Melhoramento Genético V
egetal |
Artigo
Heterose e seus componentes em híbridos
de populações F
2
de milho
Sergio Doná (1*); Maria Elisa Ayres Guidetti Zagatto Paterniani (2); Paulo Boller Gallo (3);
Aildson Pereira Duarte (1)
(1) APTA, Polo Regional de Desenvolvimento do Médio Paranapanema, Caixa Postal 263, 19800-000 Assis (SP), Brasil.
(2) Instituto Agronômico (IAC), Centro de Análise e Pesquisa Tecnológica do Agronegócio de Grãos e Fibras, Av. Barão de Itapura, n.1481, 13020-902 Campinas (SP), Brasil.
(3) APTA, Polo Regional de Desenvolvimento do Nordeste Paulista, Caixa Postal 58, 13730-970 Mococa (SP), Brasil. (*) Autor correspondente: sergio@apta.sp.gov.br
Recebido: 15/abr./2011; Aceito: 2/maio/2011
Resumo
Objetivou-se verificar o potencial de 28 híbridos de populações F2 de milho, obtidos de um dialelo completo, juntamente
com oito populações F2 parentais e duas testemunhas comerciais quanto a caracteres agronômicos e à heterose e seus componentes. Os experimentos foram desenvolvidos em 2008/2009, em Campinas, Mococa e Palmital, Estado de São Paulo, no delineamento experimental de blocos ao acaso com três repetições. Avaliaram-se os caracteres florescimento masculino, altura da espiga e produtividade de grãos, que foi corrigida para 14% de umidade e estande ideal. Efetuaram-se análises de variância individuais e conjuntas, considerando-se o modelo fixo, sendo as médias comparadas pelo Teste de Tukey, além das análises dialélicas pelo modelo de Gardner e Eberhart (1966). Verificaram-se efeitos de heterose e seus componen -tes altamente significativos. Considerando-se a produtividade de grãos, as populações F2AG8060, F2Fort e F2A2555 foram promissoras em combinações híbridas, com elevados valores de heterose de parental e a F2DAS2B710 com o maior efeito de parental. Houve manifestação de heterose para produtividade de grãos com destaque para o híbrido F2A2555 x F2Fort, confirmando-se o potencial produtivo de híbridos de populações F2 como alternativa para a produção comercial de milho,
visando aos produtores de média a baixa tecnologia.
Palavras-chave:Zea mays, dialelo completo, Gardner e Eberhart, produtividade.
Heterosis and its components in F
2populations of maize hybrids
Abstract
The potential of 28 hybrids maize F2 population, obtained from a complete diallel among eight F2 populations, along with the
eight parental F2 populations and two commercial controls were evaluated for agronomic traits, heterosis and its compo
-nents. The experiments were developed in 2008/09, in Campinas, Mococa and Palmital, São Paulo State, Brazil. The experi -mental design was in randomized blocks, with three replications. The agronomic traits male flowering, ear height and grain yield were evaluated. The grain yield was adjusted to 14% of moisture. Individual and grouped variance analyses were made considering the fixed model, with the means being compared by the Tukey test. The Gardner and Eberhart (1966) model was used in the diallel analysis. It was observed that the effect of heterosis and its components were highly significant. Consider -ing the grain yield, populations F2AG8060, F2Fort, F2A2555 are promising in hybrid combinations, with high values of parental heterosis. The population F2DAS2B710 had the greatest parental heterosis effects. There was heterosis manifestation for grain yield, mainly for hybrid F2A2555 x F2Fort. It was confirmed the potential of F2 population hybrids as an alternative to the commercial maize production with medium to low technological input.
1. INTRODUÇÃO
A produtividade média de grãos da cultura do milho na Região Centro-Sul brasileira é relativamente baixa, com valor de 4.685 kg ha-1 na safra de 2008/2009 (Conab,
2009). Este valor está abaixo do obtido nas lavouras bra-sileiras desenvolvidas com elevado nível tecnológico, que
se aproxima ao da média norte-americana, 9.659 kg ha-1
(Usda, 2010), maior produtor mundial deste cereal. Do total de sementes de milho comercializadas no Brasil na safra de 2007/2008, 57,48% foram de híbridos simples (Cruz e Pereira Filho, 2010). Estes se destacam como os mais produtivos, porém o alto custo das semen-tes dificulta seu uso por produtores de nível tecnológico marginal ou baixo, que representam cerca de 67% da área cultivada no país (Mattoso e Melo Filho, 2006) e que utilizam variedades ou sementes de paiol de polinização aberta, um dos motivos da baixa produtividade no Brasil.
A obtenção de híbridos simples demanda um tempo elevado para se obter linhagens homozigóticas (Miranda Filho e Viégas, 1987; Carvalho et al., 2003) e para que este híbrido chegue ao mercado. As autofecundações sucessi-vas e a avaliação das linhagens quanto à capacidade de com-binação constituem as etapas mais trabalhosas e onerosas dos programas de híbridos de milho, encarecendo o preço final da semente híbrida. Este custo poderia ser reduzido pelo em-prego da geração F2 dos híbridos simples para a obtenção de híbridos duplos ou híbridos de F2 (Souza Sobrinho et al., 2002). Dessa forma, seriam eliminadas as etapas de obten-ção e a manutenobten-ção das linhagens parentais do híbrido, pois seriam produzidas apenas as gerações F2 de cada parental, as quais poderiam ser mantidas em lotes isolados e utilizadas todos os anos para a obtenção do híbrido de F2.
Souza Sobrinho et al. (2002) testaram a utilização da geração F2 de populações oriundas de híbridos
sim-ples, para produzir híbridos duplos de F2 e constataram que estes híbridos têm desempenho muito semelhan-te àqueles derivados da geração F1 de híbridos simples. Posteriormente, Amorim e Souza (2005) identificaram híbridos F2 intrapopulacionais com produtividade média de grãos superior a 10 t ha-1.
Hallauer e Miranda Filho (1981) apresentaram resultados de 1.394 cruzamentos intervarietais, obtendo heteroses médias em relação à média dos pais de 19,5%, para produção de grãos, evidenciando que pode existir grande potencial para a exploração da heterose em popu-lações e variedades de milho. A manifestação de heterose nos híbridos depende de genes com algum grau de domi-nância (parcial, completa ou sobredomidomi-nância) e também das diferenças entre frequências gênicas nas populações genitoras (Miranda Filho e Viégas, 1987).
O potencial heterótico de variedades e de popula-ções de milho em combinapopula-ções híbridas pode ser ava-liado pela utilização e análise de cruzamentos dialélicos. Dentre os métodos de análise dialélica tem-se a proposta
por Griffing (1956), na qual são estimados os efeitos de capacidade geral e específica de combinação e a de Gardner e Eberhart (1966) que visa avaliar o potencial de variedades per se e em combinações híbridas, por meio de estimativas de efeitos varietais e da heterose manifesta-da no híbrido (Cruz, 2006).
O uso destes métodos tem possibilitado a diversos autores a escolha de populações e híbridos mais promis-sores, com base na capacidade de combinação dos paren-tais e no potencial heterótico das combinações híbridas (Gorgulho e Miranda Filho, 2001; Silva e Miranda Filho, 2003; Oliveira et al., 2004).
Gardner e Eberhart (1966) apresentaram um mé-todo para análise de tabelas dialélicas não só de linhagens, como também para variedades e populações, permitindo um estudo mais detalhado da heterose. A análise de cruzamentos dialélicos oferece ao melhorista grande número de informa-ções genéticas que vêm sendo utilizadas, visando avaliar o potencial heterótico de variedades e de populações de milho em combinações híbridas, possibilitando a escolha de popu-lações mais promissoras, com base na capacidade de combi-nação dos parentais (Gorgulho e Miranda Filho, 2001; Silva e Miranda Filho, 2003; Oliveira et al., 2004).
Diferenças na capacidade de combinação de popula-ções F2 de milho proporcionam diferentes níveis de heterose
e de produtividade de grãos em seus híbridos. Dessa forma, o presente trabalho teve por objetivos identificar híbridos promissores de populações F2 de milho, oriundos de gera-ções F2 de híbridos simples comerciais, quanto a caracteres agronômicos e estimar parâmetros genéticos dos parentais e a heterose dos híbridos e seus componentes utilizando-se o método de Gardner e Eberhart (1966).
2. MATERIAL E MÉTODOS
Oito populações F2 de milho, denominadas de P1 a P8,
obtidas pela autofecundação de híbridos simples comer-ciais, foram cruzadas em esquema dialélico balanceado, incluindo híbridos e parentais, obtendo-se 28 híbridos de F2. Uma breve descrição dos híbridos simples utilizados
está contida na tabela 1. Para cada um dos cruzamentos,
Cultivar Empresa Populações Aparência
dos grãos
AG 8060 Monsanto P1 D
Fort Syngenta Seeds P2 SD
DAS 2B710 Dow AgroSciences P3 SD
30F87 Pioneer P4 SDT
A 2555 Bayer Seeds P5 SDT
AS 1548 Agroeste P6 D
30K75 Pioneer P7 SD
DKB 330 Dekalb P8 SDT
D = duro; SD = semiduro; SDT = semidentado.
Tabela 1. Descrição dos híbridos simples (HS) comerciais de
utilizaram-se como genitor feminino, populações F2 se-meadas em 2 linhas de 10 m de comprimento; no floresci-mento, foram coletados polens de aproximadamente cem plantas do genitor masculino, sendo misturados e utiliza-dos para realização das polinizações. Não foram efetuautiliza-dos cruzamentos recíprocos.
Os 28 híbridos de populações F2 mais as oito popu-lações F2 parentais e as testemunhas comerciais foram avaliados em 2008/2009, sob delineamento experimental de blocos ao acaso com três repetições, sendo cada parce-la constituída por duas linhas de 5 m espaçadas de 0,85 m, com um total de 50 plantas. Por ocasião do plantio, foram aplicados 350 kg ha-1 da formulação 08-28-16 +
0,5% de Zn e, aos 25 a 30 dias após semeadura foi reali-zada adubação de cobertura com aplicação de 200 kg ha-1
de uréia (45% N) a lanço, em Campinas; 250 kg ha-1 de
uréia (45% N) incorporada em Mococa e 312,5 kg ha-1 de
nitrato de amônio (32% N) a lanço, em Palmital. As testemunhas empregadas foram IAC 8333, um híbrido comercial resultado do cruzamento de dois sinté-ticos de alta divergência genética e de boas características agronômicas, e o DKB 350, híbrido triplo representativo em área e produtividade no Estado de São Paulo.
Os experimentos foram realizados em Campinas (la-titude 22o 54`S longitude 47o 3`W e altitude de 600 m),
Mococa (latitude 21o 28`S longitude 47o 01`W e altitude
665 m) e Palmital (latitude 22o 48`S longitude 50o 14`W
e altitude 501 m). Os experimentos foram semeados em Campinas e Mococa, na primeira quinzena de novem-bro de 2008 e em Palmital durante a segunda quinzena de outubro de 2008.
Foram avaliados os caracteres agronômicos floresci-mento masculino (FM), altura da espiga (AE) e produti-vidade de grãos (PG) por parcela, transformada em kg ha-1
e corrigida para 14% de umidade e estande ideal de 50 plantas por parcela, empregando-se o método da covari-ância (Vencovsky e Barriga, 1992);
Análises de variância individuais e conjuntas foram efetuadas, considerando-se o modelo fixo e as médias comparadas pelo Teste de Tukey a 5% de probabilidade.
Para análise conjunta, considerou-se a homogeneida-de das variâncias através dos quadrados médios (QM’s) dos resíduos de acordo com Banzatto e Kronka (2006), que consideram que a razão entre o maior e o menor qua-drado médio (QM) dos resíduos sendo menor que sete é uma indicação de resíduos homogêneos.
Para análise dialélica foi utilizado o método de Gardner e Eberhart (1966), de acordo com o modelo 4:
Yij = m + (pi + pj)/2 + θ (h + hi + hj + sij) + eij,
em que Yij: valor médio observado em um parental (i =
j) ou em uma combinação híbrida (i ≠ j); m: média das populações parentais; pi: efeito do parental i; pj: efeito do parental j; θ = 0, quando i = j e θ = 1, quando i ≠ j; h:
efeito da heterose média; hi: efeito da heterose do parental i; hj: efeito da heterose do parental j; sij: efeito da heterose específica; e eij: erro experimental médio.
As análises foram efetuadas empregando o programa Genes (Cruz, 2006), adaptando-se o modelo para vários ambientes. Também estimou-se a heterose em relação à média dos pais para produtividade de grãos.
3. RESULTADOS E DISCUSSÃO
Constatada a homogeneidade dos QM’s dos resíduos, efe-tuou-se a análise de variância conjunta para os três locais, havendo diferenças significativas (p<0,01) entre os trata-mentos e locais para FM, AE, e PG (Tabela 2). O efeito da interação tratamentos x locais foi significativo para FM, PG (p<0,01) e AE (p<0,05), considerando-se as médias por local destas variáveis para comparação e discussão dos resultados (Tabela 3).
Destacaram-se como mais tardios os híbridos P4 x P2 e as populações P4 e P5 em Campinas, Mococa e Palmital e o híbrido P5 x P1 em Mococa (Tabela 3). Como mais precoces, destacaram-se os híbridos P6 x P2 em Campinas e Palmital, P6 x P3 e P7 x P6 em Mococa e Palmital, P8 x P7 e P6 x P5 somente em Mococa e Palmital respec-tivamente. Ressalta-se que a média do FM dos híbridos
de F2 foram muito próximas da média das testemunhas
comerciais de ciclo precoce, evidenciando que as popu-lações em estudo atendem a objetivos propostos por pro-gramas de melhoramento de obtenção de híbridos mais precoces.
Os híbridos P4 x P2, em Campinas e P5 x P2, em Mococa e Palmital, tiveram maior valor de AE (Tabela 3). O menor valor de AE foi observado para P8 x P3, em Campinas e Mococa, entretanto, não se diferenciando es-tatisticamente das testemunhas comerciais.
A produtividade média observada foi de 8.415 kg ha-1,
7.721 kg ha-1 e 6.332 kg ha-1, respectivamente em Campinas,
FV
Quadrados médios
GL FM AE PG (1)
(d.a.s.) (2) (cm) (t ha-1)
Blocos/Locais 6 13,42 406,59 3,53 Tratamentos (T) 37 35,09** 788,96** 13,07** Locais (L) 2 4194,89** 61128,88** 128,25**
T x L 74 3,35** 145,23* 0,81**
Resíduo 222 1,49 100,33 0,49
Média 66,97 107,11 7,49
CV(%) 1,82 9,35 9,33
Tabela 2. Análise de variância conjunta dos caracteres florescimento masculino (FM), altura de espiga (AE) e produtividade de grãos (PG) de 28 híbridos de populações F2 de milho, de oito parentais e mais duas testemunhas comerciais. Campinas, Mococa e Palmital, em 2008/2009
* e **: significativo a 5% e 1%, respectivamente, pelo teste F.
Mococa e Palmital (Tabela 3).O desempenho dos híbridos foi satisfatório, com destaque para os cruzamentos P5 x P3, P2 x P1 e P5 x P2, respectivamente, em Campinas, Mococa e Palmital. Ressalta-se que oito híbridos de F2 em Campinas, cinco em Mococa e dois em Palmital tiveram PG maior (0,5% a 13%) que a média das testemunhas comerciais, evidenciando potencial de produção satisfató-rio, bem como a viabilidade de utilização dos híbridos de
populações F2 de milho em cultivos comerciais. Essa viabi-lidade também foi constatada por Amorim e Souza (2005) que obtiveram híbridos a partir de populações F2 de híbri-dos simples comerciais superiores à média das testemunhas comerciais. Souza Sobrinho et al. (2002) também consta-taram que alguns híbridos duplos de F2 avaliados foram tão produtivos quanto os melhores híbridos simples recomen-dados para a Região Sul de Minas Gerais.
Híbridos/ populações F2
FM AE PG
Camp. Moc. Palm. Camp. Moc. Palm. Camp. Moc. Palm.
(d.a.s.) (d.a.s.) (d.a.s.) (cm) (cm) (cm) (kg ha-1) (kg ha-1) (kg ha-1)
P5 x P3 70 a-f 64 a-c 69 f-k 128 d-i 107 a-d 87 a-c 10.606 a 8.965 ab 7.570 a-e P5 x P2 71 a-e 63 a-c 72 c-f 157 ab 131 a 110 a 10.309 ab 8.802 ab 8.153 ab P5 x P1 71 a-e 66 ab 71 d-h 149 a-e 129 ab 95 a-c 10.217 ab 9.001 ab 7.064 a-e P2 x P1 69 c-h 63 a-c 71 e-i 153 a-c 126 ab 102 ab 10.056 a-c 9.115 ab 7.106 a-e P5 x P4 70 b-g 64 a-c 74 a-c 131 c-i 108 a-d 88 a-c 9.878 a-d 8.757 a-c 7.215 a-e P4 x P1 70 a-f 63 a-c 71 e-i 150 a-d 114 a-d 92 a-c 9.642 a-e 8.976 ab 8.044 a-c P7 x P3 67 e-i 62 bc 68 h-k 128 d-i 94 a-d 88 a-c 9.588 a-e 7.831 a-h 7.279 a-e P4 x P2 72 a-c 66 ab 74 a-d 160 a 111 a-d 100 ab 9.583 a-e 8.942 ab 7.085 a-e P6 x P3 65 ij 60 c 67 k 117 g-i 93 a-d 75 bc 9.381 a-e 6.860 b-h 6.815 a-e P3 x P1 69 c-i 63 a-c 70 e-k 137 a-i 116 a-c 92 a-c 9.348 a-e 7.487 a-h 5.892 d-g P3 x P2 67 e-i 62 bc 69 g-k 131 c-i 97 a-d 92 a-c 9.208 a-e 6.997 b-h 6.172 c-g P7 x P1 69 c-i 63 a-c 68 h-k 145 a-f 106 a-d 88 a-c 9.077 a-f 8.476 a-e 6.472 a-e P6 x P1 67 e-i 62 bc 68 h-k 130 c-i 101 a-d 85 a-c 8.990 a-f 8.592 a-d 6.736 a-e P6 x P5 68 d-i 62 bc 67 k 123 f-i 101 a-d 75 bc 8.818 a-g 7.409 a-h 6.892 a-e P4 x P3 70 b-g 62 bc 70 e-k 124 f-i 96 a-d 88 a-c 8.807 a-h 8.439 a-e 6.805 a-e P6 x P2 62 j 62 bc 67 k 124 f-i 96 a-d 93 a-c 8.794 a-h 6.955 b-h 6.296 b-g P7 x P5 68 d-i 62 bc 68 h-k 128 d-i 99 a-d 87 a-c 8.766 a-i 8.194 a-f 7.032 a-e P7 x P2 68 d-i 61 c 68 h-k 131 c-i 95 a-d 95 a-c 8.619 a-i 8.306 a-e 7.794 a-d P8 x P2 68 d-i 61 c 68 h-k 134 b-i 117 a-c 80 a-c 8.536 a-i 8.069 a-g 6.685 a-e P7 x P4 70 b-g 62 bc 68 h-k 138 a-i 114 a-d 80 a-c 8.517 a-i 8.404 a-e 7.531 a-e P6 x P4 67 e-i 61 c 69 g-k 137 a-i 84 cd 82 a-c 8.357 a-j 7.783 a-h 6.290 b-g P8 x P1 69 c-h 63 a-c 70 e-k 139 a-h 86 cd 63 c 8.263 a-j 8.216 a-f 6.359 a-f P7 x P6 67 e-i 60 c 67 k 126 d-i 92 b-d 90 a-c 8.200 b-k 7.614 a-h 7.347 a-e P8 x P7 67 f-i 60 c 68 h-k 123 f-i 101 a-d 88 a-c 8.181 b-k 8.140 a-g 6.494 a-e P8 x P6 65 ij 61 c 68 h-k 125 d-i 107 a-d 82 a-c 7.989 b-k 8.186 a-f 5.820 e-g P8 x P3 66 hi 61 c 69 g-k 113 i 77 d 77 a-c 7.609 d-l 8.175 a-f 6.533 a-e P8 x P5 68 d-i 63 a-c 69 f-k 125 e-i 107 a-d 83 a-c 7.514 e-l 7.923 a-h 6.832 a-e P8 x P4 69 c-i 63 a-c 68 h-k 131 c-i 95 a-d 95 a-c 6.803 f-l 7.163 b-h 5.869 d-g Média dos híbridos 68,21 62,33 69,19 133,46 103,55 87,56 8.916 8.135 6.864 P1 71 a-d 66 ab 72 c-g 122 f-i 102 a-d 77 a-c 5.311 l 6.095 d-h 3.555 h P2 71 a-e 63 a-c 73 b-e 141 a-g 96 a-d 93 a-c 6.052 j-l 5.694 f-h 3.764 h P3 69 c-i 63 a-c 69 g-k 125 e-i 101 a-d 80 a-c 7.797 c-k 6.797 b-h 6.057 d-g
P4 74 a 66 ab 76 ab 141 a-g 100 a-d 100 ab 6.467 h-l 6.206 c-h 4.499 f-h
P5 73 ab 67 a 76 a 137 a-i 107 a-d 95 a-c 5.867 kl 6.040 d-h 3.486 h
P6 66 g-i 62 bc 68 ik 115 hi 78 cd 85 a-c 6.437 i-l 5.448 h 3.690 h
P7 69 c-h 63 a-c 70 e-j 128 d-i 92 b-d 83 a-c 6.579 g-l 5.987 e-h 4.370 gh P8 70 b-g 63 a-c 69 f-k 123 f-i 84 cd 77 a-c 6.764 f-l 5.578 gh 3.387 h Média dos parentais 70,50 64,04 71,50 129,00 94,96 86,25 6.409 5.981 4.101 DKB 350 67 f-i 63 a-c 67 k 129 c-i 98 a-d 83 a-c 9.636 a-e 8.282 a-e 8.260 a IAC 8333 68 d-i 61 c 68 i-k 138 a-i 104 a-d 88 a-c 9.198 a-e 9.515 a 7.371 a-e
Média 68,64 62,68 69,59 132,52 101,61 87,19 8.415 7.721 6.332
CV (%) 1,76 2,26 1,43 5,71 11,50 11,87 8,50 10,10 9,28 Tabela 3. Médias de florescimento masculino (FM), altura de espiga (AE) e produtividade de grãos (PG) de 28 híbridos de populações F2 de milho, dos oito parentais e duas testemunhas comerciais, em Campinas, Mococa e Palmital, em 2008/2009
Médias seguidas de mesma letra não diferem pelo teste de Tukey (p<0,05).
Na análise dialélica, os QM’s dos efeitos de parental e de heterose foram significativos para todos os caracte-res, indicando que há variabilidade entre os parentais e a manifestação de heterose em seus híbridos para FM, AE e PG (Tabela 4). O desdobramento dos QM’s dos efeitos de heterose em heterose média h, de parentais (hi) e especí-fica (sij) revelou significância dos efeitos de heterose média
he específica (sij) para todos os caracteres e da heterose de parental (hi) somente para AE e PG.
A significância para h indica que há suficiente
diversi-dade genética entre as populações, resultando em situação favorável à aplicação do melhoramento genético visando à obtenção de híbridos (Kvitschal et al., 2004). A signifi-cância dos efeitos de hi mostra que existe variação na con-tribuição de cada genitor para o desempenho dos híbridos, indicando dispersão nas frequências alélicas das populações (Oliveira et al., 2004), fato evidenciado para AE e PG.
A significância dos efeitos de sij indica que há presença de efeitos de dominância e epistasia na manifestação da produtivididade de grãos, altura de espiga e florescimento masculino. De acordo com Vencovsky e Barriga (1992) esta significância evidencia existência de complementa-ções específicas entre pares de genitores, em locos com algum efeito de dominância alélica, contribuindo para o melhor desempenho de certas combinações híbridas, onde um supre a deficiência genômica do outro.
A significância dos QM’s de PG para pi , h, hi e sij (Tabela 4), corrobora com os resultados de Silva e Miranda Filho (2003) e Scapim et al. (2002), que utilizaram diferen-tes populações de milho e de milho pipoca. Quanto a FM e
AE, Garbuglio e Araújo (2006), avaliando híbridos oriun-dos do cruzamento entre sete variedades de milho, não ob-servaram significância dos QM’s da heterose para FM e AE; assim como Gorgulho e Miranda Filho (2001) também não verificaram significância de heterose para AE.
Gardner et al. (1953) e Comstock e Robinson (1948) relataram que nos caracteres altura de planta e altu-ra de espiga existe predominância de efeitos aditivos e do-minância apenas parcial. Dessa forma, a significância para heterose verificada neste experimento diverge do constata-do por outros autores e indica que, entre os parentais utili-zados existe suficiente divergência das frequências gênicas, pelo menos em parte dos locos com dominância.
Em relação à decomposição da soma de quadrados da heterose na análise de variância, pode-se observar que para PG, os efeitos da h, hi sij corresponderam, respec-tivamente, a 82%, 10% e 8% do total dos seus efeitos. Nota-se contribuição maior da heterose média em relação aos demais efeitos, evidenciando existência de expressiva superioridade dos híbridos de F2 em relação à média das populações parentais. A magnitude destes efeitos corro-bora com os de Garbuglio e Araújo (2006), que avalia-ram híbridos intervarietais de milho e detectaavalia-ram efeitos semelhantes para PG, entretanto, diverge dos resultados observados por Kvitschal et al. (2004) que verificaram contribuição maior da heterose varietal, em relação à he-terose média, em trabalho que avaliou a capacidade geral de combinação de seis populações de milho e os efeitos heteróticos manifestados em seus híbridos.
Para FM, a partição da soma de quadrados da heterose também revelou maior contribuição dos efeitos da heterose média, (63%) enquanto para AE, houve superioridade dos efeitos de ŝij (60%). Estes resultados divergem dos detectados
por Gorgulho e Miranda Filho (2001) e Garbuglio e Araújo (2006) para AE, os quais verificaram significância apenas para efeitos de variedades, concluindo que no con-trole deste caráter há predominância de efeitos aditivos e de dominância apenas parcial. Ressalta-se que estes últimos au-tores utilizaram variedades, e não híbridos de populações, o que pode explicar as diferenças dos resultados.
Quanto às interações entre os efeitos de heterose com locais, observou-se que para FM, somente o efeito da heterose específica propiciou interação significativa; para AE, não foram significativos; para PG, verificou-se signifi-cância das interações entre a heterose média e de parental com os locais, enquanto os efeitos de heterose específica mantiveram-se constantes nos diferentes locais (Tabela 4). As estimativas dos efeitos de parental (pˆi), heteroses
mé-dia (hˆ) e de parental (ĥi) e heterose específica (ŝij) estão
apresentadas, respectivamente, nas tabelas 5, 6 e 7. A população P3 proporcionou as maiores estimativas de pˆi
para PG e estimativas favoráveis no sentido de redução do porte (AE) e do intervalo entre a semeadura e o floresci-mento masculino (Tabela 5). Esta população reuniu efei-tos de pˆi favoráveis nos três locais avaliados, quanto a AE,
Quadrados Médios
FV GL FM AE PG
(d.a.s.) (cm) (t ha-1)
Tratamentos (T) 35 36,01** 828,63** 13,01** Parental (P) 7 123,93** 2429,75** 5,46** Heterose (H) 28 14,03** 428,35** 14,89** H. média 1 247,38** 1283,62** 342,94**
H. parental 7 2,96 491,35** 5,68**
H. específica 20 6,24** 363,54** 1,72** Local (L) 2 4023,97** 57610,02** 124,59**
T x L 70 3,34** 153,15* 0,79**
P x L 14 5,06** 227,45** 1,21**
H x L 56 2,92** 134,58 0,68*
H. média x L 2 2,16 248,75 1,74*
H. parental x L 14 1,74 105,42 1,25** H. específica x L 40 3,36** 139,07 0,43 Resíduo Combinado 210 1,52 100,65 0,50
Média 67,05 107,13 7,42
CV (%) 1,84 9,37 9,56
Tabela 4. Análise dialélica conjunta, de acordo com o método de Gardner e Eberhart (1966), para florescimento masculino (FM), altura de espiga (AE) e produtividade de grãos (PG) de 28 híbridos
de gerações F2 de milho, dos experimentos de Campinas, Mococa e
Palmital, em 2008/2009
FM e PG, podendo ser indicada para seleção recorrente intrapopulacional e obtenção de linhagens em programas de melhoramento genético de milho.
As estimativas de heterose média (hˆ) (Tabela 6) indi-cam que para FM, a manifestação da heterose foi predomi-nante no contexto de reduzir o intervalo entre a semeadura e o FM, enquanto para AE e PG, foi predominante para aumentar seus valores. De acordo com Cruz et al. (2004), cruzamentos envolvendo parentais com maior efeito de ĥi
resultarão em combinações híbridas mais heteróticas. Desse modo, se o objetivo do programa de melhoramento for ob-ter híbridos com maior produtividade de grãos, associado ao menor porte e à maior precocidade, entre as populações avaliadas, a P5 é a que reúne efeitos de ĥi mais favoráveis.
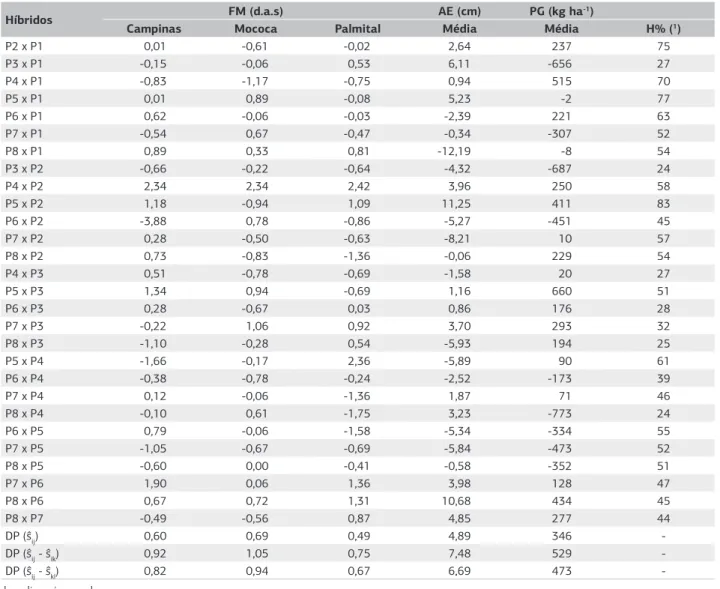
O híbrido P5 x P3 proporcionou o maior efeito de
ŝij (660 kg ha-1) para PG (Tabela 7). Entretanto, para AE,
o menor valor foi para o híbrido P8 x P1, enquanto para
FM, destacaram-se os híbridos P6 x P2 em Campinas, P4 x P1 em Mococa e P8 x P4 em Palmital. Neste grupo de híbridos evidencia-se a dificuldade em se encontrar, em um mesmo híbrido, complementações favoráveis a estas características, em locos com efeitos de dominância.
De acordo com o modelo de Gardner e Eberhart (1966), os melhores híbridos serão aqueles que reunirem um conjunto maior de efeitos favoráveis para a caracte-rística desejada. O híbrido P5 x P3 destacou-se quanto à produtividade de grãos por reunir os efeitos de pˆi favorá-veis do parental P3, os efeitos de ĥi favoráveis do parental
P5 e efeito de ŝij relativamente alto.
Com relação à porcentagem de heterose (H%), desta-caram-se os híbridos P5 x P2, P5 x P1 e P2 x P1, na média dos três locais, com valores relativamente altos (Tabela 7). A porcentagem de heterose obtida, considerando a mé-dia dos três locais, foi de 47%. Este resultado pode ser
Populações F2
ĥi
FM (d.a.s) AE (cm) PG (kg ha
-1)
Campinas Mococa Palmital
P1 0,52 8,68 1.079 429 211
P2 0,29 5,67 627 184 376
P3 0,48 -5,74 -338 -773 -1.141
P4 -0,17 -2,63 -167 141 -67
P5 -0,41 -0,59 887 321 760
P6 -0,26 -2,19 -328 -325 -103
P7 -0,30 -0,69 -329 0,3 183
P8 -0,15 -2,50 -1.431 23 -219
DP (ĥi) 0,43 3,50 253 275 210
DP (ĥi - ĥj) 0,65 5,29 382 416 317
Heterose média (hˆ) -2,10 4,79 2.507 2.154 2.763
Heterose média (%h) -3,20 4,60 39 36 67
DP (hˆ) 0,28 2,32 168 183 139
Tabela 6. Estimativas dos efeitos de heterose média (hˆ) e de parental (ĥi) para os caracteres florescimento masculino (FM) e altura de
espiga (AE), considerando a média de três locais e produtividade de grãos (PG), em Campinas, Mococa e Palmital, em 2008/2009
d.a.s: dias após semeadura.
P1: F2AG8060; P2: F2Fort; P3: F2DAS2B710; P4: F230F87; P5: F2A2555; P6: F2AS1548; P7: F230K75; P8: F2DKB330.
Populações F2 FM (d.a.s.) AE (cm) PG (kg ha
-1)
Campinas Mococa Palmital Campinas Mococa Palmital Campinas Mococa Palmital
P1 0,83 1,63 0,17 -7,33 6,71 -9,58 -1.098 114 -546
P2 0,17 -1,04 1,17 12,33 1,04 7,08 -357 -287 -337
P3 -1,50 -1,04 -2,83 -4,00 5,71 -6,25 1.388 816 1.956
P4 3,50 1,63 4,17 12,00 5,37 13,75 58 225 398
P5 2,83 2,96 4,50 7,67 12,04 8,75 -542 59 -615
P6 -4,17 -2,04 -3,83 -13,67 -16,63 -1,25 28 -533 -411
P7 -1,17 -1,04 -1,17 -1,33 -3,29 -2,92 170 6 269
P8 -0,50 -1,04 -2,17 -5,67 -10,96 -9,58 355 -403 -714
DP (pˆi) 0,67 0,76 0,55 4,06 6,25 5,70 392 426 325
DP pˆi pˆi 1,01 1,15 0,83 6,14 9,45 8,62 592 644 492
Tabela 5. Estimativas dos efeitos de parental pˆi para os caracteres florescimento masculino (FM), altura de espiga (AE) e produtividade de
grãos (PG) de oito populações F2 de milho oriundas de híbridos comerciais, em Campinas, Mococa e Palmital, em 2008/2009
d.a.s: dias após semeadura.
Híbridos FM (d.a.s) AE (cm) PG (kg ha -1)
Campinas Mococa Palmital Média Média H% (1)
P2 x P1 0,01 -0,61 -0,02 2,64 237 75
P3 x P1 -0,15 -0,06 0,53 6,11 -656 27
P4 x P1 -0,83 -1,17 -0,75 0,94 515 70
P5 x P1 0,01 0,89 -0,08 5,23 -2 77
P6 x P1 0,62 -0,06 -0,03 -2,39 221 63
P7 x P1 -0,54 0,67 -0,47 -0,34 -307 52
P8 x P1 0,89 0,33 0,81 -12,19 -8 54
P3 x P2 -0,66 -0,22 -0,64 -4,32 -687 24
P4 x P2 2,34 2,34 2,42 3,96 250 58
P5 x P2 1,18 -0,94 1,09 11,25 411 83
P6 x P2 -3,88 0,78 -0,86 -5,27 -451 45
P7 x P2 0,28 -0,50 -0,63 -8,21 10 57
P8 x P2 0,73 -0,83 -1,36 -0,06 229 54
P4 x P3 0,51 -0,78 -0,69 -1,58 20 27
P5 x P3 1,34 0,94 -0,69 1,16 660 51
P6 x P3 0,28 -0,67 0,03 0,86 176 28
P7 x P3 -0,22 1,06 0,92 3,70 293 32
P8 x P3 -1,10 -0,28 0,54 -5,93 194 25
P5 x P4 -1,66 -0,17 2,36 -5,89 90 61
P6 x P4 -0,38 -0,78 -0,24 -2,52 -173 39
P7 x P4 0,12 -0,06 -1,36 1,87 71 46
P8 x P4 -0,10 0,61 -1,75 3,23 -773 24
P6 x P5 0,79 -0,06 -1,58 -5,34 -334 55
P7 x P5 -1,05 -0,67 -0,69 -5,84 -473 52
P8 x P5 -0,60 0,00 -0,41 -0,58 -352 51
P7 x P6 1,90 0,06 1,36 3,98 128 47
P8 x P6 0,67 0,72 1,31 10,68 434 45
P8 x P7 -0,49 -0,56 0,87 4,85 277 44
DP (ŝij) 0,60 0,69 0,49 4,89 346
-DP (ŝij - ŝik) 0,92 1,05 0,75 7,48 529
-DP (ŝij - ŝkl) 0,82 0,94 0,67 6,69 473
-Tabela 7. Estimativas dos efeitos de heterose específica (ŝij) de 28 híbridos de gerações F2 de milho para florescimento masculino (FM),
em Campinas, Mococa e Palmital, e para altura de espiga (AE) e produtividade de grãos (PG), considerando as médias dos três locais, em 2008/2009
d.a.s: dias após semeadura.
(1) Porcentagem de heterose em relação à média dos parentais, considerando a média de Campinas, Mococa e Palmital. P1: F2AG8060; P2: F2Fort; P3: F2DAS2B710; P4: F230F87; P5: F2A2555; P6: F2AS1548; P7: F230K75; P8: F2DKB330.
considerado de boa magnitude, quando comparado ao valor de 19,5% apresentado por Hallauer e Miranda Filho (1981), referente a 1.394 cruzamentos interva-rietais de milho e evidencia que há divergência entre as populações F2 de milho utilizadas neste experimento. É provável que os altos valores de H% sejam porque estas populações, oriundas de gerações F2 de híbridossimples, possuem base genética mais estreita, com certo grau de endogamia, o que acarreta em alta expressão da heterose quando combinadas (Silva e Miranda Filho, 2003).
4. CONCLUSÃO
As populações P1 (F2AG8060), P2 (F2Fort) e P5 (F2A2555) são promissoras em combinações híbridas visando à maior produtividade, com elevados efeitos de heterose de paren-tal para produtividade de grãos, enquanto a população
P3 (F2DAS2B710) é a que proporciona combinações
hí-bridas de menor porte e altura de espiga. A população P3 (F2DAS2B710) possui bom potencial per se e pode ser indi-cada para seleção recorrente intrapopulacional e obtenção de linhagens em programas de melhoramento de milho.
Há manifestação de heterose entre os híbridos de po-pulações F2 para produtividade de grãos e confirma-se o potencial produtivo destes híbridos como alternativa para a produção comercial de milho, visando aos produtores de média a baixa tecnologia, com destaque para os
cru-zamentos P5 x P3 (F2A2555 x F2DAS2B710), P5 x P2
(F2A2555 x F2Fort), P5 x P1 (F2A2555 x F2AG8060).
AGRADECIMENTOS
REFERÊNCIAS
AMORIM, E.P.; SOUZA, J.C. Híbridos de milho inter e intrapopulacionais obtidos a partir de populações S0 de híbridos simples comerciais. Bragantia, v.64, p.561-567, 2005.
BANZATTO, D.A.; KRONKA, S.N. Experimentação agrícola. 4.ed. Jaboticabal: Funep, 2006. 237p.
CARVALHO, A.D.F.; SOUZA, J.C.; RIBEIRO, P.H. Desempenho de híbridos de linhagens parcialmente endogâmicas de milho em regiões dos Estados de Roraima e Minas Gerais. Ciência e Agrotecnologia, v.27, p.985-990, 2003.
CONAB – COMPANHIA NACIONAL DE ABASTECIMENTO, 2009. Disponível em: http://www.conab.gov.br/conabweb/download/ safra/12_levantamento_set2009.pdf. Acesso em: 21/11/2009.
COMSTOCK, R.E.; ROBINSON, H.F. The components of genetic variance in populations of biparental progenies and their use in estimating the average degree of dominance. Biometrics, v.4, p.254-266, 1948.
CRUZ, C.D. Programa GENES: biometria. Viçosa: UFV, 2006, 382p.
CRUZ, C.D.; REGAZZI, A.J.; CARNEIRO, P.C.S. Modelos Biométricos Aplicados ao Melhoramento Genético. v.1. 3. ed. Viçosa: UFV, 2004. 480p.
CRUZ, J.C.; PEREIRA FILHO, I.A. Cultivo do Milho. http://
sistemasdeproducao. cnptia.embrapa.br/FontesHTML/Milho/
CultivodoMilho_5ed/cultivares.htm. Acesso em 20/4/ 2010.
GARBUGLIO, D.D.; ARAÚJO, P.M. Avaliação de Híbridos Intervarietais de Milho por Meio de Cruzamento Dialélico Parcial, Considerando Quatro Ambientes. Semina: Ciências Agrárias, v.27, p.379-392, 2006.
GARDNER, C.O.; EBERHART, S.A. Analysis and interpretation of the variety cross diall and related populations. Biometrics, v.22, p.439-452, 1966.
GARDNER, C.O.; HARVEY, P.H.; COMSTOCK, R.E. Dominance of genes controlling quantitative characters in maize. Biometrics, v.45, p.186-191, 1953
GORGULHO, E.P.; MIRANDA FILHO, J.B. Estudo da capacidade combinatória de variedades de milho no esquema de cruzamento dialélico parcial. Bragantia, v.60, p.1-8, 2001.
GRIFFING, J.B. Concept of general and specific combining ability in relation to diallel systems. Australian Journal of Biological Science, v.9, p.463-493, 1956
KVITSCHAL, M.V.; SCAPIM, C.A.; TONET, A.; PINTO, R.J.B.; RETUCI, V.S.; AMARAL JÚNIOR, A.T.; BRACCINI, A.L.. Análise dialélica de populações de milho na região noroeste do Paraná, na “safrinha”. Revista Ceres, v.51, p.19-32, 2004.
HALLAUER, A.R.; MIRANDA FILHO, J.B. Quantitative genetics in maize breeding. Ames: Iowa State University Press, 468p. 1981.
MATTOSO, M.J.; MELO FILHO, G.A. Cultivo do Milho. EMBRAPA – Empresa Brasileira de Pesquisa Agropecuária, 2006. Disponível em: http://sistemasdeproducao.cnptia. embrapa.br/ FontesHTML/Milho/CultivodoMilho_2ed/coeficientestecnicos. htm. Acesso em: 28/9/2010.
MIRANDA FILHO, J.B.; VIÉGAS, G.P. Milho híbrido. In: PATERNIANI, E.; VIÉGAS, G.P. (Ed.). Melhoramento e produção do milho. 2.ed. Campinas: Fundação Cargill, v.1, p.277-326, 1987.
OLIVEIRA, J.P.; CHAVES, L.J.; DUARTE, J.B.; BRASIL, E.M.; FERREIRA JUNIOR., L. F.; RIBEIRO, K.O. Teor de proteína no grão em populações de milho de alta qualidade protéica e seus cruzamentos. Pesquisa Agropecuária Tropical, v.34, p.45-51, 2004.
SCAPIM, C.A.; PACHECO, C.A.P.; TONET, A.; BRACCINI, A.L.; PINTO, R.J.B. Análise dialélica e heterose de populações de Milho-Pipoca. Bragantia, v.61, p.219-230, 2002.
SILVA, R.M.; MIRANDA FILHO, J.B. Heterosis expression in
crosses between maize populations: ear yield. Scientia Agricola,
v.60, p.519-524, 2003.
SOUZA SOBRINHO, F.; RAMALHO, M.A.P.; SOUZA J.C. Alternatives for obtaining double cross maize hybrids. Revista Brasileira de Milho e Sorgo, v.1, p.70-76, 2002.
USDA – UNITED STATES DEPARTMENT OF
AGRICULTURE. 2010. Disponível em: http://www.fas.usda.gov/ psdonline/psdreport.aspx?hidReportRetrievalName =BVS&hidRe portRetrievalID=884&hidReportRetrievalTemplateID=1. Acesso em: 8/3/2010.