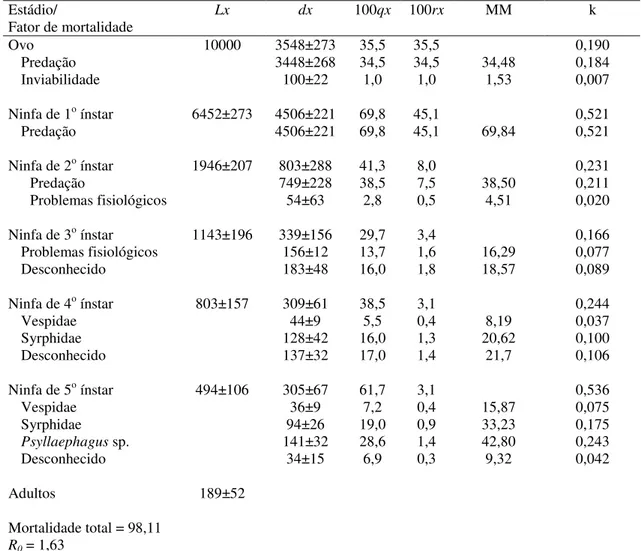
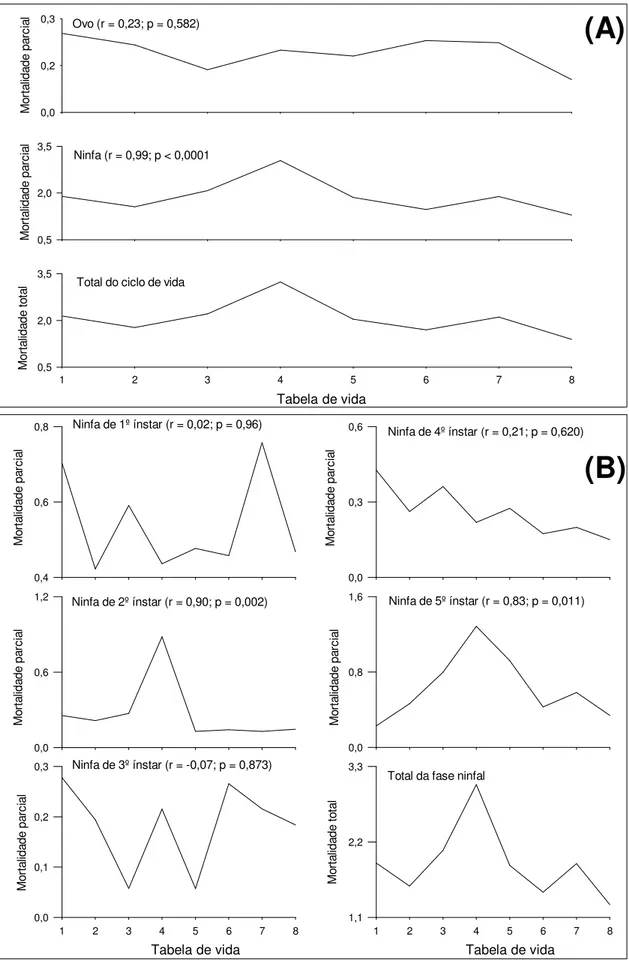
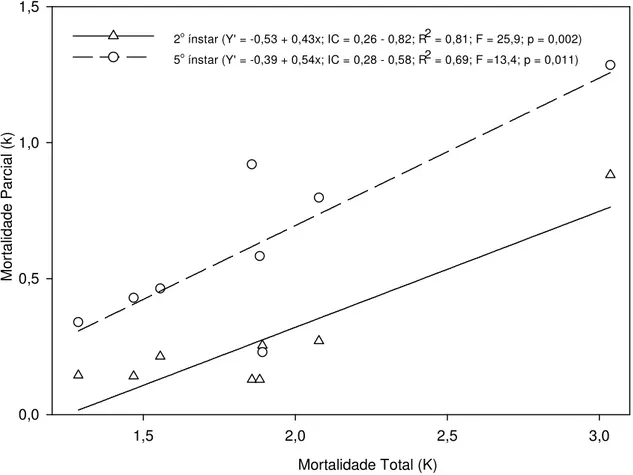
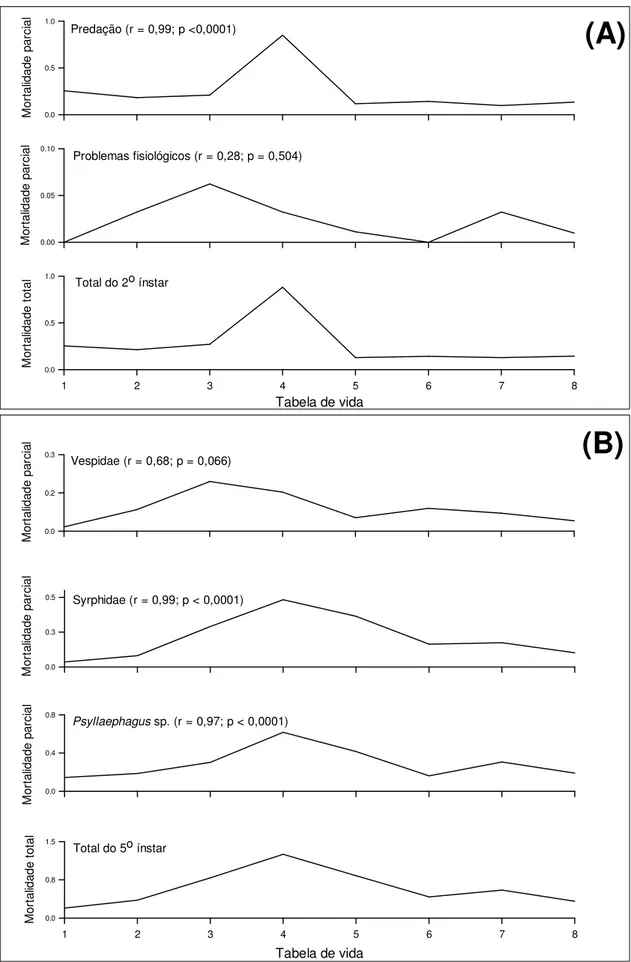
Controle natural de Triozoida limbata em goiabeira
Texto
Imagem
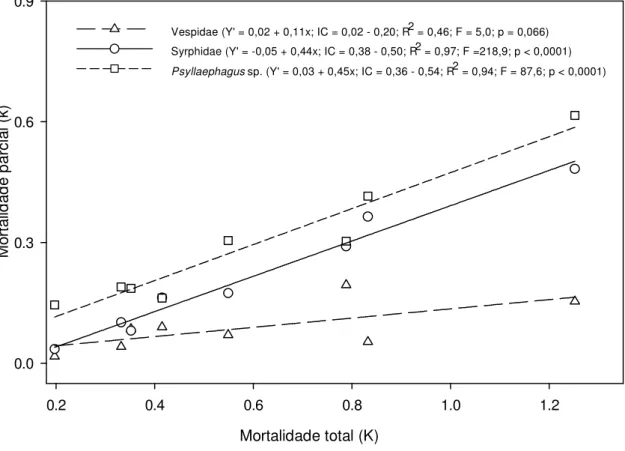
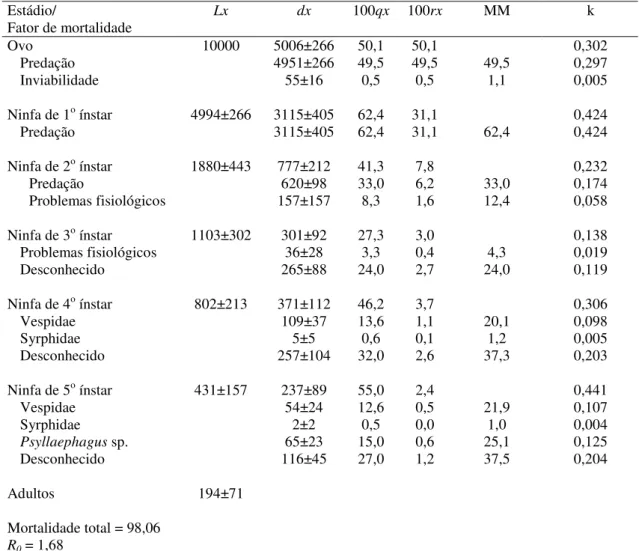
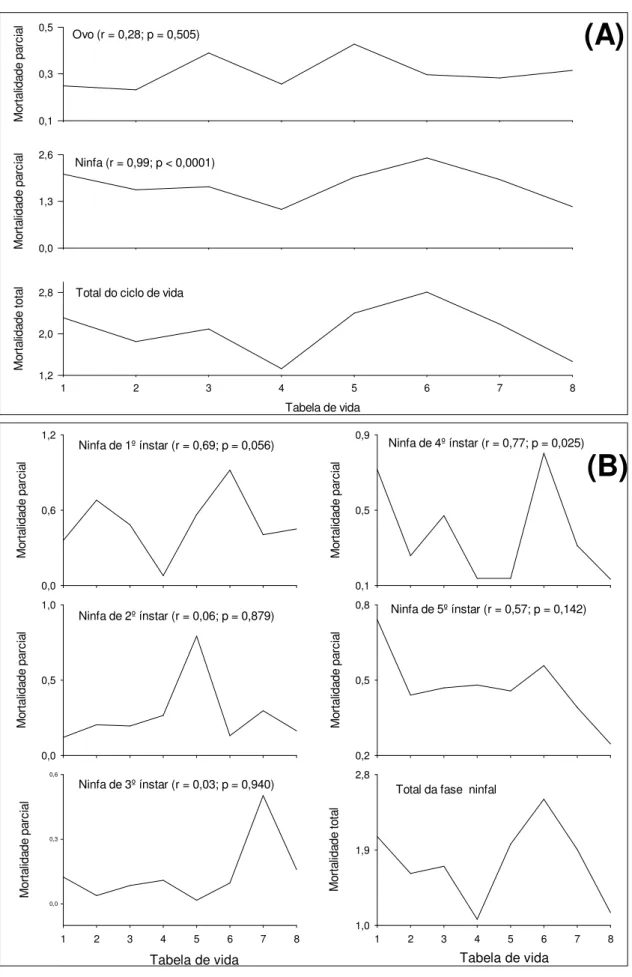
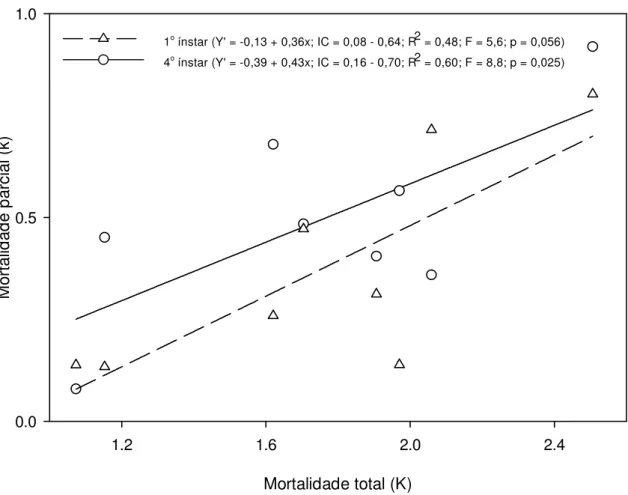
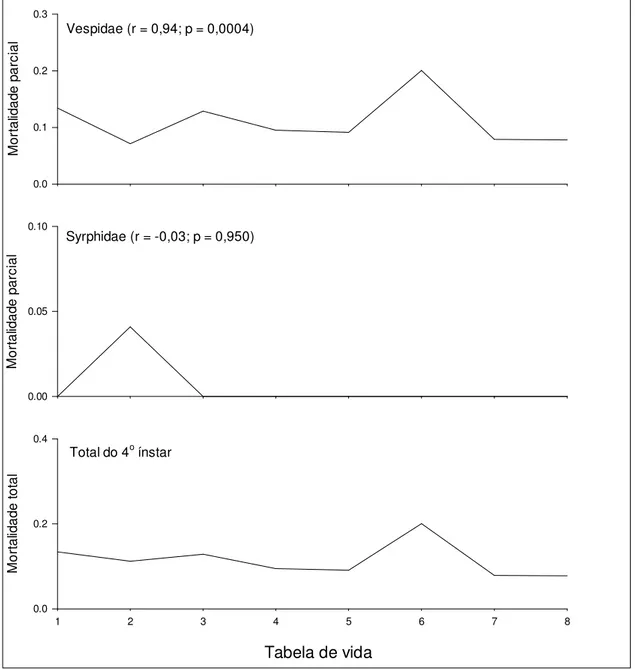
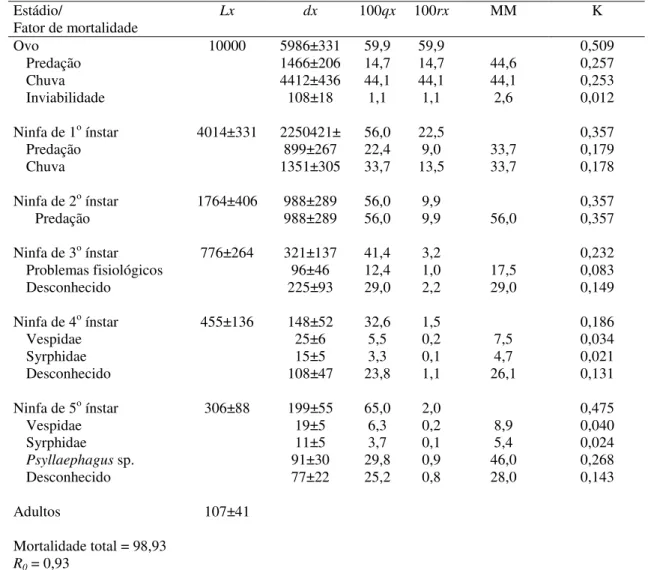
Outline
Documentos relacionados
Faça um programa para que o robô, no ambiente dado pelo mapa passBeacons.map, saia da posição inicial (salão à esquerda) e vá para o ponto marcado em branco no salão à
As IMagens e o texto da Comunicação (com as legendas incluídas) devem ser enviadas por correio eletrônico. Comitê
A USP se destaca por possuir produção significativamente maior que as dos demais programas (quase 70 artigos). O segundo grupo, composto por Unicamp, UnB, UFRGS
A proposta da tese surge ainda das necessidades de reflexão sobre o trabalho com música no dia a dia da escola de educação infantil e, com isso, de questões
Receita de Produtos (Quantidade) Margem de Produtos (Qualidade) Alavancadores de Produtividade Investimento Investimento Processo Processo Mercado Mercado
(2013 B) avaliaram a microbiota bucal de oito pacientes submetidos à radioterapia na região de cabeça e pescoço através de pirosequenciamento e observaram alterações na
Na aplicação das políticas contábeis da Companhia e das controladas, a Administração deve fazer julgamentos e elaborar estimativas a respeito dos valores contábeis
Cesca (2004) aponta em seu estudo a importância do psicólogo buscar garantir os direitos dos envolvidos, a busca da cidadania e visar a saúde mental dos mesmos, o autor
