MARIA APARECIDA FERNANDES
PADRÕES TEMPORAIS E GRAU DE DIVERSIFICAÇÃO CARIOTÍPICA
EM ESPÉCIES ATLÂNTICAS DA FAMÍLIA ACANTHURIDAE
(PERCIFORMES)
Orientador: Prof. Dr. Wagner Franco Molina
NATAL-RN 2015
Catalogação da Publicação na Fonte. UFRN / Biblioteca Setorial do Centro de Biociências
Fernandes, Maria Aparecida.
Padrões temporais e grau de diversificação cariotípica em espécies Atlânticas da família Acanthuridae (Perciformes) / Maria Aparecida Fernandes. – Natal, RN, 2015.
54 f.: il.
Orientador: Prof. Dr. Wagner Franco Molina.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Programa de Pós-Graduação em Sistemática e Evolução.
1. Evolução cromossômica. – Dissertação. 2. Fusões cêntricas. –
Dissertação. 3. Acanthuridae. – Dissertação. I. Molina, Wagner Franco II. Universidade Federal do Rio Grande do Norte. III. Título.
2
MARIA APARECIDA FERNANDES
PADRÕES TEMPORAIS E GRAU DE DIVERSIFICAÇÃO CARIOTÍPICA
EM ESPÉCIES ATLÂNTICAS DA FAMÍLIA ACANTHURIDAE
(PERCIFORMES)
Área de concentração: Sistemática e Evolução
Aprovada em 26/03/2015
BANCA EXAMINADORA:
__________________________________________________________ Dr. Wagner Franco Molina
Universidade Federal do Rio Grande Do Norte (Orientador)
___________________________________________________________ Dra. Sathyabama Chellappa
Universidade Federal do Rio Grande do Norte
___________________________________________________________ Dr. Roberto Ferreira Artoni
Universidade Estadual de Ponta Grossa
3 DEDICATÓRIA
A meus pais Vilmar (In memoriam) e Inácia,
AGRADECIMENTOS
Gostaria de agradecer especialmente aos meus pais, por todo esforço empenhado em minha educação. As minhas irmãs (Josivânia e Josenilda) e irmãos (Adriano, Mariano e João Paulo) pela maravilhosa convivência familiar, pelo carinho demonstrado e principalmente pela compreensão nos momentos difíceis!
Ao meu esposo Leandro pelo companheirismo e por contribuir sempre de maneira incondicional para o meu crescimento pessoal e profissional.
Ao professor Dr. Wagner Franco Molina, pelo apoio, orientação e confiança depositados durante a realização deste trabalho.
Aos companheiros de luta do LGRM: Gideão, Karlla, Rafael, Rodrigo, Amanda, Paulo, Clóvis, Juliana, Inailson, Allyson, Josi, Roberta, Calado e Cris. Obrigada a todos pelas significativas contribuições seja nos procedimentos laboratoriais e/ou em campo, e pelos momentos de descontração durante nossos cafezinhos.
Registro também meu agradecimento à minha querida professora Drª. Simone Almeida por todo apoio, disponibilidade e conselhos (acadêmicos e pessoais) desde os primeiros momentos da minha vida acadêmica.
Agradeço também as professoras Drª Danielle Pereti, Danielly Alves e Drª Maisa Clari pelo apoio durante a graduação e a transição para o mestrado.
À CAPES pela concessão da bolsa de estudos.
À UFRN e ao Programa de Pós-Graduação em Sistemática e Evolução por me permitir concluir mais esta estapa de meu processo educacional.
SUMÁRIO
1. INTRODUÇÃO
1.1. Família Acanthuridae 11
1.2. Filogenia e evolução da família Acanthuridae 12 1.3. Aspectos biológicos e taxonômicos do gênero Acanthurus 13
1.4. Citogenética como ferramenta para inferências evolutivas 16
2. OBJETIVOS
2.1. Objetivo geral 19
2.2. Objetivos específicos 19
3. MATERIAL E MÉTODOS
3.1. Material 20
3.2. Métodos 20
3.2.1. Obtenção de cromossomos mitóticos 20 3.2.2. Análises cromossômicas e montagem do cariótipo 20 3.2.3. Detecção de regiões organizadoras de nucléolo 21 3.2.4. Detecção de heterocromatina constitutiva 21 3.2.5. Dupla coloração com fluorocromos base-específicos 21 3.2.6. Hibridação in situ fluorescente FISH 22
3.2.6.1. Obtenção de sondas para hibridação 22 3.2.6.2. Hibridação cromossômica
22
4. CAPÍTULO I - Sequential Steps of Chromosomal Differentiation in
Atlantic Surgeonfishes: Evolutionary Inferences
24
5. CAPÍTULO II - Atlantic Surgeonfishes Bear Only Minor
Microstructural Changes in Highly Derived Karyotypes
37
6. CONCLUSÕES 48
LISTA DE FIGURAS
Parte introdutória
Figura 1. (a) Hipótese filogenética para a família Acanthuridae, modificado de Sorenson et al. (2013). (b) Relações entre espécies do gênero Acanthurus presentes no Atlântico, proposta
por Bernal & Rocha (2011).
13
Figura 2. Espécime de Acanthurus coeruleus. Em destaque indivíduo na
fase juvenil. Barra: 2 cm.
15
Figura 3. Espécime de Acanthurus chirurgus. Barra: 2 cm. 15
Figura 4. Espécime de Acanthurus bahianus. Barra: 2 cm. 16
Capítulo 1
Figura 1. Mapa da América do Sul mostrando os pontos de coleta de
Acanthurus coeruleus (a), A. bahianus (b), e A. chirurgus (c) no
estado do Rio Grande do Norte (1) e Bahia (2), nordeste do Brasil.
26
Figura 2.
Figura 3.
Cariótipos de Acanthurus coeruleus ((a) e (b)) com 2� = 48, A. bahianus ((c) e (d)) com 2� = 36, e A. chirurgus ((e) e (f) ) com
2� = 34, após coloração convencional com Giemsa ((a), (c), e (e)) e badamento-C ((b), (d) e (f)). Os cromossomos portadores de RONs, de cada espécie, após tratamento com nitrato de prata são apresentados em caixas (par 2 de A. coeruleus e par de 8 de A. bahianus e A. chirurgus).
Idiogramas de conjuntos cromossômicos de Acanthurus coeruleus (a), A. bahianus (b), e A. chirurgus (c) mostrando as
características citogenéticas mais conspícuas divididas em caixas. Em (d), o cariótipo basal dos Perciformes (2� = 48a) e uma hipótese filogenética com base em rearranjos
28
7 cromossômicos sequênciais inferidos nas três espécies
Acanthurus.
Capítulo 2
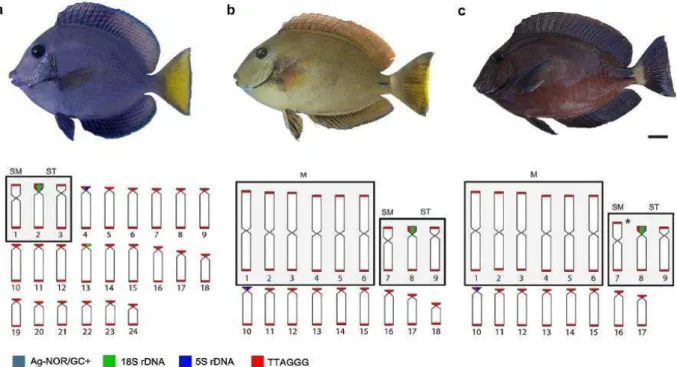
Figura 1. Cariótipo e metáfases de A. coeruleus (A), A. bahianus (B) e A. chirurgus (C) após a coloração Giemsa (esquerda), two-color
FISH (centro) com DNAr 18S (vermelho) e 5S DNAr (sinais verdes) distribuição de sondas de (TTAGGG)n sequências (direita). Caixa, os cromossomos portadores de RONs são mostrados após a coloração de nitrato de prata (esquerda) e coloração com fluorocromos (centro). Barra = 5m.
40
Figura 2. Idiograma de A. coeruleus (A), A. bahianus (B) e A. chirurgus
(C), combinando os resultados de NOR / localização de região rica em GC e mapeamento físico de ribossomal e sequências teloméricas. Os cromossomos homeólogos derivados de inversões pericêntricas (cromossomos sm-st) e fusões cêntricas (m-cromossomos) são destacados nas caixas. O asterisco no par 7 de A. chirurgus indica uma suposta autapomorfia (fusão em
tandem) para esta espécie.
LISTA DE ABREVIATURAS
2n – Número diploide a – Acrocêntrico
AFLP – Amplified fragment lenght polymorphism AgNO3 – Nitrato de prata
Ag-RONs – Regiões organizadoras de nucléolo evidenciadas pela impregnação com nitrato de prata
AT – Adenina e Timina
Ba(OH)28H2O – Hidróxido de bário BC – Bandamento C
CMA3 – Cromomicina A3
DAPI –4`4’,6-diamidino-2-fenilindol DNA – Ácido desoxirribonucleico
DNAr/rDNA – Ácido desoxirribonucleico ribossômico FISH – Fluorescence in situ hybridization
GC – Guanina e Citosina HCl – Ácido clorídrico KCl – Cloreto de potássio
LGRM – Laboratório de Genética de Recursos Marinhos m – Metacêntrico
Ma – Milhões de anos MM - Mitramicina n – Número amostral NF – Número fundamental RN – Rio Grande do Norte
RNAr/rRNA – Ácido ribonucleico ribossômico RONs – Regiões organizadoras de nucléolo sm – Submetacêntrico
st – Subtelocêntrico
9 RESUMO
A família Acanthuridae é um grupo bastante representativo dentre os peixes marinhos e que desempenha um papel fundamental na dinâmica ecológica dos recifes de corais. Três espécies pertencentes ao gênero Acanthurus são comuns ao longo dos recifes costeiros do Atlântico
Ocidental: A. coeruleus, A. bahianus e A. chirurgus. No presente estudo, são apresentados
dados citogenéticos para estas três espécies de Acanthurus com base em métodos
citogenéticos clássicos e no mapeamento de sequências ribossomais repetitivas, como DNAr 18S e 5S, além de sondas teloméricas com a finalidade de auxiliar na compreensão da carioevolução deste grupo. O padrão citogenético dessas espécies indica que as etapas sequenciais de rearranjos cromossômicos, que datam 19-5 milhões de anos atrás (Ma), são responsáveis por suas diferenças interespecíficas. Acanthurus coeruleus (2n=48; 2sm + 4st +
42a), A. bahianus (2n=36; 12m + 2sm + 4st + 18a) e A. chirurgus (2n=34; 12m + 2sm + 4st +
16a) compartilham um antigo conjunto de três pares cromossômicos originados através de inversões pericêntricas. Um conjunto de seis grandes pares metacêntricos formados por translocações Robertsonianas (Rb) encontrado em A. bahianus e A. chirurgus e uma suposta
fusão em tandem presente em A. chirurgus são eventos mais recentes. A falta de sequências
teloméricas intersticiais (ITS), apesar de várias fusões cêntricas em A. bahianus e A. chirurgus pode estar relacionada com o longo período de tempo após a sua ocorrência
(estimado em 5 Ma). Além disso, as homeologias entre os pares de cromossomos que carregam os genes ribossomais, além de outras características estruturais, destacam grandes regiões cromossômicas conservadas nas três espécies. Nossos resultados indicam que as mudanças macroestruturais ocorreram durante a cladogênese dessas espécies não foram seguidas por rearranjos microestruturais visíveis nos cariótipos.
ABSTRACT
The Acanthuridae family is a representative group from the marine fish that plays a key role in ecological dynamics of coral reefs. Three species are common along coastal reefs of Western Atlantic: Acanthurus coeruleus, Acanthurus bahianus and Acanthurus chirurgus. In
the present study, cytogenetic data are presented for these three species Acanthurus based on
classical cytogenetic methods and mapping of repetitive sequences such as ribosomal 18S and 5S rDNA and telomeric repeats to improve their karyotype evolutionary analyses. The cytogenetic pattern of these species indicated sequential steps of chromosomal rearrangements dating back 19 to 5 millions of years ago (M.a.) that accounted for their interspecific differences. A. coeruleus (2n=48; 2sm+4st+42a), A. bahianus (2n=36;
12m+2sm+4st+18a) and A. chirurgus (2n=34; 12m+2sm+4st+16a) share an older set of three
chromosomal pairs that were originated through pericentric inversions. A set of six large metacentric pairs formed by Robertsonian (Rb) translocations found in A. bahianus and A. chirurgus and a putative in tandem fusion found in A. chirurgus are more recent events. The
lack of interstitial telomeric sequences (ITS) in spite of several centric fusions in A. bahianus
and A. chirurgus might be related to the long period of time after their occurrence (estimated
in 5 M.a.). Furthermore, the homeologies among the chromosome pairs bearing ribosomal genes, in addition to other structural features, highlight large conserved chromosomal regions in the three species. Our findings indicate that macrostructural changes occurred during the cladogenesis of these species were not followed by conspicuous microstructural rearrangements in the karyotypes.
1. INTRODUÇÃO
1.1. Família Acanthuridae
A família Acanthuridae está inserida na Ordem dos Perciformes, subordem Acanthuroidei. Esta família conta com seis gêneros (Acanthurus, Ctenochaetus, Naso, Paracanthurus, Prionurus e Zebrasoma) subdivididos em 80 espécies (NELSON, 2006). É
caracterizada morfologicamente por indivíduos de pequeno a médio porte, podendo chegar a 36 cm, com um corpo alto, oval e comprimido lateralmente (RANDALL, 2001).
São conhecidos popularmente como peixes-cirurgião (surgeonfishes), devido à
presença de um espinho dobrável alojado dentro de uma bainha em cores, em ambos os lados do pedúnculo caudal, utilizado durante comportamentos agressivos inter e intraespecíficas. Tal característica constitui-se como a autapomorfia compartilhada pelo grupo (RANDALL, 2001).
Os cirurgiões são exclusivamente marinhos e distribuídos em mares tropicais e subtropicais de todo o mundo, sendo ausentes somente no Mediterrâneo (NELSON, 2006). A maioria das espécies de Acanthuriadae são herbívoras errantes diurnos com associação obrigatória a ambientes recifais (BELLWOOD; WAINWRIGHT, 2002; NELSON, 2006). A maioria das espécies de Acanthuridae têm estômagos com paredes finas e pastam em algas, e alguns têm estômagos semelhantes a moelas, com paredes espessas estes utilizam a areia para macerar detritos (FERREIRA et al., 2004; RANDALL, 2005).
Este grupo possui uma posição relevante do ponto de vista ecológico e evolutivo nas comunidades dos recifes tropicais. Sendo um importante agente físico (bioerosão) devido sua atividade de escavar o substrato contribuindo para sedimentação intensiva e biológica (herbivoria) influenciando na composição, desenvolvimento, abundância e padrão de distribuição da população de algas e comunidades associadas (HORN, 1989).
Ao longo de sua história de vida sofrem mudanças ontogenéticas sobre os aspectos da preferência por tipos de habitats (FERREIRA et al., 2004; ROCHA, 2002) com ovos planctônicos que eclodem após cerca de um dia, um período pelágico larval relativamente longo variando entre 51 até 75 o que lhes conferem enorme capacidade de dispersão e adultos sedentários associados a recifes de corais (THRESHER, 1984; MUSS et al., 2001; ROCHA et al., 2002, RANDALL, 2005).
ser encontrados constituindo grandes cardumes heterogêneos (RISK, 1998; LAWSON, et al., 1999; BELL e KRAMER, 2000; ROCHAet al., 2002).
1.2. Filogenia e evolução da Familia Acanthuridae
Acanthuridae é um grupo monofilético relativamente antigo, com cerca de 54 milhões anos (SORBINI e TYLER, 1998; TANG et al., 1999; SORENSON et al., 2013). A primeira aparição da família se deu no Eoceno Médio. A maioria dos acanthurideos fósseis conhecidos foi encontrada na região do Monte Bolca, no norte da Itália, que é a região mais representativa da rica ictiofauna recifal do Eoceno (BELLWOOD, 1996; CLEMENTS et al., 2003;. KLANTEN et al., 2004).
Embora muitos dos caracteres supraespecíficos que definem este grupo tenham uma duração prolongada no registro fóssil, o moderno conjunto de espécies parece ser de origem recente. Apoio à origem recente das espécies de peixes de recife é fornecido por McMillan e Palumbi (1997), que apresentaram argumentos para especiação recente ligada às mudanças no nível do mar, temperatura dos mares e clima global, ocorridas no Pleistoceno.
Questões referentes à filogenia desta família têm sido estudadas profundamente através de evidências morfológicas (WINTERBOTTOM, 1993; GUIASU e WINTERBOTTOM, 1993; TYLER et al., 1989) e análises moleculares (HOLCROFT e WILEY, 2008; KLANTEN et al., 2004; TANG et al., 1999). De forma geral, as relações filogenéticas dentro da família Acanthuridae estão bastante esclarecidas, entretanto, a relação entre os gêneros Acanthurus e Ctenochaetus tem gerado controvérsias ao longo das décadas.
Embora a possibilidade de parafiletismo entre esses dois grupos já tivesse sido sugerida com base em caracteres osteológicos compartilhados e caracteres merísticos (RANDALL, 1955; WINTERBOTTOM, 1993; GUIASU e WINTERBOTTOM, 1993), esta questão só foi elucidada por Clements et al. (2003) e mais tarde corroborada por Sorenson et al. (2013), estabelecendo uma filogenia mais compreensiva desta família através de análises
moleculares (Figura1a). Além da corroboração da hipótese de parafiletismo entre os gêneros
Acanthurus/Ctenochaetus, existem indícios de que os gêneros desta família são relativamente
antigos, com intensos episódios de especiação, pelo menos três ao longo da história filogenética (SORENSON et al., 2013; BERNAL e ROCHA, 2011).
Análises filogeográficas identificaram que A. bahianus, em relação às espécies A. chirurgus e A. coeruleus, parece altamente sensível a restrição ecológica imposta pela pluma
geográfica. De fato, as linhagens do Atlântico Sul e do Caribe de A. bahianus são, na verdade,
espécies diferentes, a linhagem caribenha foi nomeada como A. tractus (Figura 1b) (BERNAL
e ROCHA, 2011).
O plano corporal comum aos cirurgiões atuais apareceu no limite entre o Paleoceno/Eoceno Inferior, e passou por uma dramática radiação durante o Eoceno Médio (50 Ma). Dentro do gênero Acanthurus a irradiação da espécie A. coeruleus ocorreu em meados
do Mioceno (20 Ma) e a separação entre A. chirurgus e A. tractus (espécie irmã da A. bahianus)noFinal do Mioceno (10 m.a.) (SORENSON et al., 2013).
1.3. Aspectos biológicos e taxonômicos do gênero Acanthurus
O gênero Acanthurus possui ao todo 36 espécies, sendo o único, dentre toda a
família, a ocorrer no Atlântico Ocidental (RANDALL, 2002). Na costa brasileira podemos encontrar três espécies deste gênero: Acanthurus coeruleus Bloch & Schneider (1801), Acanthurus chirurgus (BLOCH, 1787), Acanthurus bahianus Castelnau (1855) (RANDALL,
1956; MENEZES e FIGUEIREDO, 1985; FLOETER et al., 2008; BERNAL e ROCHA, 2011).
A taxonomia detalhada das espécies citadas é representada por: Filo - Chordata
Subfilo - Craniata
Figura 1. (a) Hipótese filogenética para espécies do gênero Acanthurus, modificado de Sorenson et al. (2013).
(b) Relações entre espécies do gênero Acanthurus presentes no Atlântico, proposta por Bernal & Rocha (2011).
Superclasse - Gnasthostomata Classe - Actinopterygii Subclasse - Neopterygii Divisão - Teleostei Subdivisão - Euteleostei
Superordem - Acanthopterygii Série - Percomorpha
Ordem - Perciformes Subordem - Acanthuroidei Família - Acanthuridae Subfamília - Acanthurinae Tribo - Acanthurini
Gênero - Acanthurus Forsskål (1775)
Espécies - Acanthurus chirurgus (BLOCH, 1787)
Acanthurus coeruleus Bloch & Schneider (1801) Acanthurus bahianus Castelnau (1855)
A espécie A. coeruleus (Figura 2), exibe um comportamento territorialista, embora
A mais comum dentre as espécies de acanthurideos encontradas no litoral brasileiro,
A. chirurgus (Figura 3), geralmente, formam grandes cardumes, tem preferência por recifes
mais rasos e possui hábito alimentar semi-detritívoro (DIAS et al., 2001; FRANCINI-FILHO et al., 2010). Morfologicamente se diferencia das demais espécies por apresentar a nadadeira dorsal com 9 espinhos e 24-25 raios; a nadadeira anal com 3 espinhos e 22-23 raios e caudal emarginada. A coloração do corpo é acinzentada com 10 barras verticais mais escuras e o pedúnculo caudal mais claro que o restante do corpo (CERVIGÓN et al., 1993; FIGUEIREDO e MENEZES, 2000; CARPENTER, 2002).
A espécie A. bahianus (Figura 4) é caracterizada por apresentar na nadadeira dorsal
com 9 espinhos e 23-26 raios e anal com 3 espinhos e 21-23 raios. O padrão de cor do corpo é semelhante ao A. chirurgus, com ausência das barras verticais nos flancos e uma nadadeira
caudal lunada (CERVIGÓN et al., 1993; FIGUEIREDO e MENEZES, 2000; CARPENTER, 2002). Ocorre em grupos de cinco ou mais indivíduos, habitando recifes rasos de até 25
Figura 2. Espécime de Acanthurus coeruleus. Em destaque indivíduo na fase juvenil. Barra: 2 cm.
metros (ROCHA, 2002). Considerada também como semi-detritívora, tem em comum com A. chirurgus e algumas outras espécies do gênero, a presença de um estômago constituído por
paredes grossas semelhantes a uma “moela” onde a areia ingerida ajuda a triturar as algas (FERREIRA e GONÇALVES, 2006; DIAS et al., 2001; FRANCINI-FILHO et al., 2010).
1.4. Citogenética como ferramenta para inferências evolutivas
Análises citotaxonômicas contribuem significativamente para o estudo da evolução pelo fato do material genético estar contido nos cromossomos (ALMEIDA-TOLEDO, 1998). Portanto, alterações como, rearranjos cromossômicos, polimorfismos estruturais e/ou numéricos, poliploidia natural, sistemas de cromossomos sexuais entre outros, são quase sempre significativas para que se possa inferir o rumo evolutivo das espécies (GUERRA, 1988).
As técnicas citogenéticas convencionais (Giemsa, banda-C, Ag-RONs) e moleculares como a marcação com fluorocromos base-específicos (DAPI/MM) e o desenvolvimento nos últimos anos, de novas ferramentas de análise mais aprofundadas como a técnica de Hibridação in situ Fluorescente (FISH), são de grande importância na identificação de
diversificação cromossômica. Juntas, a citogenética clássica e a molecular têm fornecido contribuições importantes para o entendimento da composição e estrutura dos cromossomos e em estudos que visem à formulação de hipóteses filogenéticas (BERTOLLO et al., 1978; FELDBERG et al., 2003; MARTINS et al., 2011).
A técnica por impregnação por nitrato de prata (Ag-RONs) é utilizada para evidenciar as Regiões Organizadoras de Nucléolos (RONs), porção particulalrmente variável do genoma eucarioto, as quais possuem sítios ribossômicos ativos, que podem estar
localizados nas posições terminais ou intersticiais dos cromossomos (FORESTI e TOLEDO, 1985). A caracterização do número e posição das RONs tem sido muito utilizada em peixes e pode constituir um excelente marcador citotaxonômico para alguns grupos (GALETTI, 1998; MOLINA et al., 2002).
Os bandamentos cromossômicos foram as primeiras ferramentas usadas para comparação de genomas, pois, espécies próximas apresentam um padrão muito similar de bandas tornando o estudo comparativo viável. O bandamento C permite a localização de regiões ricas em heterocromatina constitutiva (sequências de DNA altamente repetitivas). Em estudos evolutivos, a análise de heterocromatina pode fornecer dados para a caracterização de ocorrência de rearranjos em espécies próximas (MEDRANO et al., 1988).
O uso de fluorocromos é muito importante, principalmente DAPI e MM tem sido muito usados para discriminar a composição de bases da região heterocromática e o número e localização dos sítios de DNA ribossomal (MARTINS et al., 2011).
Com o desenvolvimento da técnica de hibridação in situ fluorescente (FISH), a
citogenética obteve avanços significativos para o conhecimento do genoma com base nos cromossomos (MARTINS et al., 2011). Esta técnica definiu a transição da era clássica da citogenética à era molecular, permitindo estudos mais detalhados com a integração da informação molecular das sequências de DNA e sua localização física ao longo dos cromossomos e genomas (SCHWARZACHER, 2003; JIANG e GILL, 2006).
Análises citogenéticas moleculares utilizando a técnica de FISH são atualmente muito utilizadas e vêm se mostrando eficientes ferramentas no estudo da evolução cariotípica em peixes, devido ao mapeamento de sequências ribossomais 5S e 18S (MARTINS e GALETTI, 2001; MOLINA, 2002; MARTINS et al., 2011a; MOTTA-NETO et al., 2011b; CALADO et al., 2013; CALADO et al., 2014).
Informações citogenéticas em peixes marinhos têm aumentado nas últimas décadas, Entretanto, somente cerca de 7% das espécies, em sua maioria da Ordem Perciformes, grupo mais representativo entre os teleósteos (NELSON, 1996; GALLETTI et al., 2000). Onde, em grande parte de suas espécies, um padrão marcante de conservadorismo cromossômico, com número diploide composto por 48 cromossomos acrocêntricos (MOLINA, 2007; MOTTA-NETO et al., 2011).
que se mostram adaptadas ou isoladas em ambientes singulares, como, as que são associadas a recifes de corais e ilhas oceânicas, onde as massas de água que os circundam podem funcionar como a barreira geográfica e limitar a dispersão (PALUMBI, 1994; ROCHA, 2003).
Os peixes recifais são principalmente compostos por Perciformes e contêm uma elevada diversidade de espécies que representam linhagens evolutivas distintas. Dentre os Perciformes um dos grupos mais representativos e carismáticos é a família Acanthuridae (ROBERTSON, 1983; LAWSON et al., 1999), que devido a sua diversidade na forma de forrageamento e sua forte associação com recifes de corais são um grupo bastante utilizado em estudos que busquem identificar o ritmo de evolução que conduz a biodiversidade em peixes de corais, e na investigação da importância dos fatores ecológicos na diversificação de ambientes recifais (SORENSON et al., 2013).
A família Acanthuridae é relativamente bem estudada, sobretudo sob aspectos da evolução morfológica (TYLER, 1970a, b; GUIASU e WINTERBOTTOM, 1993; BORDEN, 1998; WINTERBOTTOM, 1993), e da filogenia molecular (ROCHA et al., 2002; CLEMENTS et al., 2003; BERNAL e ROCHA, 2011; SORENSON et al., 2013). Entretanto, no que tange a evolução cromossômica, a família Acanthuridae ainda está sub-representada, menos de 5% das espécies, sendo todas no oceano Pacífico (ARAI, 2011).
As três espécies analisadas do Pacífico, Acanthurus triostegus (LINNAEUS, 1758), Prionorus scalprum Valenciennes, 1835 (ARAI e INOUE, 1976) e Ctenochaetus striatus
2. OBJETIVOS
2.1. Objetivo geral
Tendo em vista a importância das informações citogenéticas e a carência delas no que diz respeito às espécies da família Acanthuridae, o presente trabalho tem como foco principal estabelecer aspectos carioevolutivos, com base em análises da citogenética clássica e molecular de três espécies da família Acanthuridae presentes no Atlântico Ocidental.
2.2 Objetivos específicos
Caracterizar cromossomicamente por meio de bandamento C, Ag-RONs e coloração com fluorocromos base-específicos MM/DAPI as espécies A. coeruleus, A. bahianus e A. chirurgus.
Mapear as sequências das subunidades ribossomais DNAr 5S e 18S por meio de dual-color FISH das espécies analisadas das famílias Acanthuridae.
3. MATERIAL E MÉTODOS 3.1. Material
Os exemplares das espécies A. coeruleus (n=23), A. bahianus (17) e A. chirurgus
(12) foram coletados ao longo da costa de Natal, Rio Grande do Norte (5o46'S, 35o12'O) e Salvador, Estado da Bahia (13°00'S, 38°32'O), nordeste do Brasil. As coletas ocorreram por meio de mergulho livre em recifes costeiros nas áreas citadas utilizando-se de apetrechos de pesca como puçás e tarrafas. Após a coleta os exemplares foram transportados vivos ao Laboratório de Genética de Recursos Marinhos, da Universidade Federal do Rio Grande do Norte, e mantidos em aquários aerados até que se procedesse à realização de preparações cromossômicas.
3.2. Métodos
3.2.1. Obtenção de cromossomos mitóticos
Vinte e quatro horas antes da preparação cromossômica, os animais foram inoculados via intramuscular com uma solução de complexos antígenos para indução mitótica (MOLINA et al., 2010). Após esse período, os exemplares foram anestesiados e sacrificados seguindo procedimento estabelecido por Blessaing et al. (2010).
Para obtenção de cromossomos mitóticos, fragmentos do rim anterior foram adicionados em meio de cultura RPMI 1640 de acordo com a técnica in vitro preconizada por
Gold et al. (1990). Os fragmentos foram dissociados em 9 ml de meio de cultura RPMI 1640 através de aspirações com seringas de vidro de 10 ml até obter um mistura homogênea, onde foi adicionado 5 gotas de colchicina 0,025%, deixando agir por 30 minutos em temperatura ambiente. Após o processo anterior o material foi centrifugado por 10 minutos a 1000 rpm. Após descarte do sobrenadante, foi adicionado 8 ml da solução hipotônica de KCl a 0,075M, que após ser homogeneizada agiu por 28 minutos em temperatura ambiente. A suspensão celular foi pré-fixada com 0,5 ml de solução de metanol e ácido acético (3:1) recém-preparado. Após homogeneização a suspensão foi centrifugada por 10 minutos. O processo de fixação do material foi repetido por 3 vezes, após o que as suspensões celulares foram estocados em tubos Eppendorf de 1,5 ml à -20oC.
3.2.2. Análises cromossômicas e montagem do cariótipo
As análises foram realizadas em microscópio óptico de epifluorescência Olympus™ BX51 e fotografadas sob aumento de 1.000X através de sistema digital de captura Olympus DP70 utilizando o software DPController 1.2.1.108 (Olympus Optical Co. Ltd.). Cerca de trinta metáfases foram analisadas para cada espécime. As melhores metáfases foram selecionadas para montagem do cariótipo a partir da nomenclatura preconizada por Levan et al. (1964) com modificações, onde os cromossomos foram definidos, quanto à posição dos centrômeros, em metacêntricos (m), com a razão entre o braço maior e menor (RB) variando de 1,00 a 1,70; submetacêntricos (sm), RB=1,71–3,00; subtelocêntricos (st), RB=3,01–7,00; e acrocêntricos (a), RB>7,01.
3.2.3. Detecção de regiões organizadoras de nucléolos
A detecção de regiões organizadoras de nucléolos foi obtida pela técnica de impregnação por prata preconizada por Howell & Black (1980). Sobre uma lâmina previamente preparada com suspensão celular foram adicionadas 7 gotas de solução gelatinosa (1g de gelatina incolor, dissolvida em 50ml água e 0,5ml ácido fórmico), e 6 gotas de solução de nitrato de prata (AgNO3) à 50%. A solução foi homogeneizada com a extremidade de uma lamínula e recoberta por ela sendo então incubada em estufa a 60°C, até obtenção da cor amarelo escuro. Após a remoção da lamínula com jatos de água destilada, eventualmente, para destacar as marcações argênteas, as preparações podiam ser coradas por cerca de 20 segundos com solução de Giemsa a 5%.
3.2.4. Detecção de heterocromatina constitutiva
A análise das regiões heterocromáticas foi realizada através do bandamento-C, segundo Sumner (1972). Foram utilizadas lâminas com material envelhecido em estufa a 37°C por no mínimo 3 dias, após esse período a lâmina foi imersa em HCl 0,2 N por 14 minutos em temperatura ambiente, sendo lavada com água destilada e seca. Posteriormente a lâmina foi mergulhada em uma solução a 5% de Ba(OH)28H2O à 42°C por cerca de 1 minuto e 35 segundos sendo rapidamente lavadas em HCl 0,1N e posteriormente água destilada. Após secar ao ar, a lâmina foi então imersa em solução salina 2x SSC a 60°C em estufa por 40 minutos. Para análise o material foi corado com Giemsa 5% por 8 minutos.
3.2.5. Dupla coloração com fluorocromos base-específicos
pares de bases AT, de acordo com Carvalho et al. (2005). As lâminas com suspensão celular de cada exemplar foram coradas com 30µl de (MM) 0,5 mg/ml cobertas com lamínulas e depositadas em câmara úmida, no escuro, sendo lavadas com água destilada após 60 minutos, coradas com 30 µl de DAPI 2µl/ml, cobertas por lamínulas e colocada em câmara úmida, no escuro, por 30 minutos. Posteriormente as lâminas foram lavadas com água destilada e montadas com tampão glicerol-Mcllvaine pH 7,0 (1:1) coberto com lamínula e selado com esmalte incolor. As lâminas foram então guardadas à 4oC em câmara escura e analisadas após 3 dias (DAPI) e 15 dias (MM) com fotomicroscópio de epifluorescência (Olympus™ BX51) com filtros apropriados, em aumento de 1.000x. As preparações foram capturadas pelo sistema digital Olympus DP73 com uso do software DPController, v. 1.2.1.108 (Olympus Optical Co. Ltd.).
3.2.6. Hibridação in situ fluorescente – FISH
3.2.6.1. Obtenção de sondas para hibridização
A FISH foi realizada de acordo com Pinkel et al. (1986) usando sondas teloméricas de 18S e 5S. As sondas de DNAr 5S (200 pb) e 18S DNAr (1400 pb) foram obtidos a partir de amostras de DNA de A. coeruleus via PCR utilizando os iniciadores A 5'-TAC CCG CGA
TCT CGT CCG ATC-3 '/ B 5'-CAG GGT GCT ATG CCG GTA AGC-3' (PENDAS et al., 1994) e NS1 5'-GTC ATA GTA TGC TTG TCT C-3' / NS8 5'-TCC GCA GGT TCA CCT ACG AG-3' (WHITE et al., 1990), respectivamente. A sonda 18S foi marcada por nick translation digoxigenina-11-dUTP (Roche) e a sonda DNAr 5S foi marcada por nick translation biotina-14-dATP (Roche) , de acordo com as especificações do fabricante. As
sequências (TTAGGG)n foram mapeadas pela técnica de FISH com Kit FISH Telomere PNA/FITC de acordo com as instruções do fabricante (DakoCitomation).
3.2.6.2. Hibridização cromossômica
FISH (Fluorescence In Situ Hybridization) foi realizada em metáfases mitóticas
(PINKEL et al., 1986) das três espécies. Análise por dual-color FISH foi desenvolvida usando
4. Capítulo I
Abstract
Surgeonfishes are a species-rich group and a major biomass on coral reefs. Three species are commonly found throughout South Atlantic, Acanthurus bahianus, A. chirurgus, and A. coeruleus. In this paper, we present the first cytogenetic data of these species, revealing a
sequential chromosomal diversification. A. coeruleus was characterized by a relatively
conserved karyotype evolved by pericentric inversions of some pairs (2n = 48, 2sm + 4st + 42a). In contrast, the karyotypes of A. bahianus (2n = 36) and A. chirurgus (2n = 34) were
highly differentiated by the presence of six large metacentric pairs in A. bahianus (12m + 2sm
+ 4st + 18a) and A. chirurgus (12m + 2sm + 4st +1 6a) probably derived by chromosomal
fusions that corroborate their closer relationship. A discernible in tandem fusion represents an autapomorphic character to A. chirurgus. In spite of macrostructure variation, single nucleolar
organizer regions (NORs) on short arms of a subtelocentric pair and similar distribution of C-bands were observed in the three species. Overlapping of chromosomal data with molecular phylogeny indicated pericentric inversions which took place nearly at 19Ma while centric fusions are as recent as 5Ma. A physical mapping of coding and noncoding sequences in
Acanthurus could clarify the role of additional rearrangements during their chromosomal
1. Introduction
Acanthuridae are a monophyletic fish family composed of about 80 species, popularly known as surgeonfishes or tangs (NELSON, 2006). This is an ancient group (nearly 54Ma) and most of genera (Acanthurus, Ctenochaetus, Naso, Paracanthurus, Prionurus e Zebrasoma) diverged between 17 and 21 Ma in Early Miocene (NELSON, 2006).
Apparently, the Pacific Ocean is the center of origin of surgeonfishes, retaining most of Acanthuridae richness (TANG et al., 1999; GUIASU e WINTERBOTTOM, 1993). Further colonization events resulted in distribution of this family to virtually all tropical and subtropical seas of the world, but Mediterranean Sea (SORENSON et al., 2013; TANG et al., 1999) The genus Acanthurus is the largest within the family, but monophyly of the genus is
still controversial (TANG et al., 1999; CLEMENTS et al., 2003). This fish group is morphologically and ecologically diversified, mainly in relation to foraging behavior and dentition, composing one of themost representative herbivorous fish group on coral reefs (TANG et al., 1999; FRANCINI-FILHOet al., 2010)
A total of four species of Acanthuridae, all belonging to the genus Acanthurus, are
present in western Atlantic (BERNAL e ROCHA, 2011). Three species are common along the Brazilian coast (Western South Atlantic): Acanthurus coeruleus (blue tang), A. bahianus
(barber surgeonfish), and A. chirurgus (doctorfish) (BERNAL e ROCHA, 2011;
FIGUEIREDO e MENEZES, 2000). Another Acanthurus species (A. monroviae) was also
recorded off southeastern coast of Brazil, but it seems to be an occasional occurrence (LUIZ JR. et al., 2004). Moreover, A. bahianus was thought to range from USA to southern Brazil,
but morphological and genetic analyses have shown that populations from Massachusetts to Caribbean actually refer to another species, validated as A. tractus (FIGUEIREDO e
MENEZES, 2000).
In spite of the low diversity of Atlantic species when compared to Pacific and Indian oceans, surgeonfishes are a dominant fish group forming large assemblages in several reef areas from South Atlantic (TANG et al., 1999). A. coeruleus specimens are usually solitary
due to their territoriality behavior, while A. bahianus and A. chirurgus are commonly found in
However, reports about cytogenetic patterns of Acanthuridae are still underrepresented (less than 5% of species) and restricted to Indo-Pacific species (ARAI, 2011). The three analyzed species from Pacific, Acanthurus triostegus, Prionurus scalprum
(ARAI & INOUE 1976), and Ctenochaetus striatus (OJIMA e YAMAMOTO, 1990) all share
a conservative Perciformes-like karyotype with 2� = 48a, considered basal to this fish group (BRUM e GALETTI, 1997).
In order to increase the karyotypic data of Acanthuridae and to infer the chromosomal evolution of Atlantic species, cytogenetical analyses were carried out for three Acanthuridae species from Brazilian coast, South Atlantic.
2. Material and Methods
Nine individuals of A. coeruleus, four individuals of A. bahianus, and 17 individuals
of A. chirurgus were cytogenetically studied. Animals were collected using hand nets (60x100
cm) by snorkeling at coastal reef areas from the states of Rio Grande do Norte (5°46’S, 35°12’W) and Bahia (13°00’S, 38°32’W and 13°52’S, 38°56’W) in northeastern Brazilian shore (Figure 1). Right after collection, specimens were transported in plastic bags with
Figure 1. Map of South America showing the collections sites of Acanthurus coeruleus (a), A. bahianus (b),
oxygen to the laboratories and placed in 60 L tanks equipped with filtration and aeration systems.
Twenty-four hours prior to chromosomal preparation, the animals were inoculated via intramuscular with a solution of antigen complexes (Munolan) for mitotic induction (MOLINA et al., 2010). After this period, the specimens were anesthetized and euthanized by immersion in in water at 0–4º Cup to complete interruption of gill movements (Blessaing et al., 2010). To obtain mitotic chromosomes in vitro, portions of anterior kidney were removed
and transferred to RPMI medium (Cultilab) with about 50 �L of 0.025% colchicine, followed by hypotonic treatment (KCL 0.075M) for 20 minutes at 37∘C and fixation in Carnoy’s fixative (methanol : acetic acid 3 : 1) (GOLD et al., 1990). Chromosomes were stained with 5% Giemsa in phosphate buffer (pH 6.8) for karyotypic analyses. Nucleolus organizer regions (NORs) were detected by silver nitrate staining (Ag-NORs) (HOWELL e BLACK, 1980), whereas heterochromatic regions were evidenced by Cbanding (SUMNER, 1972).
Metaphases were photographed using an Olympus BX51 (Olympus, Tokyo, Japan) epifluorescence photomicroscope equipped with digital capture system. Chromosomes were classified as metacentric (m), submetacentric (sm), subtelocentric (st), and acrocentric (a) based on arm ratio (LEVAN, 1964). The pairs were arranged in decreasing order size according to each morphological category (m, sm, st, and a) in karyotypes using the software Adobe Photoshop CS6 v. 13.0.
3. Results
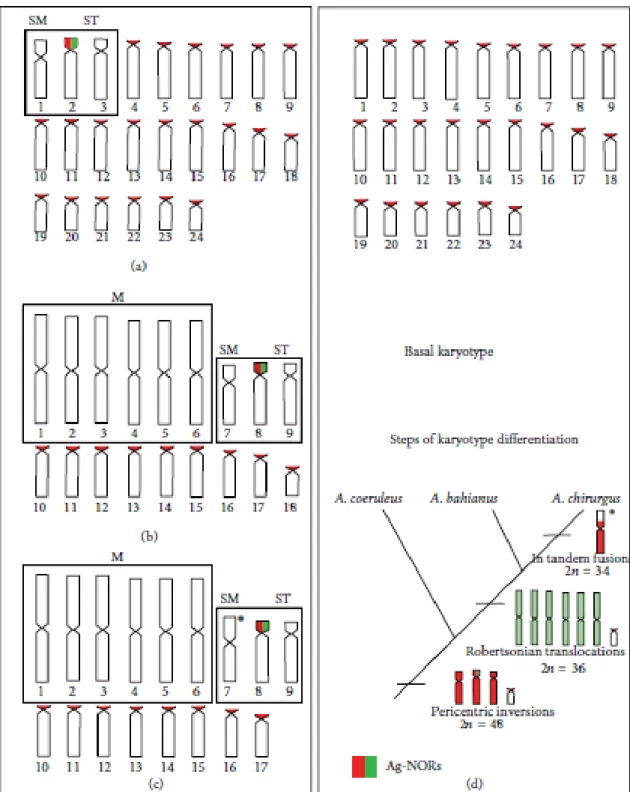
The three Acanthurus species showed remarkable karyotype diversification. Acanthurus coeruleus presented 2�=48, composed of two submetacentric, four subtelocentric,
and 42 acrocentric chromosomes (Figure 2(a)). The diploid number of A. bahianus equals
2�=36 with a karyotype composed of 12 large metacentric, two submetacentric, four subtelocentric, and 18 acrocentric chromosomes (Figure 2(c)) while A. chirurgus was
characterized by 12 large metacentric, two submetacentric, four subtelocentric, and 16 acrocentric chromosomes (2�=34) (Figure 2(e)).
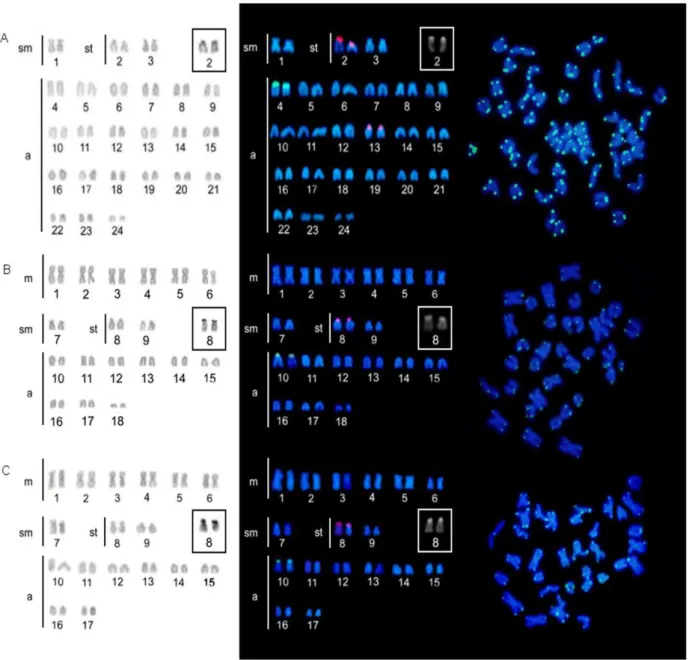
Small amounts of heterochromatin were detected mainly at pericentromeric regions and interspersed with NORs in studied species (Figures 2(b), 2(d), and 2(f)). In A. bahianus,
Single nucleolar organizer regions (NORs) were located by silver nitrate staining on short arms of the largest subtelocentric pair in the three Acanthuridae species (Figure 2, inbox). Based on chromosomal data, idiograms were generated to highlight particular karyotype traits for each species and the inferred pathways of chromosomal differentiation based on a phylogenetic hypothesis to Atlantic Acanthurus (Figures 3(a)–3(d)).
Figure 2: Karyotypes of Acanthurus coeruleus ((a) and (b)) with 2�=48, A. bahianus ((c) and (d)) with 2�=36,
and A. chirurgus ((e) and (f)) with 2�=34 after conventional Giemsa staining ((a), (c), and (e)) and
4. Discussion
It is assumed that the presence of 48 acrocentric chromosomes represents a plesiomorphic feature within Perciformes (BRUM e GALETTI JR., 1997; GALETTI JR. et al., 2006). This condition is particularly frequent among marine fish and could be related to
Figure 3. Idiograms of chromosomes sets of Acanthurus coeruleus (a), A. bahianus (b), and A. chirurgus (c)
dispersal abilities (high gene flow) between populations, thereby preventing the fixation of new chromosomal rearrangements and karyotypic divergence (MOLINA, 2007). In fact, the low genetic structure in reef fish species has been correlated to the production of planktonic eggs and/or larvae that can be dispersed over large distances (EBLE et al., 2011). This trend (2n=48a) seems to be valid for Acanthuridae species from Indo-Pacific Ocean of different genera, such as Acanthurus, Ctenochaetus, and Prionurus (ARAI, 2011). However,
inconsistent relationship between pelagic larval duration (PLD) and genetic connectivity or chromosomal patterns has been reported in some marine species (ROCHA et al., 2002; AFFONSO e GALETTI JR., 2005). In these cases, ecological and biogeographic aspects of each species might be more relevant to explain the genetic variation than PLD itself, as observed in the present study.
As expected for widely distributed species with long PLD, A. coeruleus presented
typical Perciformes-like features, that is, a diploid number of 48, single NORs, and a large number of acrocentric chromosomes (Figure 2a).The karyotype of this species (2sm + 4st + 42a) demonstrates the occurrence of pericentric inversions in three chromosome pairs (1st, 2nd, and 3rd pairs), a common rearrangement in Perciformes that accounts for most of karyotype diversification in marine fish (GALETTI JR. et al., 2006).
A similar set of three chromosomal pairs in both morphology and size is also observed in A. bahianus and A. chirurgus, represented by a submetacentric pair and two
subtelocentric pairs, including the NOR-bearing pair. Because of the high resemblance of such pairs in the three Acanthurus species, they are supposed to share a common origin before
the differentiation of each lineage, thereby indicating a symplesiomorphic trait. Estimates of divergence time between the subclade that comprises A. coeruleus and that clusters A. chirurgus and A. tractus (SORENSON et al., 2013), a sister-species of A. bahianus, suggest
that these putative homeologous pairs (sm and st) had arisen at nearly 19Ma.
On the other hand, A. bahianus and A. chirurgus presented an evolutionary
chromosomal pattern rarely found in typical marine fishes. The drastic reduction in diploid number from 48 chromosomes to 2n=36 and 2n=34, respectively, along the presence of large metacentric pairs is evidence of sequential Robertsonian rearrangements or centric fusions (Figures 2c and 2e). Indeed, the uniqueness of Robertsonian translocations in karyotypes of A. bahianus and A. chirurgus (Figure 3), and similar size of metacentric pairs
karyotype since this species has an exclusive large sm pair (7th) and lacks the smallest acrocentric pair observed in the other two species (Figure 2e).
The chromosomal speciation observed in Acanthurus species of South Atlantic is
likely to reflect historical events. Extensive analysis of biogeography and evolution of reef fish from Atlantic indicated that changes in ocean dynamics over the past 10Ma have determined the differential richness and endemism levels of fish genera and families of reef fish (FLOETER et al., 2008). As discussed by Galetti et al. (2006) the rate of chromosomal evolution in reef fish from Atlantic Ocean also seems to be strongly related to habitat isolation of coastal areas during glaciation periods followed by further sea level uprising.
Unfortunately, no reports about time of divergence between A. bahianus and A. chirurgus are available, thus hindering the minimum time span after Robertsonian
rearrangements that gave rise to the large metacentric chromosomes, herein referred as a single trait. However, estimates inferred for A. chirurgus and A. tractus (SORENSON et al.,
2013) being the latter a sibling species of A. bahianus (FRANCINI-FILHO et al., 2010),
point out that these rearrangements took place by at least 5Ma. Even though the time estimates for the occurrence of chromosomal fusions in both Acanthurus species might
require some bias correction, they are intermediary to periods of major biogeographic isolation events in Atlantic Ocean such as Amazon outflow (∼10Ma) and uplifting of Panama isthmus (∼3Ma) (Rocha, 2003). Nonetheless, the influence of these biogeographic barriers in the putative fixation of chromosomal rearrangements remains unclear and further cytogenetic studies in other species, particularly A. tractus, are highly encouraged.
Different from pericentric inversions, centric fusions or Robertsonian rearrangements are usually reported in non-Perciformes marine fish, such as flatfish (Pleuronectiformes) (BITENCOURT et al., 2014) toadfish (Batrachoidiformes) (COSTA e MOLINA, 2009) and some mullets (Mugiliformes) (NIRCHIO et al., 2005) Centric fusions are particularly common in Batoidea (stingrays, guitarfish, and skates), the most derived superorder of elasmobranchs (ROCCO et al., 2007). Conversely, these rearrangements have been scarcely identified in Perciformes at a polymorphic stage in Pomacentridae (MOLINA e GALETTI JR., 2002) Gobiidae (GILES et al., 1985; THODE et al.,1985), Lutjanidae (NIRCHIO et al., 2008), Apogonidae (OJIMA e KOJIMA, 1985) and Uranoscopidae (VITTURI et al., 1991) or else restricted to a particular taxon like Sparisoma (parrotfishes) (SENA e MOLINA, 2007). Therefore, the karyotypes of A. bahianus and A. chirurgus can be regarded as highly
While the macrostructure was variable, the NOR-bearing chromosomes seem to be conserved in the three species (Figure 2, inbox). This pattern indicates that these chromosomal regions as poor cytotaxonomic markers, differing from other Atlantic fishes in which the identification of ribosomal cistrons has proved to be efficient to distinguish apparently homogeneous karyotypes, as in Lutjanidae (ROCHA e MOLINA, 2008) Serranidae (MOLINA et al., 2002) and Gerreidae (CALADO et al., 2012) or even population units (ACCIOLY et al., 2012) Similarly, heterochromatin was virtually similar among A. coeruleus, A. bahianus, and A. chirurgus, being mainly dispersed over
centromeres and NORs, as commonly found in most Perciformes (MOLINA et al., 2013) Therefore, it is unclear if the deep divergences in these fish karyotypes are followed by microstructural changes. Further analyses using other banding methodologies and mapping of sequences by fluorescence in situ hybridization (FISH) are required to evaluate the extension of such apparent homogeneity of specific chromosomal regions in Acanthurus.
The amount of chromosomal traits in the three Acanthurus species from Brazilian
coast allows raising a phylogenetic hypothesis to them. Indeed, the ordination and sharing of the traits show a closer phylogenetic relationship between A. bahianus and A. chirurgus than
to A. coeruleus. This result is corroborated by previous genetic analyses. Indeed, analysis of
CytB sequences showed a more basal condition between A. coeruleus in relation to the other
two congeners (FRANCINI-FILHO et al., 2010). Recently, a phylogenetic analysis of
Acanthuridae based on sequence data of two mitochondrial and seven nuclear genes (SORENSON et al., 2013) corroborated the ancestral position of A. coeruleus in relation to A. chirurgus and A. tractus, which replaces A. bahianus in the Caribbean
(FRANCINI-FILHO et al., 2010; CALASTELLANOS-GELL et al., 2012).
Thus, for analyzed Atlantic species, the chromosomal traits show robust support to clarify the phylogenetic arrangement among them serving as useful markers to evolutionary studies in Acanthuridae. Moreover, in agreement with phylogeographic studies (ROCHA et al., 2002; FLOETER et al., 2008), the chromosomal differences between Atlantic species of
Acanthurus seem to be more related to ecology and evolutionary history than to dispersal
potential since the three species share a relatively long PLD.
5. Conclusion
In conclusion, the chromosomal analyses in Acanthurus allowed identifying
karyotype modification can be inferred from the most basal pattern, involving few structural rearrangements (pericentric inversions in A. coeruleus) to high derived ones, originated by
Robertsonian fusions in both A. bahianus and A. chirurgus and additional in tandem fusion in A. chirurgus. This scenario reveals a unique condition to tracing back the order of
chromosomal evolutionary changes in Atlantic surgeonfish.
Acknowledgments
The authors thank CAPES, CNPq (no. 556793/2009-9), INCT “Ciências do Mar,” and FAPESB (no. APP0064/2011) for the financial support and ICMBio/SISBIO (licenses 19135-1, 131360-1, and 27027-2) for the authorization in collecting specimens. They are also grateful to Dr. José Garcia Júnior for the taxonomic identification of specimens.
References
ACCIOLY, I. V.; et al. Chromosomal population structuring in carangids (Perciformes) between the north-eastern and southeastern coasts of Brazil, African Journal of Marine Science, vol. 34, no. 3, pp. 383–389. 2012.
AFFONSO, P. R. A.; M. and GALETTI JR., P. M. Chromosomal diversification of reef fishes from genus Centropyge (Perciformes, Pomacanthidae), Genetica, vol. 123, no. 3, pp.
227–233, 2005.
ARAI, R. and INOUE, M. Chromosomes of seven species of Pomacentridae and two species of Acanthuridae from Japan, Bulletin of the National Science Museum, 2:73–78, 1976.
ARAI, R. Fish Karyotypes. A Check List, Springer, Tokyo, Japan, 2011.
BERNAL, M.A.; ROCHA, L.A. Acanthurus tractus Poey, 1860, a valid western Atlantic
species of surgeonfish (Teleostei, Acanthuridae), distinct from Acanthurus bahianus
Castelnau, 1855. Zootaxa, Vol. 2905, pp. 63-68, 2011.
BLANCO, D.R.; et al. Comparative cytogenetics of giant trahiras Hoplias aimara and H. intermedius (Characiformes, Erythrinidae): chromosomal characteristics of minor and
major ribosomal DNA and cross-species repetitive centromeric sequences mapping differ among morphologically identical karyotypes. Cytogenet. Genome Research.
Vol. 132, pp.71-78, 2011.
BLESSING, J. J.; MARSHALL, J. C. and BALCOMBE, S. R. “Humane killing of fishes for scientific research: a comparison of two methods,” Journal of Fish Biology, vol. 76, pp. 2571–2577, 2010.
BRUM, M. J. I. and GALETTI JR., P. M. Teleostei ground plan karyotype,” Journal of Comparative Biology, vol. 2, pp. 91–102, 1997.
telomere sequences (ITS). Journal of Experimental Marine Biology and Ecology, vol
.452, pp. 101-104, 2014.
CALADO, L.L.; et al. Evolutionary dynamics of rDNA genes in the chromosomes of the
Eucinostomus fishes: cytotaxonomic and karyoevolutive implications. Genetics Molecular Research, 2014 .
CALADO, L.L.; et al. Cytogenetic studies of Atlantic mojarras (Perciformes: Gerreidae): chromosomal mapping of 5S and 18S ribosomal genes using double FISH.
Aquaculture Research, vol. 44, pp. 829-835, 2013.
CALADO, L. L. et al. Cytogenetic studies of Atlantic mojarras (Perciformes: Gerreidae): chromosomal mapping of 5S and 18S ribosomal genes using double FISH, Aquaculture Research, vol. 44, pp. 829–835, 2012.
CAPUTO, V.; et al. Heterochromatin heterogeneity and chromosome variability in four species of gobiid fishes (Perciformes: Gobiidae). Cytogenetics and Cell Genetics, vol.
79, pp. 266-271, 1997.
CASTELLANOS-GELL, J. et al. The surgeonfish, Acanthurus bahianus, has crossed the
Amazon–Orinoco outflow barrier. Marine Biology, vol. 159, pp. 1561-1565, 2012.
CIOFFI, M.B.; MARTINS, C.; BERTOLLO, L.A.C. Comparative chromosome mapping of repetitive sequences. Implications for genomic evolution in the fish, Hoplias malabaricus. BMC Genetics, vol. 10, pp. 1-8, 2009.
CIOFFI, M.B. et al. Chromosomal distribution of repetitive DNA sequences highlights the independent differentiation of multiple sex chromosomes in two closely related fish species. Cytogenetic Genome Research, vol. 134, pp. 295-302, 2011.
CLEMENTS, K. D.; GRAY, R.D. and CHOAT, J.H. Rapid evolutionary divergences in reef fishes of the family Acanthuridae (Perciformes: Teleostei), Molecular Phylogenetics and Evolution, vol. 26, no. 2, pp. 190–201, 2003.
COSTA, G. W. W. F. and MOLINA, W. F. Karyoevolution of the toadfish Thalassophryne nattereri (Batrachoidiformes: Batrachoididae), Genetics and Molecular Research, vol.
8, no. 3, pp. 1099–1106, 2009.
EBLE, J. A. et al. Not all larvae stay close to home: insights into marine population connectivity with a focus on the brown surgeonfish (Acanthurus nigrofuscus), Journal of Marine Biology, vol. 2011, Article ID 518516, 12 pages.
FLOETER, S. R.; ROCHA, L. A.; ROBERTSON D. R. Atlantic reef fish biogeography and evolution, Journal of Biogeography, vol. 35, no. 1, pp. 22–47, 2008.
FIGUEIREDO, J. L. and MENEZES, N. A. Manual de Peixes Marinhos do Sudeste do Brasil. VI. Teleostei (5), Museu de Zoologia, Universidade de São Paulo, São Paulo,
Brazil, 2000.
interference competition. Journal of the Marine Biological. Association of the United
Kingdom, vol. 90, no. 3, pp. 481–492, 2010.
GALETTI JR., P. M.; et al. Assessing genetic diversity of Brazilian reef fishes by chromosomal and DNA markers, Genetica, vol. 126, no. 1-2, pp. 161–177, 2006 .
GILES, V.; THODE, G. and ALVAREZ, M. C. A new Robertsonian fusion in the multiple chromosome polymorphism of a Mediterranean population of Gobius paganellus
(Gobiidae, Perciformes), Heredity, vol. 55, pp. 255–260, 1985.
GOLD, J. R. et al. Improved methods for working with fish chromosomes with a review of metaphase chromosome banding,” Journal of Fish Biology, vol. 37, no. 4, pp. 563–
575, 1990.
GUIASU R. C. and WINTERBOTTOM, R. Osteological evidence for the phylogeny of recent genera of surgeonfishes (Percomorpha, Acanthuridae), Copeia, no. 2, pp. 300–
312, 1993.
HOWELL, W. M. and BLACK, D. A. Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: a 1-step method, Experientia, vol. 36,
no. 8, pp. 1014–1015, 1980.
LAWSON, G. L.; KRAMER, D. L. and HUNTE, W. Size-related habitat use and schooling behavior in two species of surgeonfish (A. bahianus and A. coeruleus) on a fringing
reef in Barbados, West Indies, Environmental Biology of Fishes, vol. 54, no. 1, pp. 19–
33, 1999.
LEVAN, A.; FREDGA, K. and SANDBERG, A. A. Nomenclature for centromeric position on chromosomes, Hereditas, vol. 52, pp. 201–220, 1964.
LUIZ JR., O. J. et al. The occurrence of Acanthurus monroviae (Perciformes: Acanthuridae)
in the south-western Atlantic, with comments on other eastern Atlantic reef fishes occurring in Brazil, Journal of Fish Biology, vol. 65, no. 4, pp. 1173–1179, 2004.
MOLINA, W. F. and GALETTI JR., P. M. Robertsonian rearrangements in the reef fish
Chromis (Perciformes, Pomacentridae) involving chromosomes bearing 5s rRNA
genes, Genetics and Molecular Biology, vol. 25, no. 4, pp. 373–377, 2002.
MOLINA, W. F.; MAIA-LIMA, F. A. and AFFONSO, P. R. A. M. Divergence between karyotypical pattern and speciation events in Serranidae fish (Perciformes),
Caryologia, vol. 55, no. 4, pp. 299–305, 2002.
MOLINA, W. F. Chromosomal changes and stasis in marine fish groups, in Fish Cytogenetics, E. Pisano, C. Ozouf-Costaz, F. Foresti, and B. G. Kapoor, Eds., pp. 69– 110, Science Publishers, Enfield, NH, USA, 2007.
MOLINA, W. F. et al. Performance of human immune stimulating agents in the improvement of fish cytogenetic preparations, Genetics and Molecular Research, vol. 9, no. 3, pp.
1807– 1814. 2010.
implications for the high hybridization success, Zoologischer Anzeiger, vol. 253, pp.
137–142, 2013.
NELSON, J.S. Fishes of the World, JohnWiley & Sons, New York, NY, USA, 3rd edition,
2006. 354p.
NIRCHIO, M.; RONDÓN, R. C. and OLIVEIRA. O. Cytogenetic studies in three species of
Lutjanus (Perciformes: Lutjanidae: Lutjaninae) from the Isla Margarita, Venezuela, Neotropical Ichthyology, vol. 6, no. 1, pp. 101–108, 2008.
OJIMA Y. and KOJIMA, K. Chromosomal polymorphisms in Apogonidae fishes,
Proceedings of the Japan Academy B, vol. 61, pp. 79–82, 1985.
ROCHA, L. A. et al. Adult habitat preferences, larval dispersal, and the comparative phylogeography of three Atlantic surgeonfishes (Teleostei: Acanthuridae), Molecular Ecology, vol. 11, no. 2, pp. 243–252, 2002.
ROCHA, L. A. Patterns of distribution and processes of speciation in Brazilian reef fishes,
Journal of Biogeography, vol. 30, no. 8, pp. 1161–1171, 2003.
ROCHA, E. C. and MOLINA, W. F. Cytogenetic analysis in western Atlantic snappers (Perciformes, Lutjanidae), Genetics and Molecular Biology, vol. 31, no. 2, pp. 461–
467, 2008.
ROCCO, L. et al. Molecular and karyological aspects of Batoidea (Chondrichthyes, Elasmobranchi) phylogeny, Gene, vol. 389, no. 1, pp. 80–86, 2007.
SORENSON, L. et al. A multilocus timetree of surgeonfishes (Acanthuridae, Percomorpha), with revised family taxonomy, Molecular Phylogenetics and Evolution, vol. 68, no. 1,
pp. 150–160, 2013.
SENA D. C. S. and MOLINA, W. F. Robertsonian rearrangements and pericentric inversions in Scaridae fish (Perciformes), Genetics and Molecular Research, vol. 6, no. 3 pp.
575–580, 2007.
SENA, D. C. S. and MOLINA, W. F. Chromosomal rearrangements associated with pelagic larval duration in Labridae (Perciformes), Journal of Experimental Marine Biology and Ecology, vol. 353, no. 2, pp. 203–210, 2007.
SUMNER, A. T. A simple technique for demonstrating centromeric heterochromatin,
Experimental Cell Research, vol. 75, no. 1, pp. 304–306, 1972.
TANG, K. L. et al. The phylogenetic relationships of the suborder Acanthuroidei (Teleostei: Perciformes) based on molecular and morphological evidence, Molecular Phylogenetics and Evolution, vol. 11, no. 3, pp. 415–425, 1999.
THODE, G.; GILES, V. and ALVAREZ, M. C. Multiple chromosome polymorphism in
Gobius paganellus (Teleostei, Perciformes), Heredity, vol. 54, no. 1, pp. 3–7, 1985.
5. Capítulo II
Abstract
Acanthurus is a representative and widespread genus of marine fish that plays a key role in
ecological dynamics of coral reefs. Three species are common along coastal reefs of Western Atlantic: A. coeruleus, A. bahianus and A. chirurgus. The cytogenetic pattern of these species
indicated sequential steps of chromosomal rearrangements dating back 19 to 5 millions of years ago (M.a.) that accounted for their interspecific differences. Acanthurus coeruleus
(2n=48; 2sm+4st+42a), A. bahianus (2n=36; 12m+2sm+4st+18a) and A. chirurgus (2n=34;
12m+2sm+4st+16a) share an older set of three chromosomal pairs that were originated through pericentric inversions. A set of six large metacentric pairs formed by Robertsonian (Rb) translocations found in A. bahianus and A. chirurgus and a putative in tandem fusion
found in A. chirurgus are more recent events. In the present study, new cytogenetic data are
being reported for these three Acanthurus species based on mapping of repetitive sequences
such as ribosomal 18S and 5S rDNA and telomeric repeats to improve their karyotype evolutionary analyses. The lack of interstitial telomeric sequences (ITS) in spite of several centric fusions in A. bahianus and A. chirurgus might be related to the long period of time
1. Introduction
The Acanthuridae family (Perciformes) is a monophyletic and relatively ancient group of marine fishes (nearly 54 millions of years ago) with fast evolutionary divergence (CLEMENTS et al., 2003; SORENSON et al., 2013). Nowadays, this family is divided into six genera and comprises about 80 species, popularly known as surgeonfishes (NELSON, 2006).
The genus Acanthurus Forsskål (1775) is one of the mostemblematic groups of
surgeonfishes closely associated to coastal reefs (BELLWOOD and WAINWRIGHT, 2002; NELSON, 2006). This genus is widespread over tropical and subtropical seas, playing a major role in the ecology and evolution of coral reefs due to their herbivory habit (VALENTINE and HECK JR., 1999). The large assemblages of Acanthuridae and other grazers (e.g. parrotfishes and damselfishes) influence the coral reef structure by promoting bioerosion and controlling the spread of macroalgae cover (HORN, 1989; PADDACK et al., 2006).
Four Acanthurus species are recorded in Western Atlantic, being three of them
commonly found throughout brazilian coast; A. coeruleus Bloch and Schneider (1801), A. bahianus Castelnau (1855), and A. chirurgus (BLOCH, 1787) (BERNALL and ROCHA,
2011; MENEZES and FIGUEIREDO, 1985). These three species present highly differentiated karyotypes (2n=48, 36, and 34, respectively) in which Robertsonian rearrangements reduced the diploid values from 2n=48 to 2n=36 in A. bahianus and 2n=34 in A. chirurgus.
In fact, six large metacentric pairs, which are absent in A. coeruleus, were derived
from centric fusions and are apparently homeologous between A. bahianus and A. chirurgus,
corroborating their close phylogenetic relationship. Such remarkable differences in the karyotype macrostructure are rare in marine fishes. Comparing these results to molecular data available for Acanthuridae, it was possible to infer that centric fusions have taken place about 5 M.a. while the pericentric inversions shared by the three species were more ancient events (nearly 19 M.a.) (AFFONSO et al., 2014).
In view of the unusual cytogenetic variation in Atlantic Acanthurus, chromosomal
banding and cytogenetic mapping of ribossomal genes and telomeric sequences were performed in A. coeruleus, A. bahianus and A. chirurgus. Our purpose was to infer if
2. Material and methods
Cytogenetic analyses were carried out in specimens of A. coeruleus (N=23), A. bahianus (N=12) and A. chirurgus (N=17) collected along the shoreline of Natal, Rio Grande
do Norte state (5o46’S, 35o12’W) and Salvador, Bahia state (13°00’S, 38°32’W), northeastern Brazil. Mitotic stimulation was performed by the inoculation of antigen complexes in the specimens (MOLINA, 2001, MOLINA et al., 2010). After 24 h, the animals were anesthetized with crave oil (1 mL/15 L of salt water) and euthanized for removal of anterior kidney. The experiments and euthanasia of specimens were authorized by the Committee of Animal Ethics (CEUA/UESB) from Universidade Estadual do Sudoeste da Bahia (#32/2013).
Chromosomal spreads were obtained by interrupting the cell cycle in vitro (GOLD et
al., 1990). The active nucleolar organizer regions (NORs) were detected by silver nitrate staining (HOWELL and BLACK, 1980). Once heterochromatin patterns have been previously reported in the analyzed species (AFFONSO et al., 2014), chromosomes were stained with base-specific fluorochromes to locate GC- and AT-rich sites by mithramycin (MM) and 4’-6-diamino-2-fenilindole (DAPI), respectively (SCHWEIZER, 1980).
FISH was performed according to Pinkel et al. (1986) using 18S, 5S and telomeric probes. The probes of 5S rDNA (nearly 200 bp) and 18S rDNA (nearly 1400 bp) were obtained from DNA samples of A. coeruleus via PCR using the primers A 5’-TAC GCC
CGA TCT CGT CCG ATC-3’/B 5’-CAG GCT GGT ATG GCC GTA AGC-3’ (Pendás et al.,
1994) and NS1 5’-GTA GTC ATA TGC TTG TCT C-3’/NS8 5’-TCC GCA GGT TCA CCT ACG GA-3’ (WHITE et al., 1990), respectively. Both probes were labeled by nick translation (Roche, Mannheim, Germany). The 5S rDNA probes were labeled with biotin-14-dATP (Invitrogen) and detected with avidin-FITC (Sigma); while the 18S rDNA probes were labeled with digoxigenin-11-dUTP (Roche) and detected with anti-digoxigenin-rhodamin (Roche). The (TTAGGG)n sequences were mapped by FISH using Telomere PNA FISH Kit/FITC according to manufacturer’s instructions (DakoCitomation).