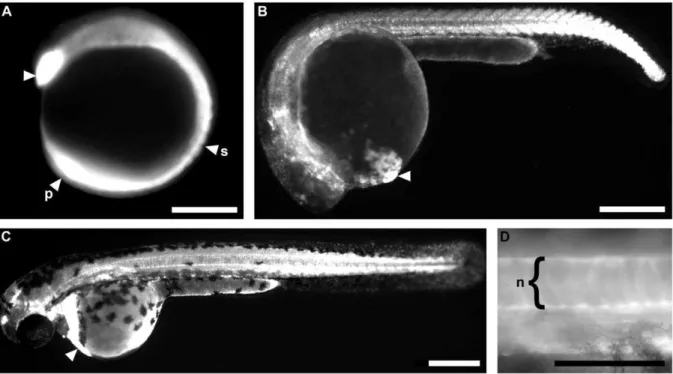
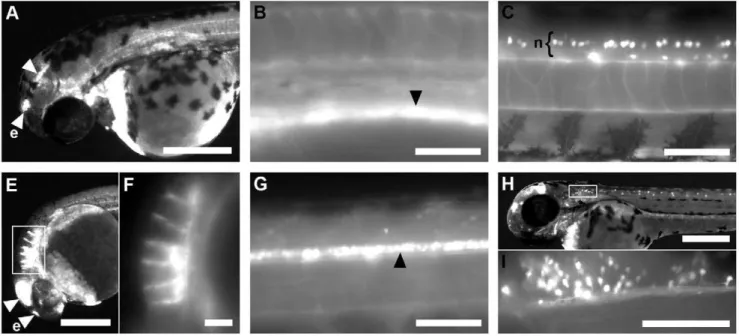
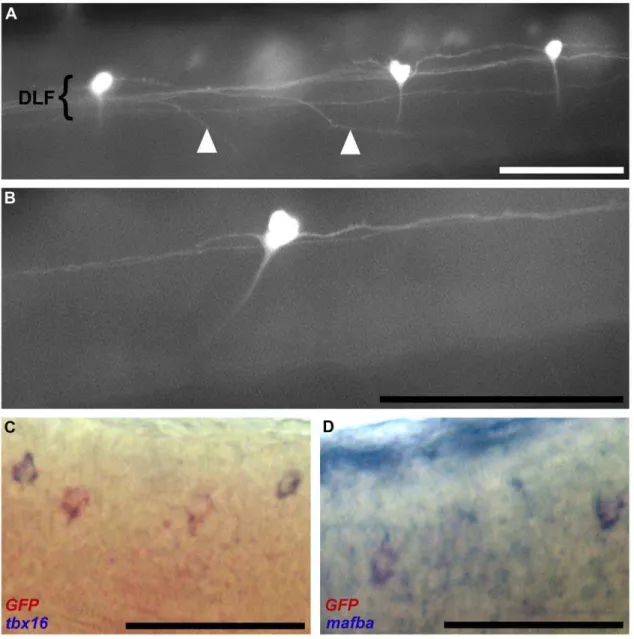
Transgenic zebrafish recapitulating tbx16 gene early developmental expression.
Texto
Imagem

Documentos relacionados
In our experiments, the G9 promoter fused to the reporter gene uidA showed maximal expression at stage 10 in transgenic tobacco (Figure 4), coinciding with the transcriptional
Enhancement of the methionine content of seed protein by the expression of a chimeric gene encoding a methionine-rich pro- tein in transgenic plants.. Accumulation
Furthermore, gene expression profiling of Oct4-gfp + /cKit bright cells from the Oct4-gfp line and SSEA1 + /cKit bright cells from J1 EBs at day 6 revealed enriched expression of
In recombinant mapping and transgenic lines ’ assays, the viral titre or gene expression in resistant flies relative to susceptible flies was calculated as 2 - ΔΔ Ct , where ΔΔCt
To understand the expression of Oct4 in OSCC cell lines (OSCCs), the endogenous protein level of Oct4 in nine established OSCC cell lines and one normal oral epithelial cell line SG
For example, transgenic line (FHZ.03, Figure 3) had an insertion into the third intron of Npas3, a gene mutated in rare forms of familial schizophrenia [13]. The expression of
Comparative analysis of the expression levels of OsDR10 in different rice lines showed that this gene had a similar expression level in the three resistant rice lines (Figure
Expression of the GFP-tagged wild type version of the regulatory light chain, RLCT20S21-GFP, in cells depleted of the endogenous RLC rescued the cytokinesis defect (Figure 2B),
