Características fenotípicas de Enterobacter cloacae controladas pelo sistema quorum sensing
Texto
Imagem






Documentos relacionados
O falo, assim, é o significante (do desejo) da completude imaginária, inacessível, pois da ordem do Real (LACAN, 1985b, p. O falo é, pois, o significante ordenador da estrutura e
A descrição da vontade de verdade e da vontade de saber realizada por Foucault (2000a), nos permitiu observar a constituição da medida como norma e, a realidade do discurso como
Além de diminuir os riscos das instâncias subnacionais, houve um incremento nos estímulos para que estados e municípios estruturem suas políticas e assumam maiores
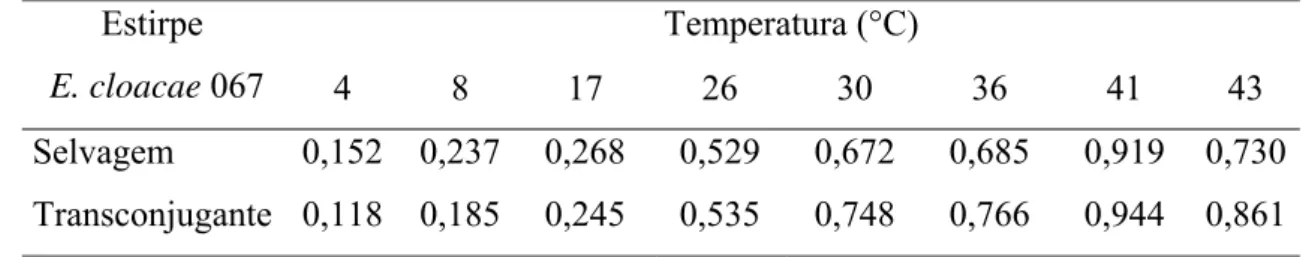
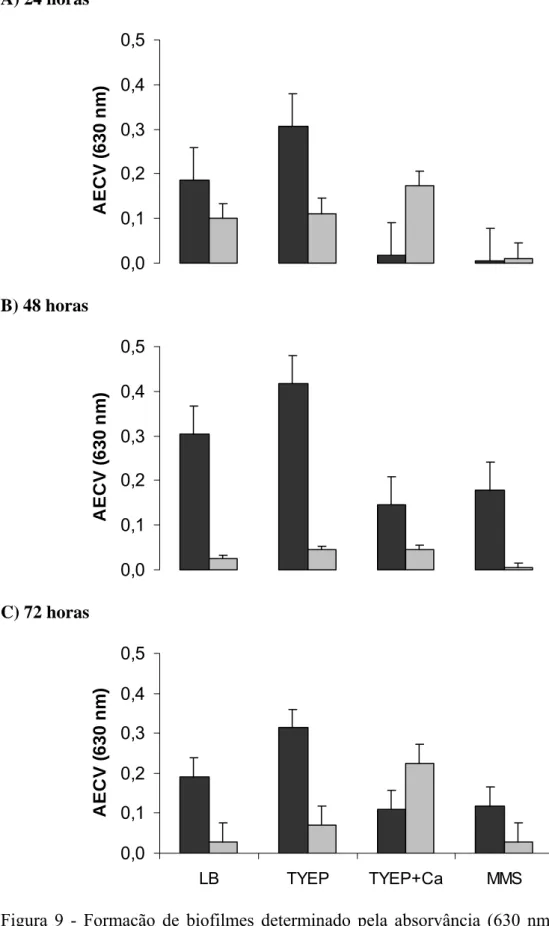
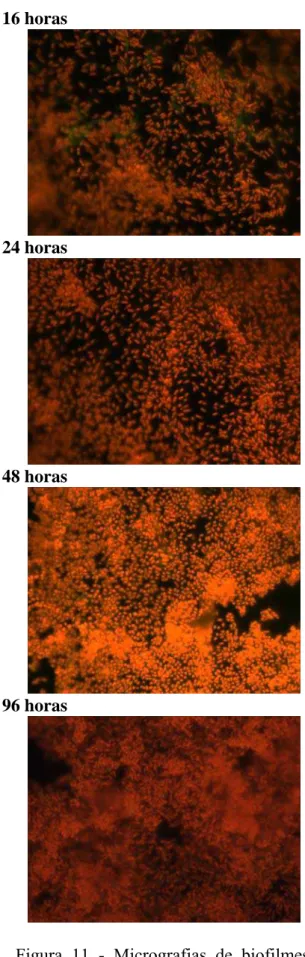
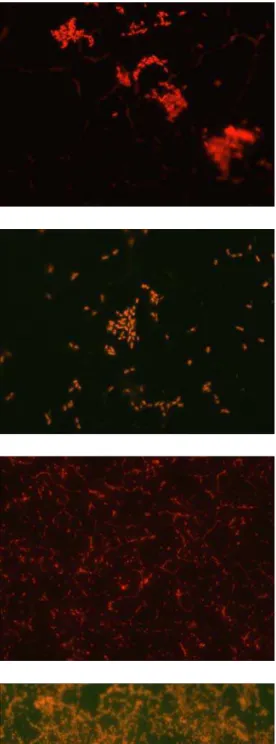
Constam na tabela 2, os valores médios da variável de remoção de CE para o fator tipo de material orgânico dentro de cada nível de tempo de aplicação e cada nível de
Inventário e avaliação da sanidade das árvores de Vila Real...
Diante do exposto, este trabalho tem como objetivo analisar, através de técnicas de simulação e de otimização, a dinâmica operacional do processo de atendimento
O Comitê Gestor é um organismo colegiado formado por representantes da administração pública paulista e coordenado pelo Arquivo Público do Estado de São Paulo, unidade
massa de ovos, percentuais de eclosão e eficácia de nove carrapaticidas em partenóginas de Boophilus microplus (Teste in vitro), colhidas de bovinos pertencentes a 17
