RICARDO ALMEIDA EMIDIO
Otimização no uso de martelos e bigornas para quebrar sementes
por macacos prego (Cebus flavius e C. libidinosus) no Bioma Caatinga.
Dissertação apresentada à Universidade Federal do Rio Grande do Norte, para obtenção do título de Mestre em Psicobiologia
RICARDO ALMEIDA EMIDIO
Otimização no uso de martelos e bigornas para quebrar sementes
por macacos prego (Cebus flavius e C. libidinosus) no Bioma Caatinga.
Dissertação apresentada à Universidade Federal do Rio Grande do Norte, para obtenção do título de Mestre em Psicobiologia
Orientadora: Renata Gonçalves Ferreira
Catalogação da Publicação na Fonte. UFRN / Biblioteca Setorial do Centro de Biociências Emidio, Ricardo Almeida.
Otimização no uso de martelos e bigornas para quebrar sementes por macacos prego (Cebus flavius e C. libidinosus) no Bioma Caatinga/ Ricardo Almeida Emidio. – Natal, RN, 2010.
60 f. : Il.
Orientadora: Profª. Renata Gonçalves Ferreira.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Programa de Pós-Graduação em Psicobiologia.
1. Macaco pregos – Dissertação. 2. Caatinga – Dissertação. 3. Estratégia de forrageiro – Dissertação. I. Ferreira, Renata Gonçalves. II. Universidade Federal do Rio Grande do Norte. III. Título.
Título: Otimização no uso de martelos e bigornas para quebrar sementes
por macacos prego (Cebus flavius e C. libidinosus) no Bioma Caatinga.
Autor: Ricardo Almeida Emidio
Data da defesa:
Banca Examinadora:
___________________________________ Profa. Maria Adélia Borstelmann de Oliveira Universidade Federal Rural de Pernambuco, PE
___________________________________ Profa. Fívia de Araújo Lopes
Universidade Federal do Rio Grande do Norte, RN
___________________________________ Profa. Renata Golçalves Ferreira
AGRADECIMENTOS
Agradeço,
Aos meus pais, o alicerce na minha vida, sou grato ao amor e dedicação incondicional. Sem dúvida, eles abdicaram do conflito pais-prole (hehe ).
A Daniela K. Menezes (futuramente DKM Emidio) por ter acompanhado, apoiado, vivido, dado suporte e acima de tudo por me amar, para mim, isso já basta ;).
Ao Elmo, dona Gláucia e o Rafael, membros da família da minha noiva e, porque não minha também? Sou grato por me oferecerem amor de filho e de irmão, pelos conselhos, observações e pelo total apoio. Vocês são muito importantes para mim!
A minha inestimável e querida professora Renata Gonçalves Ferreira pela incrível orientação, pelos incentivos e puxões de orelha no decorrer do trabalho. Orientar um hiperativo tem os seus desafios... Também pelo pleito da bolsa ao CNPq, sem ela não este trabalho não teria sido realizado.
A todos os professores da Psicobiologia pela dedicação que faz deste programa de pós-graduação em ciências do comportamento o melhor do Nordeste. Em especial, a professora Fátima Arruda pelos seus conselhos de como otimizar o trabalho, muito obrigado!
Ao CNPq (processo nº 564275/20085) pela bolsa de mestrado – que pena, ela já foi – e pela oportunidade de dizer ao professor Arrilton: “Na sua disciplina posso tirar até Z, porque não perco a bolsa!”.
Aos meus amigos e colegas da PPg Psicobiologia, em especial ao Tiago Eugênio (Kutako), Luciane Coletti e Felipe Nalon pelas boas conversas, bons momentos e discussões sobre os nossos trabalhos.
A minha querida e amada professora Maria Adélia Borstelmann de Oliveira pelas boas palavras e oportunidade de iniciar os estudos de comportamento.
A todos que de alguma forma ajudaram na parte logística do trabalho. Em especial, a Lídia Brasileira pelo gerenciamento da RPPN Stossel Britto (Jucurutu, Rio Grande do Norte). Sou grato ao Alan por identificar algumas plantas e pela total disponibilidade em cooperar. Agradeço ao IDEMA por fornecer as imagens de satélite apresentadas neste trabalho. Também sou grato aos meus queridos mateiros (Galego de Zeca, Chagas, Formiga, Seu Caboclo, Neto e Pitota) pela companhia e auxílio na coleta de dados. Não posso esquecer de agradecer as ótimas cozinheiras (Amailda e dona Carmem). Afinal, comida pronta após um dia duro de trabalho não tem preço!!
A banca avaliadora por melhorar o meu trabalho.
SUMÁRIO
RESUMO ... 1
ABSTRACT ... 2
INTRODUÇÃO ... 3
O uso de instrumentos e o aumento da razão neocortical ... 3
Utilização de ferramentas – uma visão geral ... 6
O uso de ferramentas entre os primatas viventes ... 8
Macacos prego como modelos de estudo ... 10
Características do genero Cebus ... 10
Uso de instrumentos por macacos-prego ... 12
Otimização no uso de instrumentos ... 19
MÉTODO ... 24
Área de estudo ... 24
Animais de estudo ... 26
Esforço amostral ... 27
Registro metodológico ... 27
Parcelas monitoradas... 27
Fenologia das espécies vegetais produtoras de sementes e frutos cultivares ... 28
Definição e registro de presença de sítio de quebra ... 33
Registro do peso dos martelos ... 34
Preparação do banco de dados geográficos: Distância mais próxima entre pontos ... 35
Análise de dados ... 35
RESULTADOS ... 36
Visão geral ... 36
Escolha dos martelos para a quebra de sementes ... 36
Escolha espacial do sítio de quebra... 42
DISCUSSÃO ... 45
Escolha do martelo – eficiência na quebra de sementes ... 46
Escolha das bigornas – estratégias para reduzir riscos de predação e custos de competição direta. ... 48
CONCLUSÕES ... 52
INDÍCE DE FIGURAS
Figura 1 - Áreas de estudo contendo mapa geral (A), área de estudo de Jucurutu (B) e área de estudo de Luís Gomes (C). Maiores detalhes conferir a legenda do mapa acima. ... 30
Figura 2 – Sazonalidade nas áreas de estudo. Figuras a-b = Jucurutu (Caatinga arbustiva aberta) estação chuvosa e estação seca, figuras c-d = Luís Gomes (Caatinga
arbóreo-arbustiva) estação chuvosa e estação seca. ... 31
Figura 3 – Exemplos de bigornas e martelos usados nos sítios de quebra. Figuras: a = representa um sítio de quebra de semente de maniçoba (Manihot dichotoma), b = sítio de quebra com bigornas grandes (> 1m), c = zoom da bigorna, d = imagem de martelos utilizados em sítios de quebra de maniçoba, e = sítios de quebra ativos de Syagrus cearensis. ... 32
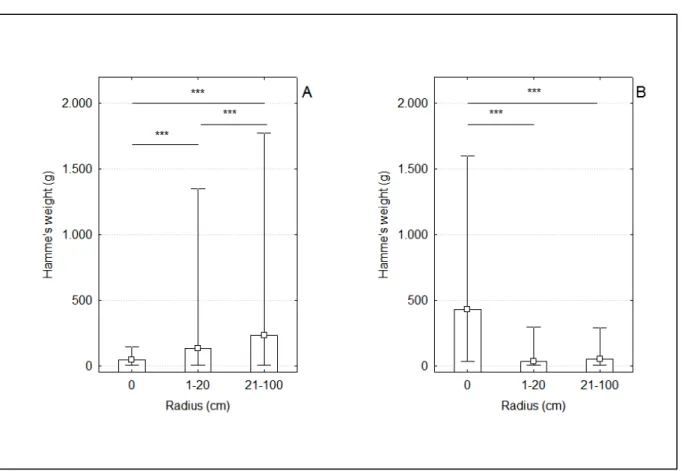
Figura 4 – Análises estatísticas correspondentes à predição 1. (A) Caatinga arbustiva aberta: massa das pedras encontradas sobre a bigorna (0 cm, N = 414); entre 1-20 cm da bigorna (N =162) e entre 21-100 cm da bigorna (N=363). (B) Caatinga arbóreo-arbustiva: massa das pedras encontradas sobre a bigorna (0 cm, N = 93); entre 1-20 cm da bigorna (N =115) e entre 21-100 cm da bigorna (N=346). Mediana, Distribuição não outlier dos dados, *** p < 0,001. ... 37
Figura 5 – Análises estatísticas correspondentes às predições 2 e 3. (A) Massa dos martelos utilizados para quebrar sementes por macacos prego em ambos os tipos de Caatinga. M. dichotoma1 (N= 414); M. dichotoma2 (N= 13); Mix2 (Sítios de quebra mistos, N = 4) e
Figura 6 – Sementes consumidas pelos macacos prego e indicadores de estágios de
maturação. (A-D) Coco catolé (Syagrus cearensis), (E-F) Maniçoba (Manihot dichotoma). (A&E) Estágio verde. (B&F) Semente quase madura. (C&G) Estado maduro. (D) Estágio podre. ... 40
Figura 7 – Análises estatísticas correspondentes às predições 4 e 5. (A) Distâncias entre as espécies vegetais produtoras de sementes nas áreas de estudo (Distance1). Manihot
dichotoma1 (N =1306); M. dichotoma2 (N = 146); Syagrus cearensis2 (N = 233). (B) Distâncias entre os sítios de quebra e as respectivas espécies produtoras de sementes em ambas as áreas de estudo (Distance2). Manihot dichotoma1 (N =2192); M. dichotoma2 (N = 50); Syagrus cearensis2 (N = 412). (C) Distâncias entre os sítios de quebra (Distance3) para cada espécie de semente em áreas de estudo. Manihot dichotoma1 (N =1912); M. dichotoma2 (N = 88); Syagrus cearensis2 (N = 380). (D) Distâncias entre os sítios de quebra (Distance3) em ambas as áreas de estudo. Caatinga arbustiva aberta (N =2192), Caatinga arbustivo-arbórea (N = 462). medianas e distribuição não outlier dos dados; *** = p < 0,001; Manihot dichotoma1 = Caatinga arbustiva aberta; Manihot dichotoma2 e Syagrus cearensis2 = Caatinga arbustivo-arbórea. ... 43
INDÍCE DE TABELAS
Tabela 1 – Compilação dos resultados estatísticos. ... 41
RESUMO
ABSTRACT
INTRODUÇÃO
O uso de instrumentos e o aumento da razão neocortical
Duas pressões ambientais são apresentadas para explicar o aumento da razão neocortical nos hominídeos: habilidades na aquisição de alimento (teorias dos mapas cognitivos e uso de instrumentos) e relacionamento social entre indivíduos (teoria da inteligência maquiavélica).
A teoria de mapas cognitivos proposta por Milton (1981) argumenta que para encontrar alimentos é preciso aprendizado dos itens desejáveis adquiridos dentro da sua área de uso em estações sazonais apropriadas. Para que este processo ocorra é necessária uma boa memória, que seja capaz de armazenar informações sobre locais onde há fontes de alimento e em que época, assim como direcionar sua rota. Ou seja, o aumento da razão neocortical deve-se a melhor relação custo/benefício na aquisição de nutrientes e os animais com maior neocortex seriam capazes de utilizar rotas com possibilidade de encontro de alimentos de alta quantidade e qualidade além do melhor investimento de forrageio.
Entretanto, outros fatores relacionados à alimentação têm sido sugeridos como pressão para aumento da razão neocortical, referindo-se não ao encontro do alimento, mas à obtenção do mesmo. Entre estes fatores destaca-se a habilidade de uso de instrumentos na coleta e processamento de itens encapsulados.
Em espécies de primatas que possuem adaptações morfológicas para a obtenção de itens encapsulados não é registrado o uso de instrumentos em condições naturais. Por exemplo, o sagui do nordeste (Callitrhix jacchus), uma espécie de primata neotropical possui adaptações morfológicas e filogenéticas para a obtenção de itens encapsulados (Feagle, 1998). Com os incisivos inferiores longos, estes primatas conseguem roer troncos e posteriormente adquirir goma. Analogamente poderíamos afirmar que nestes primatas, os dentes da arcada inferior atuam como ferramentas naturais e eficientes na obtenção deste tipo de alimento.
A terceira teoria para explicar o aumento da razão neocortical em primatas e hominídeos é a inteligência social (ou inteligência maquiavélica). Dentre as suas várias versões, esta teoria agrega a proposta de que as interações sociais atuam como pressão para o processo de adaptação e complexidade da inteligência dos hominídeos (Byrne & Whiten, 1997).
Humphrey (1976) em seu manuscrito que tornou-se um dos fundamentos teóricos para o surgimento da teoria da inteligência maquiavélica, sugeriu que, ao contrário do que se imagina, intelecto está relacionado principalmente às relações sociais, contextos mais freqüentes e que necessitam de maior atenção para a sobrevivência dos indivíduos do que tentar resolver problemas matemáticos, abstratos por natureza, e não contextualizados com hábito de vida dos animais.
Estratégias comportamentais direcionadas a manipulação e dissimulação social são observadas sobre a ótica da Inteligência maquiavélica. Byrne (1995) afirma que estes comportamentos são apresentados somente entre os haplorríneos, espécies de um grupo taxonômico que adotam dissimulações frequentemente, e, aparentemente aprendidas por tentativa e erro.
espaço-tempo, torna-se plausível afirmar que cada pressão atuou com diferentes intensidades e interagiu com outras diferentes forças entre diversas espécies (Byrne, 1995)
Utilização de ferramentas – uma visão geral
Neste trabalho utilizamos o conceito de uso de instrumentos definido por Amant & Horton (2008, pp 1203) “Uso de instrumentos é a execução do controle sobre uma manipulação livre de um objeto externo (a ferramenta) com o objetivo de (1) alterar as propriedades físicas de outro objeto, substâncias, superfícies ou o centro ( o alvo, como pode ser o próprio usuário de ferramentas ou outro organismo) via uma interação dinâmica mecânica, ou (2) mediante o fluxo de informações entre o usuário da ferramenta e do ambiente ou de outros organismos no ambiente ”
previamente cativos e reintroduzidos no habitat natural ou semi-natural utilizam ferramentas, que podem ser fruto do aprendizado com humanos.
assim indivíduos juvenis com e sem tutores apresentaram taxas similares de sucesso nas tarefas de uso de instrumentos.
Há descrições de que golfinhos selvagens (Tursopis sp.) usam instrumentos. Estes animais usam esponjas em atividades de forrageio, e há um forte viés quanto ao sexo, deste modo, somente fêmeas usam instrumentos (Krützen et al., 2005). Krützen et al., (2005) observaram maiores semelhanças genéticas entre mães que usam instrumentos do que aquelas que não usam instrumentos, sugerindo assim um forte componente genético na manutenção do uso de instrumentos.
O uso de ferramentas entre os primatas viventes
Dentre os primatas viventes, os chimpanzés (Pan troglodytes) são os que apresentam o repertório comportamental mais complexo de uso de instrumentos, tanto em condições de cativeiro, que favorecem o uso de ferramentas, quanto em vida livre. McGrew (1992) faz um compilado da complexidade de uso de ferramentas desses animais como o uso de martelos (de madeira e pedra) para quebrar sementes, esponjas para absorver água, troncos de árvores como armas contra co-especificos ou predadores. Há relatos da escolha de diversas matérias-primas para a construção de ferramentas que promovam os mesmos fins, como galhos ou troncos para construir iscas de capturas de insetos sociais. Também estes primatas usam o mesmo material para construir diversos instrumentos ao exemplo de usar folhas como esponjas, para atividades de limpeza e investigação. Diante dessa diversidade e singularidade no uso de instrumentos em algumas populações, McGrew (1992) sugere que estas diferenças são dependentes de uma condição cultural e não de variações dependentes das condições ecológicas.
instrumentos, como a quebra de galhos que serviam como pontes entre árvores, e o uso efetivo de instrumentos. Dentre o uso de instrumentos, foi observado a emissão de sons em árvores, a produção de pontes que consistia no uso do próprio peso do corpo para deslocar-se entre árvores e o uso de folhas como função de coberturas contra chuva. O uso de ferramentas em Pan paniscus está fortemente associado a contextos sociais, diferentemente do observado em Pan troglodytes (McGrew, 1992). Esta autora sugere que, em bonobos, a tecnologia e socialidade caminham juntos, deste modo, ao passo que os animais manipulam objetos e usam instrumentos também acabam manipulando socialmente co-específicos. A autora conclui que a similaridade no entendimento intelectual das habilidades entre os grandes primatas são expressas de formas diferentes a depender do requerimento das particularidades ecológicas e sociais.
Acreditava-se que orangotangos (Pongo pygmaeus) não usariam instrumentos na natureza, apesar do seu uso comum em situações de cativeiro (McGrew, 1992). Porém van Shaink & Fox (1996) relataram a criação e o uso de instrumentos em animais cativos que foram soltos no ambiente natural. Os orangotangos foram capazes de selecionar galhos para capturar insetos sociais, modificaram galhos de árvores para acessar sementes de Neesia sp. Diante destes resultados, os autores indicam que o uso de instrumentos não está limitado apenas ao clado homem-chimpanzé sugerindo que o uso de instrumentos também está presente nos orangotangos e nos hominídeos africanos.
floresta do Congo e foi verificado que uma fêmea estava deslocando-se através de um lago então coletou um tronco que foi utilizado para medir a profundidade da lagoa. Em outra observação, outra fêmea utilizou um tronco para coletar plantas aquáticas encontradas na borda de um lago. Estes autores sugerem que o uso de instrumentos nestes animais esteja relacionado fortemente a aquisição de rotas e que é mais um fator comparativo de uso de instrumentos nos primatas do velho mundo.
Não há relato de instrumentos tanto em cativeiro quanto em vida livre em gibões (Hylobates sp.), apesar de estudos de mais de 50 anos na região sudoeste da África, não foi observado o uso de ferramentas em vida livre (McGrew, 1992).
Macacos prego como modelos de estudo
Dentre os primatas neotropicais, os macacos prego (Cebus spp.) vêm sendo utilizados como modelo para o entendimento das três teorias que buscam explicar como ocorreu o aumento do neocórtex (mapas cognitivos, uso de instrumentos e inteligência maquiavélica). Exibindo um alto grau de enefalização (2,21; tabela 1 de Kudo & Dunbar, 2001), este gênero é conhecido por apresentar mapas mentais das suas áreas de vida (Presotto & Izar 2010), apresenta estratégias sócias de formação de coalizões (Perry et al., 2003; Ferreira 2003; Ferreira et al., 2006) e recentemente vem sendo alvo de diversos estudos acerca do uso de instrumentos em ambientes com restrição de alimento (ver revisão em Ottoni & Izar, 2008).
Características do genero Cebus
destreza manual. Os macacos prego vivem em grupos sociais entre oito a 40 individuos apresentando muitos machos adultos, femeas adultas e infantes. Eles possuem grande dimorfismo sexual (machos até 30% maiores que as femeas, a dependeder da espécie) e hierarquia de dominancia entre machos, sendo mais despotica em C. apella e menos despotica em C. albrifrons. Macacos prego tem grandes áreas de uso (até 600ha na amazônia) e deslocamentos diários de cerca de 3km (Feagle, 1998; Ferreira, 2003).
O genero Cebus é considerado o grupo taxonomico de primatas neotropicias muito divergente (Silva Jr. 2001). Uma classificação recente divide o gênero em dois subgêneros (Sapajus e Cebus, respectivamente – Silva Jr., 2001). O subgênero sem tufo, Cebus, foi dividido em quatro espécies: Cebus capuchinus, C.albrifons, C. olivaceus e C. kaapori. O subgenero com tufo, Sapajus, foi dividido em oito espécies: Cebus apella, C. macrocephalus, C. lidinosus, C. cay, C. xanthosternos, C. robustos, C. nigritus (Silva Jr., 2001) e C. flavius (Oliveira & Langguth, 2006).
Macacos pregos são conhecidos por possuir uma dieta onivora, assim ampliando suas possibilidades de ingestão de alimento. Estes animais alimentam-se desde frutos, sementes, castanhas, flores, goma, néctar, fungos, seiva, ovos, invertebrados, pequenos vertebrados e algumas espécies de invertebrados habitantes de ambientes de manguezais (Fragaszy et al., 2004a). Estas características fazem que este gênero possua uma das maiores capacidades de habitar uma diversidade de ambientes bem contrastantes.
Amazônica (Port-Carvalho et al., 2003), regiões de manguezais (Fernandes, 1991) e também encontrados na América Central (Phillips, 1995a, b; Hall & Fedigan, 1997).
Dentro da estrutura espacial de grupos de macacos prego, há formação de subgrupos que variam de acordo com as características e as estratégias sociais dos indivíduos que balanceiam os custos (risco de predação e competição por alimento) e benefícios (defesa contra predadores e aquisição de alimento) de viver em grupo. Janson (1990), Hall & Fedigan (1997) relatam com detalhes a ocorrência destas escolhas espaciais nas formações destes subgrupos nas tarefas de forrageio: machos e fêmeas dominantes e infantes no centro, juvenis com livre acesso, além dos animais periféricos como machos e fêmeas adultos subordinados que evitavam o centro (e, consequentemente, os animais dominantes) e machos e fêmeas subadultos que posicionavam-se na frente do grupo.
As agressões podem interferir significativamente na taxa de sucesso de forrageio (Janson, 1985). Em macacos pregos, grande parte das agressões ocorre em atividades de forrageio, logo,indivíduos que recebem menor número de agressões forrageiam melhor do que aqueles que sofrem maior incidência de injúrias (Janson, 1985). Assim, os animais tendem a manter-se afastados para reduzir estas taxas de agressão e, consequentemente,aumentarem o sucesso de forrageio (Janson, 1990; Phillips 1995a, b).
Uso de instrumentos por macacos-prego
instrumentos para alcançar alimentos fora do alcance com objetos, quebrar sementes com o auxílio de martelos, absorver líquido com papel toalha e usar galhos e pedras para cortar um revestimento em vários pedaços. Em outros estudos são descritos comportamentos de modificação ou contrução de ferramentas, como amassar papéis toalha para servir como esponjas. A produção de farpas em rochas para cortar couro e o uso de bambú para produzir brechas em tubos plásticos.
Fernandes (1991) promoveu o primeiro registro de uso de instrumentos em macacos prego em vida livre. Entre o seu resgistro até o ano de 2000, este comportamento era considerado como esporádico, provavelmente pela falta de estudos mais longos em ambientes propícios. A partir de 2004, o interesse pelo uso de ferramentas por Cebus spp. tornou-se destaque na ciência, e, em seis anos mais de dois artigos por ano foram publicados sobre essa temática. Nos manuscritos publicados até o presente período, tivemos a oportunidade de observar por novos ângulos, estratégias comportamentais associadas ao uso de instrumentos, sendo este complexo e intrigante.
Em região de manguezal no estado do Maranhão, Fernandes (1991) observou estes animais (Cebus apella), utilizando pedaços de conchas de (Crassostrea rhizophorae) para as atividades de abertura e retirada das ostras da mesma espécie. Ainda, tal autor sugeriu que a capacidade de alimentar-se de uma variedade de potenciais recursos atua como componente fundamental para a sobrevivência e permanência de macacos prego em ambientes de mangue.
mastigadas além do uso de ferramentas para quebrar sementes de maniçoba (Manihot sp.), coco catolé (Syagrus oleracea). Apesar dos autores não observarem in loco a quebra de sementes pelos macacos prego, eles encontraram sítios de quebra de coco catolé com martelos pesando aproximadamente um quilo.
Em trabalho pioneiro, Ottoni & Mannu (2001) fizeram a primeira descrição comportamental de quebra de sementes de Syagrussp. com a utilizacão de pedras que serviam com função de martelos e bigornas por macacos prego semi-cativos (Cebus apella) no Parque Ecológico do Tietê, São Paulo. ) Neste trabalho, ocorreu o importante relato de que animais tolerados (infantes e juvenis) frequentemente roubavam amêndoas de animais adultos, estes autores questionaram acerca da importância da manutenção deste comportamento num grupo social. Posteriormente, em 2005 Ottoni e colaboradores responderam tais questionamentos (ver abaixo).
Fragaszy et al., (2004a) estudaram os macacos pregos (C. libininosus) no limite dos biomas Caatinga-Cerrado do estado do Piauí, utilizando as seguintes metodologias: a) observações diretas; e b) busca de evidências físicas de quebra de sementes. Estes autores encontraram os seguintes resultados: i) a identificação direta da utilização de martelos e bigornas na quebra de sementes de Syagrus spp e Atallea spp. , ii) a constatação de depressões pouco profundas nas superfícies das bigornas; iii.) a presença de lascas e destroços de cascas de sementes; e iv.) a presença de pedras soltas com peso apropriado para a quebra de sementes em cima das bigornas.
Ottoni et al., (2005) decidiram investigar os questionamentos levantados por Ottoni & Mannu (2001) e encontraram uma intensa relação custo/benefício entre os eventos de roubos de amêndoas e a facilitação da aquisição de componentes energéticos de difícil acesso associados ao aprendizado da quebra de sementes por observação. Ottoni et al., (2005) sugeriram que este padrão é um importante componente para a permanência do uso de instrumentos neste grupo.
Waga et al.,(2006) estudaram macacos pregos (C. libidinosus) no Parque Nacional de Brasília, região representativa do Cerrado brasileiro. Utilizaram amostras comportamentais diretas e observações de vestígios, encontrando os seguintes resultados: i.) episódios de utilização de ferramentas por macacos pregos e ii) utilização de ferramentas por dois anos consecutivos evidenciando o uso contínuo deste comportamento nos grupos monitorados.
que isto, os dados indicam escolhas ativas dos animais, tanto para a utilização de bigornas quanto para utilização dos martelos. Foi verificada uma distribuição não randômica das bigornas, de forma que estas se localizavam próximas às árvores, mas não exatamente as produtoras de sementes. Já os martelos eram derivados de acidentes rochosos em maior freqüência do que as rochas sedimentares. Os martelos mais pesados foram encontrados próximos às bigornas, o que evidencia que os animais transportavam tantos as sementes quanto as ferramentas (os martelos) para os sítios de quebra.
Silva (2008) relatou as estratégias comportamentais relacionadas ao roubo e partilha de frutos encapsulados por macacos pregos (C. libidinosus). Este autor encontrou resultados que corroboram com os achados de Ottoni et al., (2005) e sugeriu, ainda, que as estratégias adotadas para observar animais quebrando sementes pode variar de acordo com o tamanho da semente. Segundo Silva (2008), quanto maior o tamanho da semente, maior era a possibilidade do animal aproximar-se para observar e usurpar sementes quebradas e ao mesmo tempo poderia ocorrer maior tolerância com sementes maiores do que sementes menores, mas apenas para infantes e juvenis.
cheirar e ou esfregar várias partes do corpo como a face, o peito e as mãos. Ainda não se sabe a função deste uso do pó. Em várias ocasiões foram registradas partes de plantas utilizadas como ferramentas. Os animais utilizavam os pedaços de galhos para inspecionar troncos de árvores quebrados anteriormente por martelos e obtiveram sucesso superior a 70% das tentativas. Os autores sugerem que as variações no uso de ferramentas em Boa Vista e Serra da Capivara (Piauí) podem ser fruto de tradições locais, proposta defendida pela observação da ontogenia do uso de ferramentas em grupos provisionados no parque do Tietê, São Paulo.
Canale et al., (2009) fizeram o primeiro registro de uso de instrumentos por C. xantosternos e adicionou novas áreas de ocorrência por C. libidinosus, nos Biomas de Caatinga, Cerrado e seu ecótono entre os estados de Sergipe, Bahia e Minas Gerais. Estes autores encontraram a quebra de sementes associada a espécies vegetais como Syagrus spp., Atallea spp. e Cnidoscolus sp. Através destes dados os autores sugerem que o uso de instrumentos é uma característica dos biomas Caatinga e Cerrado, onde ocorre limitação de água e alimento ou consideram uma característica intrínseca do gênero Cebus.
Emidio & Ferreira (2009) foram os primeiros a descrever o uso de instrumentos por C. flavius no bioma Caatinga no estado do Rio Grande do Norte. Tais autores registraram sementes de maniçoba (Manihot dichotoma) associadas a sítios de quebra e o uso de instrumentos para desenterrar tubérculos de maniçoba.
Em situação não experimental ou induzida, Ferreira et al., (2010) confirmaram a sugestão de escolhas inteligentes de martelos ao quebrar sementes propostas por Visalberghi et al., (2009a). Os autores promoveram o primeiro relato de escolhas ativas de martelos por macacos prego de acordo com a espécie de semente. Assim, pedras mais leves foram destinadas para quebrar sementes menos resistentes e pedras mais pesadas utilizadas para quebrar sementes mais resistentes, algo anteriormente somente observado em chimpanzés por Boesch & Boesch, (1983).
Visalberghi et al., (2009b) apresentaram um panorama das características ecológicas do ecótono Caatinga-Cerrado na região da Boa Vista e descreveram que as bigornas e plantas produtoras de sementes são encontradas com maior abundância em regiões de penhascos. Tais autores descreveram que, devido a baixa disponibilidade de pedras adequadas em regiões onde localizam-se as plantas produtoras das sementes encapsuladas, os animais realizam o transporte de sementes e martelos para as bigornas. Estes achados são interpretados promovendo um comparativo com o observado em chimpanzés por Boesch & Boesch (1983) e Sakura & Matsuzawa (1991).
Por fim, Fragaszy et al., (2010) observaram que macacos prego com características intrínsecas diferentes, tais como peso do corpo e experiências anteriores, podem gerar diferentes balanços energéticos da quebra de sementes. Ou seja, machos adultos são os animais mais proficientes na quebra de sementes entre os membros do grupo (Silva 2008; Fragaszy et al., 2010). Também foi observado que assim como humanos, os macacos preferem quebrar sementes em locais em que há pequenas depressões para reduzir os custos de quebra de sementes.
martelos para diversas atividades, desde a quebra de sementes a produção de pó com pedras para passar na face dos indivíduos (Mannu & Ottoni, 2008); ii) transporte de martelos e sementes para os sítios de quebra em áreas em que estes não são comuns (Visalberghi et al., 2007; Visalberghi et al., 2009b); iii) capacidade de escolher martelos para quebrar diversas espécies de sementes (Ferreira et al., 2010); iv) seleção de porções de uma determinada bigorna a ser utilizada na quebra de sementes (Fragaszy et al., 2010); v) diferenças entre os indivíduos da proficiência na execução do uso de instrumentos (Ottoni & Mannu, 2001; Ottoni et al., 2005; Silva, 2008; Fragaszy et al., 2010).
Entretanto, o número de grupos estudados ainda é pequeno, e a maior parte dos resultados derivam de três grupos (Tietê, Boa Vista e Serra da Capivara). Diversos aspectos ainda precisam ser elucidados, principalmente para verificar o quanto o uso inteligente de instrumentos varia a depender de condições ecológicas diferentes (possibilidade de terrestrialidade, disponibilidade de alimentos, presença de predadores e rotas de fuga, disponibilidade de material adequado) ou ainda o quanto o uso de pedras como instrumento deriva de tradições locais e aprendizagem social ou derivam de convergência comportamental devido a similaridade ambiental (Perry et al., 2003).
Otimização no uso de instrumentos
estas bigornas e dispor as sementes sobre locais planos, além de lançar martelos sobre as sementes (Visalberghi et al., 2009b).
O uso ótimo de pedras para quebra de sementes foi abordado pela primeira vez por Boesch & Boesch (1983) através de observações diretas e de vestígios em grupos não habituados de chimpanzés (Pan troglodytes) no Parque Nacional Tai, Costa do Marfim. Os autores identificaram um padrão complexo da quebra de sementes: os chimpanzés utilizavam martelos com massa inferior a três quilos para quebrar sementes menos resistentes (Coula edulis, Olacaceae), enquanto que as sementes de palmeiras que eram mais resistentes (Panda oleosa, Pandaceae) frequentemente eram abertas com martelos com massas que variavam entre três e 9 kg ou superiores a 9 kg. Boesch & Boesch (1983) também mostraram que chimpanzés transportaram pedras mais pesadas mais frequentemente, e por maiores distâncias, que pedras mais leves ou que martelos de madeira, sugerindo um planejamento da ação de quebra de sementes. Por outro lado, Sakura & Matsuzawa (1991) estudando chimpanzés em Bossou relataram a escassez dos eventos de transporte de martelos, pois nesta área havia abundância de material adequado para quebra de sementes.
Em macacos prego, dois diferentes estudos experimentais investigaram a importância da massa na escolha de martelos. O primeiro revelou que macacos prego cativos faziam escolhas de martelos mais pesados para quebrar as sementes de palmeiras difíceis de serem quebradas (Schrauf et al., 2008). O segundo descreveu que os macacos prego provisionados observados na Fazenda Boa Vista (Piauí, Brasil) promoviam escolhas racionais de martelos artificiais (oferecidos pelos experimentadores) e selecionavam aqueles que possibilitam maior efetividade na quebra de sementes de palmeiras (Visalberghi et al., 2009a).
utilizadas por macacos prego não habituados (Rio Grande do Norte, Brasil) e relataram que três diferentes populações de macacos prego (C. libidinosus) em situação não experimental nem induzida escolhiam martelos em função do tamanho das sementes alvo. Assim, foram verificadas que pedras mais pesadas associadas a sementes maiores e mais resistentes, enquanto martelos significativamente mais leves foram observados associados a sementes menores.
Em chimpanzés, também foi observado escolhas otimizadas de bigornas. Boesch & Boesch (1983) verificaram que chimpanzés (Pan troglodytes) quebravam sementes maiores em bigornas mais resistentes, enquanto que para as sementes menores não havia preferência quanto ao tipo de bigorna. Visaberghi et al., (2007) descrevem que as bigornas ativas localizavam-se significativamente mais próximas de rotas de fuga (árvores) apesar da presença de diversas outras potenciais bigornas. Segundo este trabalho, a escolha da bigorna reflete uma otimização entre quebrar sementes no solo e a possibilidade de escape aos predadores. Ambos os estudos relatam uso repetido de bigornas.
de martelos deve ser analisada tendo em vista a ecologia do local, e sugere que o transporte e escolha de martelos deve ocorrer em situações especificas, quando há baixa disponibilidade de material adequado.
Diante da falta de informações acerca da otimização no uso de instrumentos por macacos prego que podem ser representadas através das escolhas eficientes de martelos e bigornas em diversos aspectos (ex: peso dos martelos, espécie de semente a ser quebrada e distribuição espacial de bigornas), decidimos testar as seguintes hipóteses e predições:
H1: Macacos prego selecionam martelos para quebrar sementes.
P1: Martelos encontrados sobre as bigornas (0 cm) terão diferentes massas daqueles encontrados a raios de 1-20 cm e 21-100 cm.
P2: Pedras mais leves são utilizadas para quebrar sementes menores e pedras mais pesadas são utilizadas para quebrar sementes maiores.
P3: Pedras de diferentes massas são utilizadas para quebrar diferentes estágios de maturação de uma mesma espécie de semente.
A hipótese nula é que macacos prego não selecionam martelos. Assim, os pesos das pedras deverão ser similares em diferentes distâncias das bigornas, espécies de sementes e estágios de maturação.
H2: Macacos prego selecionam bigornas.
P5: Sítios de quebra serão mais próximos entre si em ambientes com menor disponibilidade de árvores de grande porte, que possam servir como rotas de fuga (pressupõe-se que neste caso, que os indivíduos (pressupõe-se agruparão para o aumento da proteção contra predadores).
MÉTODO
Área de estudo
A Caatinga é o único Bioma exclusivamente brasileiro, se estende por 750.000 Km2 e engloba todos os nove estados nordestinos e Minas Gerais (Ab’Sáber, 2003). Constitui um dos Biomas menos estudados do Brasil (Prado, 2003) e cerca de 20% da sua área está desertificada devido a processos antrópicos (Alves et al., 2009). Ações governamentais tentam mitigar este processo através de estratégias para a conservação deste domínio (MMA, 2004).
Este Bioma é caracterizado por clima seco seguindo os critérios da nova delimitação do semi-árido (MIN, 2005): i. precipitação pluviométrica média anual inferior a 800 milímetros; ii. índice de aridez de até 0,5 calculado pelo balanço hídrico que relaciona as precipitações e a evapotranspiração potencial, no período entre 1961 e 1990; e iii. risco de seca maior que 60%, tomando-se por base o período entre 1970 e 1990. Além de apresentar precipitações baixas e irregularmente distribuídas (principalmente entre os meses de fevereiro a junho), altas temperaturas e incidências de irradiação solar, grande incidência de espécies endêmicas e especializações em plantas para a redução da perda hídrica, como a presença de espinhos, folhas caducifólias e raízes profundas em árvores lenhosas (Alves et al., 2009).
ano de estudo (2009), foi registrado o acumulado de 936,6 mm3, superando a média histórica (dados cedidos gentilmente pela Empresa de Pesquisa Agropecuária do Estado do Rio Grande do Norte no ano de 2009). Os dados foram coletados numa área de Caatinga arbustiva aberta dentro e na periferia da RPPN Stossel Brito que possui uma área de 7,5 km2. Neste tipo de Caatinga, os frutos carnosos são disponíveis principalmente por herbáceas e cactos e, em média, o tamanho destes frutos são de 0,5 cm e máximo de 4 cm (Passiflora foetida, Phoradendron affine, Cissus verticillata, Capparis cynophallophora, Commiphora
leptophloeos, Pilosocereus gounellei, Cereus jamacaru). Esta unidade de conservação pode ser considerada como a área de maior representatividade da Caatinga arbustiva aberta do estado do Rio Grande do Norte devido ao seu ótimo estado de conservação, suas riqueza e diversidade biológica. Esta área possui formação geológica com pedras de alta dureza e baixa friabilidade (ortognaisse diorítico a granítico com restos de supracrustais – CPRM, 2005a).
migmatitos variandos, xístos e anfibolitos, às vezes cortados por veios de quartzo e pegamtito (CPRM, 2005b).
Em ambas as áreas de estudo, a variação sazonal influencia fortemente a paisagem e, consequentemente, a disponibilidade de recursos para a reprodução de animais e plantas. Na estação chuvosa, é observada uma vegetação exuberante com fase reprodutiva bem definida na maioria das espécies. Na estação seca, a paisagem muda drasticamente para disponibilidade de água reduzida, árvores com a ausência de folhas e frutos carnosos e é observada uma estação reprodutiva restrita a poucas espécies (a exemplo de ninhos ativos de Juriti, Leptotilla verreauxi, estes que reproduzem ao longo do ano).
Animais de estudo
Na regiao nordeste do Brasil ocorrem três espécies de macacos prego: Cebus libidinosus, C. flavius e C. xanthosternus. Cebus flavius apresenta uma distribuição espacial restrita ao nordeste brasileiro acima do Rio São Francisco, encontrada inicialmente na Mata Atlântica (Oliveira & Langguth, 2006) e recentemente Ferreira et al., (2009) encontraram uma população habitante da Caatinga do Estado do Rio Grande do Norte. Cebus libidinosus possuem uma área de ocorrência amplamente distribuída entre os Biomas Caatinga e Cerrado.
Segundo a IUCN, dentre as duas espécies de macacos prego encontradas no nordeste brasileiro acima do Rio São Francisco, Cebus flavius é a única que apresenta o status de criticamente em perigo (Oliveira et al., 2008), enquanto que Cebus libidinosus apresenta um status de menor preocupação (Rylands et al., 2008).
animais também apresetam tufos curtos dirigidos para trás, de modo que a cabeça apresenta um perfil arredondado frontal sem tufos eretos ou crista ou almofadas (Oliveira & Langguth, 2006).Os C. libidinosus possui coloração no corpo amarronzada, seus ombros possuem tons mais escuros tendendo para o vermelho, cabeça branco-amarela, possui topete pequeno comumente formando duas pequenas pontas.
Duas populações de macacos prego não habituadas a presença humana e mapeadas por Ferreira et al., (2009) foram estudadas: Cebus flavius, habitantes da Serra do Estreiro e C. libidinosus, habitantes da Serra de Luís Gomes.
Durante o estudo, foram efetivamente contabilizados grupos com o número máximo de 40 indivíduos para a população de C. flavius (n = 8 avistagens), e 50 animais para a população de C. libidinosus (n = 7 avistagens).
Esforço amostral
Entre agosto a setembro de 2008 e janeiro de 2009 (fase preliminar) e fevereiro a dezembro de 2009 (fase efetiva) foram efetuadas 26 excursões de campo com duração de cinco dias em cada área de estudo que resultaram num montante de 1.560 horas de coleta de dados.
Registro metodológico
Parcelas monitoradas
Foram definidas quatro parcelas representantes de cada área de estudo para amostragem mensal. Na escolha destas parcelas optou-se por áreas que fossem distantes entre si (superiores a 400 metros), que apresentassem disponibilidade de espécies vegetais produtoras de sementes (a saber: Manihot dichotoma, Syagrus cearensis e/ou Diclea grandiflora), e em regiões de difícil acesso onde não havia presença humana. Cada parcela mediu em média 3 hectares (em Jucurutu) e 5 hectares (em Luís Gomes). Foram realizados registros de GPS de todas as plantas produtoras de sementes. Com o objetivo de obter registros sistemáticos na coleta de dados, dentro de cada parcela foram construídas trilhas que eram percorridas na mesma direção e sentido a cada visita amostral mensal.
Durante um estudo preliminar foi verificado que havia uma grande disponibilidade de martelos e bigornas em ambas as áreas de estudo, devido a formação geológica nestes ambientes. Pedras utilizadas como martelos apresentaram grande variação de tamanho (média = 10,13 cm; I.C 95% = ± 7,02 cm – Figura 3). Bigornas poderiam ser constituídas por pedras ou troncos de árvores, e, em especial as bigornas de pedras variavam entre 10 cm a mais de 5 metros de comprimento (Figura 3).
Fenologia das espécies vegetais produtoras de sementes e frutos cultivares
Definição e registro de presença de sítio de quebra
Neste trabalho, foram considerados sítios de quebra ativos aqueles em que havia, concomitantemente, a presença de pedras menores (martelos) sobre pedras maiores (bigornas) e sementes quebradas num raio de até 20 cm da bigorna. Sítios de quebra inativos eram aqueles em que ocorria a ausência de uma destas condições. No presente trabalho, somente os sítios de quebra ativos foram mensurados.
Nos deslocamentos entre as trilhas dentro de cada parcela, era registrada a localização geográfica dos sítios de quebra ativos, além de informações sobre as sementes quebradas como a espécie e o estado de maturação. Quando eram encontrados sítios de quebra contendo sementes de diferentes espécies e/ou estado de maturação, estes eram classificados como sítios de quebra mistos. Para o registro dos dados de GPS, utilizou-se o equipamento GPSMAP® 76CSx, Garmim com unidade em UTM e datum SAD 69.
Os estágios de maturação das sementes quebradas foram classificados de acordo com as características morfofisiológicas de cada espécie de sementes. As sementes de maniçoba (Manihot dichotoma) foram classificadas como verdes, quase maduras e maduras. Sementes verdes apresentavam o epicarpo verde e volumoso e o endocarpo estava totalmente constituído; sementes quase maduras tinham epicarpos desidratados com rachaduras e prontos para a dispersão explosiva, também era evidente o mesocarpo com coloração de maduro e endocarpo seco; sementes maduras eram encontradas dispersas em torno das plantas produtoras com epicarpo maduro e seco (ver Figura 6).
de polpa; sementes maduras eram encontradas com epicarpo e endocarpo secos; o estágio podre era observado quando o endocarpo estava fermentado e constituído por larvas de insetos (ver Figura 6).
Ao final de cada registro, os sítios eram limpos, de modo que as sementes encontradas até um raio de 20 cm da bigorna eram descartadas para além dessa distância. Quando eram encontradas sementes quebradas nos sítios de quebra inativos, estas também eram descartadas seguindo o mesmo procedimento. A limpeza de sítios de quebra deu início a partir da última excursão preliminar (janeiro de 2009), e possibilitou que as visitas contínuas dos animais às parcelas resultassem em registros que indicavam o uso mensal de sítios de quebra.
Registro do peso dos martelos
Dado o grande número de sítios de quebra ativos, os registros das massas dos martelos ocorriam sequencialmente a cada 10 sítios de quebra ativos (isto é, apenas o primeiro, 10º, 20º, 30º e assim por diante). Os martelos sobre as bigornas eram coletados e medidos com o auxílio de uma balança com precisão de 1g (modelo ER2856st). Após pesados, os martelos eram retirados e depositados a mais de 20 cm da bigorna, para evitar que as mesmas pedras fossem pesadas em meses subsequentes. Ao encontrar sítios de quebra mistos, os martelos eram medidos e registrados como pertencentes a sítios de quebra mistos.
Preparação do banco de dados geográficos: Distância mais próxima entre pontos
No programa ArcMap™ ERSI software® (versão 9.3), a partir dos dados de GPS referentes às plantas produtoras de sementes e aos sítios de quebra, foram calculadas: i.) distância mais próxima entre plantas produtoras de sementes; ii.) distância mais próxima entre sítios de quebra e plantas produtoras de sementes e iii.) distância mais próxima entre sítios de quebra.
Optou-se por calcular os pontos mais próximos e não a distância euclidiana para retirar o possível erro decorrente do formato das parcelas. Assim, mesmo as parcelas possuindo áreas semelhantes em média, parcelas com formatos mais retangulares resultariam em maiores distâncias do que parcelas com formatos quadrados.
Análise de dados
As massas dos martelos amostrados sobre as bigornas foram transformados em log10 para assumir distribuição normal segundo o teste Kolmogorov-Smirnov (N > 4; D > 0,35; p >0,20) e, portanto, empregaram-se testes paramétricos para estes dados. As medidas das distâncias não apresentaram distribuição normal segundo o Kolmogorov-Smirnov (N > 146, D < 0,23; p <0,01), tendo sido empregados testes não paramétricos.
RESULTADOS
Visão geral
Os macacos prego utilizaram cerca de quatro vezes mais sítios de quebra na Caatinga arbustiva aberta que na Caatinga arbustivo-arbórea: 2.212 sítios de quebra em Jucurutu (Caatinga arbustiva aberta) e 558 sítios de quebra em Luís Gomes (Caatinga arbustivo-arbórea).
Em Jucurutu, 100% dos sítios de quebra encontrados eram exclusivamente de sementes de maniçobas, enquanto que em Luís Gomes houve uma maior variação de sementes, sendo: 75% de coco catolé (Syagrus cearensis), 16% de maniçoba (Manihot dichotoma), 1% de olho de boi (Diclea grandiflora) e 8% para sítios de quebra mistos. Devido ao baixo registro da quebra de sementes de D. grandiflora, optou-se por não utilizar estes dados nos testes de hipóteses.
A quebra de sementes em diferentes estágios de maturação foi observada em ambas as áreas de estudo. O grupo de C. flavius (Jucurutu) foi o único a quebrar sementes de maniçoba verdes (6%), quase maduras (2%) e maduras (92%). Por outro lado, o grupo de C. libidinosus (Luís Gomes), habitante de uma região onde também eram encontradas sementes de maniçoba, quebrou apenas sementes de maniçoba maduras, mas abriu cocos catolé em diferentes estágios de maturação, sendo 8% do total para sementes verdes, 73% sementes maduras, 15% sementes podres e 4% sementes mistas.
Escolha dos martelos para a quebra de sementes
encontrados entre os raios de 1 a 20 cm e 21 a 100 cm de distância da bigorna (ver estatística na Tabela 1 e Figuras 4A e B). Entretanto, o padrão diferiu entre as áreas.
Em Jucurutu, o padrão encontrado foi de pedras localizadas acima da bigorna com massa significativamente menor (mediana = 46 g) do que aquelas localizadas nos arredores da mesma (medianas para raios entre 1-20 cm = 131g e 21-100 = 230 g – Figura 4A). O oposto foi verificado em Luis Gomes: pedras acima da bigorna significativamente com maior massa (mediana = 429 g) do que as pedras encontradas aos arredores (medianas para 1-20 cm = 32g e 21-100 = 52,5 g – Figura 4B).
Figura 4– Análises estatísticas correspondentes à predição 1. (A) Caatinga arbustiva aberta: massa
das pedras encontradas sobre a bigorna (0 cm, N = 414); entre 1-20 cm da bigorna (N =162) e entre
21-100 cm da bigorna (N=363). (B) Caatinga arbóreo-arbustiva: massa das pedras encontradas sobre a
bigorna (0 cm, N = 93); entre 1-20 cm da bigorna (N =115) e entre 21-100 cm da bigorna (N=346).
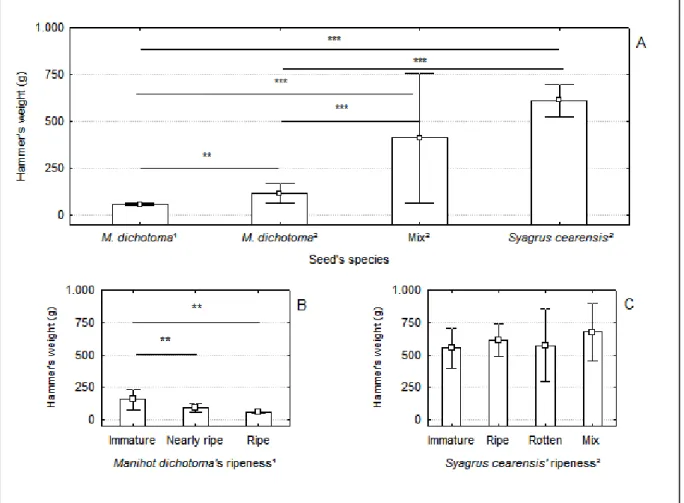
Em relação ao peso dos martelos, foi verificado que estes variaram de acordo com a espécie da semente (predições 2) a ser quebrada. Para sementes menores (Manihot dichotoma) as pedras utilizadas como martelos eram mais leves (médias entre Jucurutu e Luís Gomes = 89,01g) do que para sementes maiores (Syagrus cearensis, média = 612,65g). O peso dos martelos não diferiu entre os sítios de quebra mistos e os sítios de quebra de S. cearensis na Caatinga arbustivo-arbórea (médias: sítios de quebra mistos = 412,25 e sítios de quebra de S. cearensis = 612,65g). Ver Tabela 1 e Figura 5A e 6.
Ainda foi observado que, a depender da área de estudo, ocorreram diferenças significativa quanto ao peso dos martelos utilizados para quebrar uma mesma espécie de semente (M. dichotoma). Na Caatinga arbustiva aberta, onde existem exclusivamente sementes de M. dichotoma,os martelos encontrados sobre as bigornas foram duas vezes mais leves do que os martelos encontrados na Caatinga arbóreo-arbustiva para quebrar esta mesma espécie de semente (médias: Jucurutu = 59,25g e Luís Gomes = 118,77g). Ver Tabela 1 e Figura 5A e 6.
Figura 5 – Análises estatísticas correspondentes às predições 2 e 3. (A) Massa dos martelos utilizados
para quebrar sementes por macacos prego em ambos os tipos de Caatinga. M. dichotoma1 (N= 414);
M. dichotoma2 (N= 13); Mix2 (Sítios de quebra mistos, N = 4) e Syagrus cearensis2 (N =88). (B)
Martelos utilizados pelo grupo de Cebus flavius na quebra da semente de M. dichotoma em diferentes
estágios de maturação. Sementes verdes (N= 9); sementes quase maduras (N = 9) e sementes maduras
(N=393). (C) Martelos utilizados pelo grupo de C. libidinosus na quebra da semente de Syagrus
cearensisem diferentes estágios de maturação. Sementes verdes (N= 12); sementes maduras (N = 50);
sementes podres (N = 11) e sítios de quebra mistos (N = 15). média e intervalos de confiança da
média a ±95%; ** p < 0,01; *** p < 0,001; Manihot dichotoma1 = Caatinga arbustiva aberta;
Tabela 1 – Compilação dos resultados estatísticos.
Hipotesis a Type of test a2 N a3 Z a4 P
P1 - Open scrub Caatinga Kruskal-Wallis test 939 201,33 <0,001
P1- Scrub-arboreal Caatinga Kruskal-Wallis test 554 141,96 <0,001
P2 One-way ANOVA 516 267,517 <0,001
P3 - Open scrub Caatinga Repeated measured ANOVA 411 6,88 <0,01
P3 - Scrub-arboreal Caatinga Repeated measured ANOVA 88 0,534 0,66
P4 Kruskal-Wallis test 1685 117,48 <0,001
P5 Kruskal-Wallis test 1912 -15,98 <0,001
Tabela 2 – Resultados das correlações correspondentes a predição 4. Códigos: Espécie1 = Caatinga arbustiva aberta; Espécie 2 = Caatinga arbustivo-arbórea.
Type of test Matrix Correlations N Spearman's r Z P
Spearman's correlation of M. dichotoma1 Distances between seed-producing species
Distances between nut-crack sites and seed-producing species 1306 0,02 0,87 0,38
Distances between nut-crack 1306 -0,14 -5,2 < 0,001
Spearman's correlation of M. dichotoma2 Distances between seed-producing species
Distances between nut-crack sites and seed-producing species 65 -0,25 -2,1 <0,05
Distances between nut-crack 88 -0,08 -0,7 0,46
Spearman's correlation of S. cearensis2
Distances between seed-producing species
Distances between nut-crack sites and seed-producing species 233 0,03 0,5 0,62
Escolha espacial do sítio de quebra
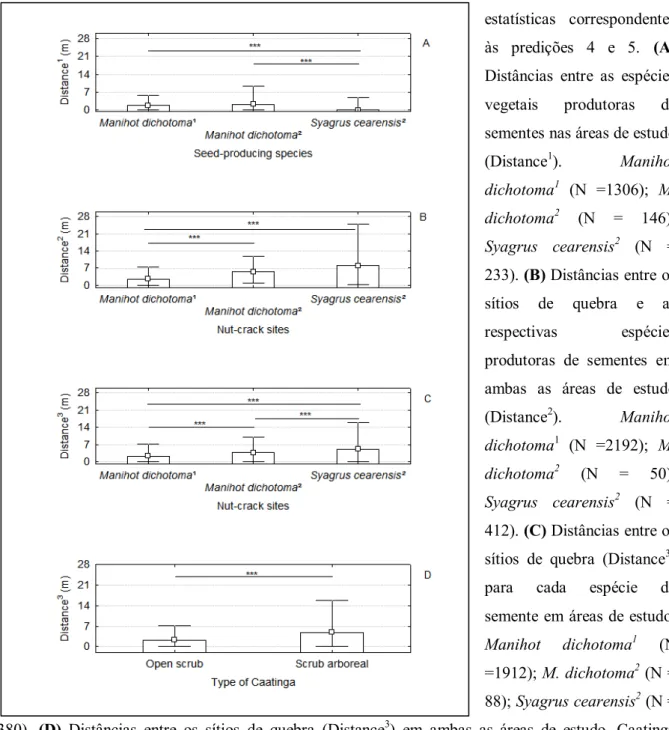
A distância entre as plantas produtoras de cocos catolé (Syagrus cearensis) foram significativamente menores (mediana = 0 m; média = 2,72 m) do que as distâncias entre plantas de Manihot dichotoma (medianas/médias Jucurutu e Luís Gomes = 1,86/2,29 e 2,37/3,42 m; respectivamente) para ambas as áreas de estudo. As distâncias entre as plantas da espécie M. dichotoma foram semelhantes em ambas as áreas de estudo.Ver Tabela 1 e Figura 7A.
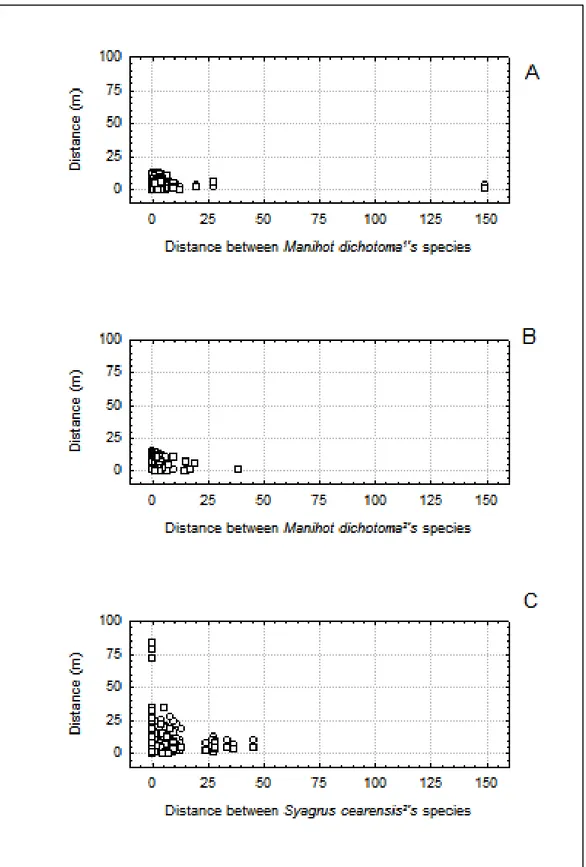
Em suporte à predição 4, foi observado que os sítios de quebra se distribuem de forma independente da distância entre as plantas produtoras de sementes. Não ocorreram correlações significativas positivas com alto poder explicativo entre as distâncias de plantas produtoras de sementes e as distâncias entre sítios de quebra às plantas. Este mesmo padrão ocorreu ao correlacionar as distâncias de plantas produtoras de sementes às distâncias entre os sítios de quebra. Ver Tabela 2 e Figura 8.
Apesar de plantas de maniçoba (Manihot dichotoma) apresentarem maiores distâncias inter-individuais do que a distância registrada entre os espécimes de cocos catolé (Syagrus cearensis), os sítios de quebra de sementes maniçoba estavam mais próximos das plantas produtoras do que os sítios de quebra de cocos catolé. Ver Tabela 1 e Figura 7B.
Também foi verificado que as distâncias entre os sítios de quebra diferiram de acordo com o tipo de semente: sítios de quebra de maniçobas estavam mais próximos entre si (medianas maniçobas de Jucurutu = 2,24 e Luís Gomes =3,72) do que os sítios de quebra de cocos catolés (mediana = 5,04). Ver Tabela 1 e Figura 7C.
estavam significativamente mais próximos (mediana = 2,24) do que na Caatinga arbusitvo-arbórea (mediana = 4,91). Conferir Tabela 1 e Figura 7D.
Figura 7 – Análises
estatísticas correspondentes
às predições 4 e 5. (A)
Distâncias entre as espécies
vegetais produtoras de
sementes nas áreas de estudo
(Distance1). Manihot
dichotoma1 (N =1306); M.
dichotoma2 (N = 146);
Syagrus cearensis2 (N =
233). (B) Distâncias entre os
sítios de quebra e as
respectivas espécies
produtoras de sementes em
ambas as áreas de estudo
(Distance2). Manihot
dichotoma1 (N =2192); M.
dichotoma2 (N = 50);
Syagrus cearensis2 (N =
412). (C) Distâncias entre os
sítios de quebra (Distance3)
para cada espécie de
semente em áreas de estudo.
Manihot dichotoma1 (N
=1912); M. dichotoma2 (N =
88); Syagrus cearensis2 (N =
380). (D) Distâncias entre os sítios de quebra (Distance3) em ambas as áreas de estudo. Caatinga
arbustiva aberta (N =2192), Caatinga arbustivo-arbórea (N = 462). medianas e distribuição não
outlier dos dados; *** = p < 0,001; Manihot dichotoma1 = Caatinga arbustiva aberta; Manihot
DISCUSSÃO
Estudos com diversas espécies de macacos prego habitantes de ambientes secos afirmam que a quebra de sementes representa uma estratégia de forrageio, tanto em Biomas de Caatinga (Cebus libidinosus: Moura & Lee 2004; Ferreira et al., 2009, 2010; C. xantosternos: Canale et al., 2009; C. flavius: Emidio & Ferreira 2009), de Cerrado (C. libidinosus: Waga et al., 2006; C. xantosternos: Canale et al., 2009) e no ecótono Caatinga-Cerrado (C. libidinosus: Fragaszy et al., 2004a; Visalberghi et al., 2007; C. xantosternos: Canale et al., 2009). Desta forma, nossos resultados corroboram os achados anteriores.
Paraíba: Langguth & Alonso, 1997; Serra do Estreito, Rio Grande do Norte: Emidio & Ferreira, 2009; Serra de Martins e Serra de Luís Gomes, RN: Ferreira et al., 2010; Fazenda Boa Vista, Piauí: Fragaszy et al., 2004a; Visalberghi et al., 2007; Silva, 2008; Serra da Capivara, PI: Mannu & Ottoni, 2008), o que é indicativo de convergência comportamental devido a semelhança ecológica.
De forma geral, nossos dados oferecem suporte à hipótese de que, apesar da abundância de material disponível, o uso de pedras por macacos prego não ocorre de forma aleatória, mas envolve escolhas, tanto do martelo quanto da bigorna. Estas escolhas diferem de acordo com as condições ecológicas encontradas nas áreas de estudo e são ajustadas de acordo com as espécies de sementes a serem quebradas.
Escolha do martelo – eficiência na quebra de sementes
disponibilidade de potenciais martelos próximos às bigornas, portanto, sugerimos que macacos prego apenas escolhem as pedras mais adequadas sem a necessidade de transportá-las.
A seleção de martelos eficientes e disponíveis foi relatada por Schrauf et al., (2008) em condições de laboratório e por Vilsaberghi et al., (2009a) em um experimento com macacos prego selvagens. Estes autores descreveram diversas tarefas para quebrar sementes de palmeiras com martelos artificiais em condições nas quais martelos eficientes eram aqueles que apresentavam maior resistência ou peso. Os resultados mostraram que os C. libidinosus escolheram, transportaram e quebraram sementes de palmeiras com os martelos mais efetivos. Ferreira et al., (2010) comparando três áreas de ocorrência de macacos prego com alta disponibilidade de material de quebra, verificaram que estes quebraram sementes de Atallea oleifera com martelos significativamente duas vezes mais pesados do que aqueles utilizados na quebra de sementes de Syagrus cearensis (as sementes mais resistentes do presente estudo), e 18 vezes mais pesados do que os martelos utilizados para quebrar sementes de Manihot dichotoma (as sementes menos resistentes do nosso trabalho).
Nossos dados revelam informações que vão além de escolhas de pedras raras para quebrar sementes resistentes. As pedras utilizadas pelas duas populações de macacos prego monitoradas (Cebus flavius e C. libidinosus) indicam que estes animais estão selecionando aquelas mais eficientes de acordo com a espécie de semente a ser quebrada, apesar da alta disponibilidade de pedras alternativas potencialmente adequadas para a quebra das sementes.
foi observada pelo grupo de C. libidinosus habitante da Caatinga arbóreo-arbustiva. Estes consumiram cocos catolé (Syagrus cearensis) em diferentes estágios de maturação, porém, utilizaram martelos com massas semelhantes para quebrar sementes em diversos estados de maturação. Da mesma forma, sementes de M. dichotoma são quebradas em ambas as áreas, entretanto, foi identificado que o grupo de C. flavius selecionou martelos duas vezes mais leves do que o grupo de C. libidinosus.
Sugerimos que esta diferença encontrada entre as escolhas de martelos não representam diferenças entre espécies, mas sim, da riqueza florística das áreas em que os animais vivem. Na Caatinga arbustiva aberta, 100% dos sítios de quebra encontrados de sementes de maniçobas (M. dichotoma), enquanto que na Catinga arbustivo-arbórea houve uma maior variação no consumo de espécies de sementes quebradas. Deste modo, na Caatinga arbustiva aberta parece ocorrer uma maior especialização (cultural ou através de tentativa e erro, tipos de aprendizados preditos por van Schaink et al., 1999) em quebrar sementes de maniçoba com pedras menores. Por outro lado, os macacos prego da Caatinga arbóreo-arbustiva parecem adotar estratégias mais generalistas nas atividades de quebrar sementes.
Escolha das bigornas
–
estratégias para reduzir riscos de predação e custos de
competição direta.
frequentemente a mais de 30m (Boesch & Boesch, 1983). Nossos achados dão suporte a abordagem construída por Vilsaberghi et al., (2009b) através de comparativos entre as relações dos deslocamentos no chão e riscos de predação em macacos prego e chimpanzés. Estes autores observaram que os macacos prego transportam martelos para os sítios de quebra a distâncias significativamente menores do que os chimpanzés. Tais autores sugerem que os custos no transporte de martelos são maiores em macacos prego do que nos chimpanzés, devido a maior vulnerabilidade de predação. Portanto, estes animais possuem maior aversão a grandes deslocamentos terrestres em condições que comprometem o equilíbrio e a velocidade.
observadas em áreas circunvizinhas (lista completa de potenciais predadores em ambas as áreas de estudo em Ferreira et al., 2009).
Destaca-se que, das duas espécies estudadas, apenas em C. flavius foram registradas ocorrências de uso de instrumentos na tentativa de afugentar potencias predadores, o que pode representar estratégias adicionais para reduzir riscos de predação. Nossos resultados corroboram com os achados por Mannu & Ottoni (2008) e Chevalier-Skolnikoff (1990) que observaram a emissão de comportamentos teatrais seguidos de lançamento de folhas, pedaços de troncos ou rochas a potenciais predadores em macacos prego (C. libidinosus e C. capucinus). No presente estudo, quando os C. flavius constatavam a presença do pesquisador, estes animais deslocavam-se para rochões mais altos, formavam grupos coesos, emitiam vocalizações e lançavam pedras em direção ao solo (Emidio & Ferreira, 2009). Nossos achados indicam que a falta de oportunidades para fugas individuais tornam os grupos mais coesos durante a permanência no chão para quebra de sementes.
As análises da distância entre os sítios de quebra possibilitam ainda inferir estratégias comportamentais adotadas para reduzir a competição por alimento. Foi possível identificar numa mesma área (Caatinga arbóreo-arbustiva) que os sítios de quebra de sementes menores e menos calóricas (i.e. M. dichotoma: volume da semente = 0,24cm3 – Ferreira et al., 2010; valor calórico de Manihot spp. = 1,13 cal/g – dados ajustados de Martins et al., 2007) localizaram-se a menores distâncias entre si do que os sítios de quebra de sementes maiores e mais calorias (i.e. S. cearensis – volume da semente = 16cm3– Ferreira et al., 2010; valor calórico de S. coronata = 5,27 cal/g – dados ajustados de Crepaldi et al., 2001).
de usurpação de amêndoas prontas para o consumo por co-específicos que sejam capazes de quebrar sementes com eficiência. Ottoni & Mannu (2001) foram os primeiros a relatarem relações de roubo nas atividades de quebra de sementes e revelam que apenas os infantes e juvenis são tolerados, permanecendo mais próximos aos animais mais proficientes em quebrar sementes. Ottoni et al., (2005) discutem que há uma relação de benefício e custo na tolerância na usurpação de alimento. Assim, ao ficarem observando a quebra de sementes, os animais tolerados roubam as amêndoas e também aprendem as técnicas para abrir itens encapsulados. Resultados semelhantes foram observados por Silva (2008) que estudou uma população de C. libidinosus habitante do ecótono Caatinga-Cerrado, sugerindo que é característico do gênero.
É típico de animais gregários apresentarem organizações espaciais que balanceiem as relações tênues entre o sucesso de forrageio e custos de predação (Hamilton, 1971). Macacos prego (Cebus spp.) são conhecidos por adotarem com destreza estratégias espaciais para reduzir a competição por alimento e riscos de predação. Em diversas espécies de macacos prego, Robinson (1981 –C. nigritus), entre outros autores (Janson, 1988 – C. apella; Phillips, 1995b, Hall & Fedigan, 1997 – C. capucinus), também confirmam que a depender das pressões de predação, os grupos podem tornar-se mais frouxos ou densos. Em nossas áreas de estudo não há escassez de bigornas adequadas para quebra de sementes. Assim, sugerimos que as escolhas dos locais das bigornas, refletem um compromisso entre custo de evitar potenciais usurpadores e a necessidade de proximidade interindividual para proteção contra predadores em áreas com baixa disponibilidade de rotas de fuga.
CONCLUSÕES