +Mg 2+ and Ca 2+ .
Then the medium was aspirated and the cells were exposed to LCM (diluted with EBM-2 medium at a ratio of 1:1) for 42 h. After exposure to LCM and control media, cells were washed once with DPBS +Mg 2+ and Ca 2+ and used for a standard lipolytic assay described in 5.8. To ensure proper media adaptation, fully differentiated adipocytes (standard culture, male donors) growing on coverslips were washed twice with DPBS + Mg 2+ and Ca 2+ and then fed with EBM-2 medium supplemented with transferrin (0.1 ug/ml).
L IPOLYSIS ASSAYS
Subsequently, the medium was aspirated and HDLECs were resuspended in EGM-2 medium with 2% fetal bovine serum added at a density of 2.5 x 104 cells/cm2 directly on adipocytes to generate a direct co-culture system. The cell density was chosen according to our experimental culture to ensure sufficient cell growth. As controls, monocultures of adipocytes and HDLECs were maintained in the same manner as the cells in co-culture. The co-culture was washed twice with DPBS + Mg2+ and Ca2+ and refed with FBS-free EGM-2 medium with or without VEGF-C (50 ng/ml) after 24 hours of incubation, which was the time required for HDLEC attachment.
FACS
E VALUATION OF ADIPOCYTE SIZE
WST-1 ASSAY
I NDIRECT IF
C ONFOCAL LASER SCANNING MICROSCOPY
With Z-stack settings ranging from 0.1 to 1 m, an appropriate number of optical slices was determined for each field of view.
I MAGE P ROCESSING
A NALYSIS OF LYMPHANGIOGENIC LOOPS
C LUSTER A NALYSIS
C ELL - COVERED AREA
G ENE EXPRESSION ANALYSIS
RNA ISOLATION
R EVERSE TRANSCRIPTION (RT)- Q PCR
RT- Q PCR WITHOUT RNA ISOLATION
MI RNA ANALYSIS
- H AEMOLYSIS
- MI RNA ISOLATION
- RT- Q PCR – IDENTIFICATION OF MI RNA
- A NALYSIS OF RESULTS
- P URIFICATION OF PRO -VEGF-C
- T HERMOLYSIN ACTIVATION ASSAY
- B A /F3 VIABILITY ASSAY
LNA (lock nucleic acids) technology improves binding affinity with high sensitivity and specificity to low expression miRNAs with improved mismatch discrimination. Reverse transcription was performed according to the manufacturer's instructions with minor modifications. Accurate prediction of miRNA targets is the next important step in the interpretation of miRNA analysis results. Today, several online databases, which are freely available, represent valuable tools to identify potential miRNA target genes.
The target genes of our selected miRNAs were predicted using the free online miRBD database (http://mirdb.org/). First, we select target genes with a planned target score of 50 for each miRNA with altered expression. After incubation, the biological activity of the treated pro-VEGF-C was assessed by the Ba/F3 viability assay.
S TATISTICAL A NALYSIS
The cells were washed three times with diluted PBS and seeded at a density of 15-10,103 cells/well in 96-well plate. Cells were incubated for 48 h, and then 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide) solution (0.25 mg/ml) was added to each well for 2 h incubation. After incubation, a lysis solution (stop solution) (10% SDS, 10 mM HCL) was added for 5–10 min, and absorbance at 540 nm was measured to reflect cell metabolic activity.
T ABLES OF S UPPLEMENTS
Compared to the NLD group, plasma glycerol levels were lower in the WLD group, the same trend (p = 0.07) was observed for FFA (Figure 16A). 85 lower, while lipid droplet coat PLIN1 expression was higher in the diseased limb compared to the healthy and healthy LYM limb (Figure 19 B). Interestingly, there was higher mRNA expression of HAND2, PDPN, and PECAM in the intact LAT, but lower expression of the negative regulator of angiogenesis, NDN, compared to AT from the healthy limb of LYM and healthy subjects (Figure 22C).
In preadipocytes, we observed lower expression of INHBA, VEGFA and GDF15 in the presence of LAT fluid (Fig. 23A). Nevertheless, mRNA levels of all target genes (ZHX1, CPEB3 and AGO1) did not show significant up/down regulation in the diseased AT limb compared to the healthy limb (Fig. 25). However, we detected higher basal levels of FFA in medium from adipocytes pretreated by LCM (Fig. 28B).
These changes in adipocytes were not seen in the absence of VEGF-C, as demonstrated by adipocyte area coverage analysis (Figure 32B). Surprisingly, basal lipolysis was dramatically increased in co-culture independent of VEGF-C addition after 2 days (Figure 36).

D ISCUSSION I
Interestingly, a low ability to stimulate lipolysis in subcutaneous abdominal AT has been suggested as a predictor of long-term weight gain in women [305]. Taken together, these changes could contribute to GAT resistance to lipolytic stimuli in WLD women. However, the exact mechanism how WLD might regulate GAT's response to lipolytic stimuli remains to be determined.
More specifically, anti-inflammatory Th2 CD4+T lymphocytes are the main players in the tissue changes induced by lymphostasis [306]. In conclusion, the main result of this study is the recognition of the possible impact of lymphatic drainage on lipolysis in the femoral AT and on the distribution of AT metabolites between the AT interstitium and circulation. Thus, the function of the lymphatic system should be taken into account in studies related to AT physiology and especially in interventions aimed at reducing AT mass.
D ISCUSSION II
Accordingly, previous mRNA profiling in AT by microarrays did not detect any difference in the expression of typical lipogenesis or adipogenesis markers in healthy compared to diseased limb of lymphedema [309]. Thus, posttranscriptional regulation is likely more involved in maintaining the diseased phenotype of lymphedema. In fact, KLF4 and KLF6, whose expression was correlated, are transcription factors with the mixed role in the regulation of adipogenesis [310].
PLIN1 and PLIN3 expression was increased in the LAT, along with a modest increase in mRNA levels of HSL, another major AT lipase. Interestingly, TNC was found to be elevated in visceral (but not subcutaneous) AT in obese humans [314], i.e., in the AT depot with an intimate connection to intestinal lymphatics. Murine model of lymphedema revealed a substantial increase in CD4+ T lymphocytes and macrophages after LS surgical injury [271].
D ISCUSSION III
D ISCUSSION IV

![Figure 7: Schematic illustration of the VEGF-C/VEGFR-3 signaling pathway. Adapted from [1] and later modified](https://thumb-eu.123doks.com/thumbv2/9pdfinfo/2882272.438435/41.892.147.542.589.1065/figure-schematic-illustration-vegfr-signaling-pathway-adapted-modified.webp)